TW202108626A - 與cd40及fap結合之雙專一性抗體 - Google Patents
與cd40及fap結合之雙專一性抗體 Download PDFInfo
- Publication number
- TW202108626A TW202108626A TW109116232A TW109116232A TW202108626A TW 202108626 A TW202108626 A TW 202108626A TW 109116232 A TW109116232 A TW 109116232A TW 109116232 A TW109116232 A TW 109116232A TW 202108626 A TW202108626 A TW 202108626A
- Authority
- TW
- Taiwan
- Prior art keywords
- antibody
- fap
- bispecific antibody
- amino acid
- acid sequence
- Prior art date
Links
Images
















Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/40—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against enzymes
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P35/00—Antineoplastic agents
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/28—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants
- C07K16/2878—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against receptors, cell surface antigens or cell surface determinants against the NGF-receptor/TNF-receptor superfamily, e.g. CD27, CD30, CD40, CD95
-
- G01N33/575—
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/31—Immunoglobulins specific features characterized by aspects of specificity or valency multispecific
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/30—Immunoglobulins specific features characterized by aspects of specificity or valency
- C07K2317/35—Valency
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/40—Immunoglobulins specific features characterized by post-translational modification
- C07K2317/41—Glycosylation, sialylation, or fucosylation
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/52—Constant or Fc region; Isotype
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/52—Constant or Fc region; Isotype
- C07K2317/522—CH1 domain
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/52—Constant or Fc region; Isotype
- C07K2317/526—CH3 domain
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/55—Fab or Fab'
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/60—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments
- C07K2317/64—Immunoglobulins specific features characterized by non-natural combinations of immunoglobulin fragments comprising a combination of variable region and constant region components
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/75—Agonist effect on antigen
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/76—Antagonist effect on antigen, e.g. neutralization or inhibition of binding
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/90—Immunoglobulins specific features characterized by (pharmaco)kinetic aspects or by stability of the immunoglobulin
- C07K2317/92—Affinity (KD), association rate (Ka), dissociation rate (Kd) or EC50 value
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/435—Assays involving biological materials from specific organisms or of a specific nature from animals; from humans
- G01N2333/705—Assays involving receptors, cell surface antigens or cell surface determinants
- G01N2333/70578—NGF-receptor/TNF-receptor superfamily, e.g. CD27, CD30 CD40 or CD95
-
- G—PHYSICS
- G01—MEASURING; TESTING
- G01N—INVESTIGATING OR ANALYSING MATERIALS BY DETERMINING THEIR CHEMICAL OR PHYSICAL PROPERTIES
- G01N2333/00—Assays involving biological materials from specific organisms or of a specific nature
- G01N2333/90—Enzymes; Proenzymes
- G01N2333/914—Hydrolases (3)
- G01N2333/948—Hydrolases (3) acting on peptide bonds (3.4)
- G01N2333/95—Proteinases, i.e. endopeptidases (3.4.21-3.4.99)
- G01N2333/964—Proteinases, i.e. endopeptidases (3.4.21-3.4.99) derived from animal tissue
- G01N2333/96425—Proteinases, i.e. endopeptidases (3.4.21-3.4.99) derived from animal tissue from mammals
- G01N2333/96427—Proteinases, i.e. endopeptidases (3.4.21-3.4.99) derived from animal tissue from mammals in general
- G01N2333/9643—Proteinases, i.e. endopeptidases (3.4.21-3.4.99) derived from animal tissue from mammals in general with EC number
- G01N2333/96433—Serine endopeptidases (3.4.21)
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Immunology (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Genetics & Genomics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Veterinary Medicine (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Pharmacology & Pharmacy (AREA)
- Animal Behavior & Ethology (AREA)
- Public Health (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Peptides Or Proteins (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Engineering & Computer Science (AREA)
- Hematology (AREA)
- Urology & Nephrology (AREA)
- Biomedical Technology (AREA)
- Microbiology (AREA)
- Cell Biology (AREA)
- Biotechnology (AREA)
- Oncology (AREA)
- Hospice & Palliative Care (AREA)
- Food Science & Technology (AREA)
- Physics & Mathematics (AREA)
- Analytical Chemistry (AREA)
- General Physics & Mathematics (AREA)
- Pathology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
本發明之目的在於提供一種與CD40及FAP結合且顯示FAP依賴性之強CD40促效劑活性的雙專一性抗體或該雙專一性抗體片段等。本發明係關於一種雙專一性抗體或該雙專一性抗體片段等,該雙專一性抗體係含有包含第一抗原結合域之IgG部分及第二抗原結合域且與人類CD40及人類FAP結合者,該IgG部分之重鏈之C末端直接或經由連接子與該第二抗原結合域結合。
Description
本發明係關於一種包含與CD40結合之抗原結合域及與纖維母細胞活化蛋白α(Fibroblast activation protein alpha,FAP)結合之抗原結合域之雙專一性抗體或該雙專一性抗體片段、編碼該雙專一性抗體或該雙專一性抗體片段之DNA、包含該DNA之載體、生產該雙專一性抗體之融合瘤及轉形株、該雙專一性抗體或該雙專一性抗體片段之製造方法、包含該雙專一性抗體或該雙專一性抗體片段之治療劑及診斷劑、使用該雙專一性抗體或該雙專一性抗體片段之治療方法及診斷方法、及包含該雙專一性抗體之檢測或測定用試劑。
CD40係鑑定為於人類B細胞表面表現之抗原之I型膜型糖蛋白質,且已知亦於B淋巴細胞、樹狀細胞、單核球、上皮細胞及纖維母細胞等多種細胞類型、或腫瘤性人類B細胞等某種腫瘤細胞表現。於CD40缺失小鼠中,確認到胸腺依賴之免疫球蛋白之類型轉變或胚芽中心之形成受到損害,證實了CD40於細胞性及液性免疫反應中之重要作用。由於CD40訊息會參與免疫球蛋白之類型轉變或細胞毒殺性T細胞(Cytotoxic T Lymphocyte;CTL)之誘導,故亦期待作為腫瘤免疫之活化或癌症疫苗之佐劑應用於醫藥品(非專利文獻1)。
FAP係85 kDa之跨膜型糖蛋白質,形成二聚物。FAP屬於絲胺酸蛋白酶超家族,具有肽酶活性。FAP於胰腺癌、乳癌、胃癌等各種癌症種類中,於浸潤於腫瘤內之纖維母細胞CAF(Cancer-associated fibroblast,癌相關纖維母細胞)、或黑色素瘤等一部分癌細胞中高表現。於健康者之正常組織中,僅於存在於淋巴組織中之纖維母細胞或骨髄中之間葉系幹細胞等至少一部分部位表現。著眼於該侷限之表現特性,而期待作為癌症分子靶向醫藥品之靶分子或診斷標記物、癌症疫苗靶(非專利文獻2)。
作為以CD40為靶之既有抗體,可列舉:Chi-Lob 7/4、HCD-122、APX005M、SEA-CD40及CP870,893(21.4.1)等(非專利文獻3)。其中,CP870,893具有強力之CD40訊息誘導能力,並對實體腫瘤實施了以全身性之免疫活化作為藥效機制之臨床試驗。然而,並未顯示出有效性,且報告有細胞激素綜合症、血栓標記物之上升、肝臟參數之上升等源自全身性免疫活化之毒性之表現(非專利文獻4)。
作為識別CD40之雙專一性蛋白質,已知有識別CD28及CD40之多價抗體(專利文獻1)。已知有具有經異型二聚化之重鏈之IgG型抗人類GPC3/抗小鼠CD40雙專一性抗體(專利文獻2)。又,已知有與結合素4(nectin-4)、PSMA、EGFR、HER2及MSLN等癌抗原及CD40結合、使CD40活化之雙專一性蛋白質(專利文獻3、4)。該等雙專一性抗體均具有源自促效性抗CD40抗體之CD40結合部位。
進而,已知有藉由對CD40及癌細胞表面抗原具有特異性之雙功能抗體(Diabody)等雙專一性分子而於癌細胞附近使CD40表現細胞活化的方法(專利文獻5)。
又,報告有具有源自促效性抗CD40抗體SGN-40之CD40結合部位及FAP結合部位,且FAP依賴性地使CD40活化之雙專一性蛋白質(專利文獻6)。已知有具有源自錨蛋白重複序列蛋白質之CD40結合性模組與FAP結合性模組之雙專一性蛋白質,且已知其FAP特異性地誘導CD40訊息,使樹狀細胞、B細胞及巨噬細胞活化(非專利文獻5、6)。
先前技術文獻
專利文獻
專利文獻1:國際公開第2009/131239號
專利文獻2:國際公開第2015/156268號
專利文獻3:國際公開第2017/205738號
專利文獻4:國際公開第2018/140831號
專利文獻5:國際公開第99/61057號
專利文獻6:國際公開第2018/185045號
非專利文獻
非專利文獻1:Diehl L et al., Nat Med. 1999 Jul; 5 (7):774-9.
非專利文獻2:Guan-Min Jiang et al., Oncotarget. 2016 May 31; 7 (22):33472-82
非專利文獻3:Beatty GL et al., Expert Rev Anticancer Ther. 2017 Feb; 17 (2):175-186.
非專利文獻4:Vonderheide RH et al., Oncoimmunology. 2013 Jan 1; 2 (1):e23033.
非專利文獻5:Fiedler U. et al., AACR Annual Meeting 2018, poster 4552. (April 14-18, 2018)
非專利文獻6:Rigamonti N. et al., SITC 33rd Annual Meeting 2018, poster 637. (November 9-11, 2018)
[發明所欲解決之問題]
本發明之目的在於提供一種與CD40及FAP結合且僅於FAP存在下顯示較強之CD40促效劑活性之雙專一性抗體或該雙專一性抗體片段、編碼該雙專一性抗體或該雙專一性抗體片段之DNA、包含該DNA之載體、生產該雙專一性抗體或該雙專一性抗體片段之融合瘤及轉形株、該雙專一性抗體或該雙專一性抗體片段之製造方法、包含該雙專一性抗體或該雙專一性抗體片段之治療劑及診斷劑、使用該雙專一性抗體或該雙專一性抗體片段之治療方法及診斷方法、以及包含該雙專一性抗體或該雙專一性抗體片段之檢測或測定用試劑等。
[解決問題之技術手段]
作為用以解決上述課題之方法,提供一種雙專一性抗體及該雙專一性抗體片段等,該雙專一性抗體係含有包含第一抗原結合域之IgG部分及第二抗原結合域且與人類CD40及人類FAP結合者,且該IgG部分之重鏈之C末端直接或經由連接子與該第二抗原結合域結合。
即,本發明係關於以下。
1.一種雙專一性抗體,其係含有包含第一抗原結合域之IgG部分及第二抗原結合域且與人類CD40及人類纖維母細胞活化蛋白α(FAP)結合者,且該IgG部分之重鏈之C末端直接或經由連接子與該第二抗原結合域結合。
2.如上述1記載之雙專一性抗體,其中上述第一抗原結合域及上述第二抗原結合域之任一者源自非促效性抗CD40抗體。
3.如上述1或2記載之雙專一性抗體,其以二價分別與人類CD40及人類FAP結合。
4.如上述1至3中任一項記載之雙專一性抗體,其中上述IgG部分之重鏈之C末端與上述第二抗原結合域直接結合。
5.如上述1至4中任一項記載之雙專一性抗體,其具有CD40促效劑活性。
6.如上述1至5中任一項記載之雙專一性抗體,其於不存在FAP表現細胞下不顯示CD40促效劑活性,僅於FAP表現細胞之存在下顯示CD40促效劑活性。
7.如上述1至6中任一項記載之雙專一性抗體,其中上述第一抗原結合域及上述第二抗原結合域之任一者為抗CD40 IgG抗體之Fab(抗CD40 Fab)。
8.如上述1至7中任一項記載之雙專一性抗體,其中上述第一抗原結合域及上述第二抗原結合域之任一者為抗FAP抗體之Fab(抗FAP Fab)。
9.如上述1至8中任一項記載之雙專一性抗體,其中上述第一抗原結合域為抗CD40 Fab,上述第二抗原結合域為抗FAP Fab。
10.如上述1至8中任一項記載之雙專一性抗體,其中上述第一抗原結合域為抗FAP Fab,上述第二抗原結合域為抗CD40 Fab。
11.如上述7、9或10記載之雙專一性抗體,其中上述抗CD40 Fab為包含如下重鏈可變區(VH)及輕鏈可變區(VL)之Fab,該重鏈可變區(VH)含有分別包含序列編號24~26所表示之胺基酸序列之互補決定區(CDR)1~3,該輕鏈可變區(VL)含有分別包含序列編號19~21所表示之胺基酸序列之CDR1~3。
12.如上述7及9至11中任一項記載之雙專一性抗體,其中上述抗CD40 Fab為包含如下VH及VL之Fab,該VH包含序列編號23所表示之胺基酸序列,該VL包含序列編號18所表示之胺基酸序列。
13.如上述8至12中任一項記載之雙專一性抗體,其中上述抗FAP Fab為包含如下VH及VL之Fab,該VH包含選自以下(i)~(v)之任一個重鏈CDR1~3,該VL含有分別包含序列編號19~21所表示之胺基酸序列之CDR1~3。
(i)分別包含序列編號33~35所表示之胺基酸序列之重鏈CDR1~3、
(ii)分別包含序列編號38~40所表示之胺基酸序列之重鏈CDR1~3、
(iii)分別包含序列編號43~45所表示之胺基酸序列之重鏈CDR1~3、
(iv)分別包含序列編號48~50所表示之胺基酸序列之重鏈CDR1~3及
(v)分別包含序列編號53~55所表示之胺基酸序列之重鏈CDR1~3。
14.如上述8至13中任一項記載之雙專一性抗體,其中上述抗FAP Fab為包含如下VH及VL之Fab,該VH包含選自序列編號32、37、42、47、52、75、77、79、81及83之任一者所表示之胺基酸序列,該VL包含序列編號18所表示之胺基酸序列。
15.如上述1至14中任一項記載之雙專一性抗體,其中上述IgG部分含有包含序列編號61所表示之胺基酸序列之重鏈恆定區。
16.如上述1至9及11至15中任一項記載之雙專一性抗體,其包含2條重鏈及4條輕鏈,該重鏈包含選自序列編號65、67、69、71、73、85、87、89、91及93之任一者所表示之胺基酸序列,該輕鏈含有包含序列編號18所表示之胺基酸序列之VL。
17.如上述1至8及10至15中任一項記載之雙專一性抗體,其包含2條重鏈及4條輕鏈,該重鏈包含選自序列編號95、97、99、101、103、105、107、109及111之任一者所表示之胺基酸序列,該輕鏈含有包含序列編號18所表示之胺基酸序列之VL。
18.一種雙專一性抗體片段,其係如上述1至17中任一項記載之雙專一性抗體之雙專一性抗體片段。
19.一種DNA,其編碼如上述1至17中任一項記載之雙專一性抗體或如上述18記載之雙專一性抗體片段。
20.一種重組體載體,其含有如上述19記載之DNA。
21.一種轉形株,其係將上述20所記載之重組體載體導入至宿主細胞而獲得。
22.一種如上述1至17中任一項記載之雙專一性抗體或如上述18記載之雙專一性抗體片段之製造方法,其特徵在於:將如上述21記載之轉形株於培養基中進行培養,使如上述1至17中任一項記載之雙專一性抗體或如上述18記載之雙專一性抗體片段於培養物中產生並蓄積,自該培養物採集雙專一性抗體或如上述18記載之雙專一性抗體片段。
23.一種與CD40及FAP之至少一者相關之疾病之治療藥及/或診斷藥,其含有如上述1至17中任一項記載之雙專一性抗體或如上述18記載之雙專一性抗體片段作為有效成分。
24.如上述23記載之治療藥及/或診斷藥,其中與CD40及FAP之至少一者相關之疾病為癌症。
25.一種與CD40及FAP之至少一者相關之疾病之治療方法及/或診斷方法,其使用如上述1至17中任一項記載之雙專一性抗體或如上述18記載之雙專一性抗體片段。
26.如上述25記載之方法,其中與CD40及FAP之至少一者相關之疾病為癌症。
27.一種如上述1至17中任一項記載之雙專一性抗體或如上述18記載之雙專一性抗體片段,其用以治療及/或診斷與CD40及FAP之至少一者相關之疾病。
28.如上述27記載之雙專一性抗體或該雙專一性抗體片段,其中與CD40及FAP之至少一者相關之疾病為癌症。
29.一種如上述1至17中任一項記載之雙專一性抗體或如上述18記載之雙專一性抗體片段之用途,其係用於製造與CD40及FAP之至少一者相關之疾病之治療劑及/或診斷劑。
30.如上述28記載之用途,其中與CD40及FAP之至少一者相關之疾病為癌症。
31.一種用以檢測或測定CD40及FAP之至少一者之試劑,其包含如上述1至17中任一項記載之雙專一性抗體或如上述18記載之雙專一性抗體片段。
32.一種雙專一性抗體或該雙專一性抗體片段之衍生物,其係使放射性同位素、低分子藥劑、高分子藥劑、蛋白質或抗體醫藥以化學或基因工程方式與如上述1至17中任一項記載之雙專一性抗體或如上述18記載之雙專一性抗體片段結合而成。
[發明之效果]
藉由本發明,可提供一種與CD40及FAP結合且僅於FAP存在下顯示較強之CD40促效劑活性之雙專一性抗體或該雙專一性抗體片段、編碼該雙專一性抗體或該雙專一性抗體片段之DNA、包含該DNA之載體、產生該雙專一性抗體或該雙專一性抗體片段之融合瘤及轉形株、該雙專一性抗體或該雙專一性抗體片段之製造方法、包含該雙專一性抗體之治療劑及診斷劑、使用該雙專一性抗體或該雙專一性抗體片段之治療方法及診斷方法、以及包含該雙專一性抗體或該雙專一性抗體片段之檢測或測定用試劑。
本發明係關於一種與人類CD40及人類FAP特異性結合之雙專一性抗體(以下,記載為本發明之雙專一性抗體)。
本發明中之CD40係以與TNF receptor superfamily member 5(TNFRSF5)、Bp50、CDW40、MGC9013及p50相同之含義使用。作為CD40,例如可列舉:包含NCBI(http://www.ncbi.nlm.nih.gov/)中基因庫登錄號(GenBank Accession No.)NP_001241或序列編號6所示之胺基酸序列之人類CD40、及包含基因庫登錄號XP_005569274或序列編號8所示之胺基酸序列之猴CD40等。又,例如可列舉:包含於序列編號6、基因庫登錄號NP_001241、基因庫登錄號XP_005569274或序列編號8所示之胺基酸序列中缺失、置換或附加1個以上之胺基酸而成之胺基酸序列且具有CD40之功能的多肽。
本發明之CD40亦包括如下多肽:包含與序列編號6、基因庫登錄號NP_001241、基因庫登錄號XP_005569274或序列編號8所示之胺基酸序列具有通常70%以上、較佳為80%以上、進而較佳為90%以上之同源性之胺基酸序列的多肽;最佳為包含具有95%、96%、97%、98%及99%以上之同源性之胺基酸序列且具有CD40之功能的多肽。
關於具有於序列編號6、基因庫登錄號NP_001241、基因庫登錄號XP_005569274或序列編號8所示之胺基酸序列中缺失、置換或附加1個以上之胺基酸殘基而成之胺基酸序列的多肽,可藉由使用定點突變導入法[Molecular Cloning, A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press (1989)、Current Protocols in Molecular Biology, John Wiley & Sons (1987-1997)、Nucleic Acids Research, 10, 6487 (1982)、Proc. Natl. Acad. Sci. USA, 79, 6409 (1982)、Gene, 34, 315 (1985)、Nucleic Acids Research, 13, 4431 (1985)、Proceeding of the National Academy of Sciences in USA, 82, 488 (1985)]等,對例如編碼序列編號6、基因庫登錄號NP_001241、基因庫登錄號XP_005569274或序列編號8所示之胺基酸序列之DNA導入定點突變而獲得。缺失、置換或附加之胺基酸之數量並無特別限定,較佳為1個~數十個、例如1~20個、更佳為1個~數個、例如1~5個胺基酸。
作為編碼CD40之基因,例如可列舉:序列編號5或基因庫登錄號NM_001250所示之人類CD40之鹼基序列、及序列編號7或基因庫登錄號XM_011766922所示之猴CD40之鹼基序列等。又,本發明之編碼CD40之基因例如亦包括:包含編碼如下多肽之DNA之基因,該多肽包含於序列編號5、基因庫登錄號NM_001250、序列編號7或基因庫登錄號XM_011766922所示之鹼基序列中缺失、置換或附加1個以上之鹼基而成之鹼基序列,且具有CD40之功能;包含編碼如下多肽之DNA之基因,該多肽包含與序列編號5、基因庫登錄號NM_001250、序列編號7或基因庫登錄號XM_011766922所示之鹼基序列具有60%以上之同源性之鹼基序列、更佳為具有80%以上之同源性之鹼基序列、進而較佳為具有95%以上之同源性之鹼基序列,且具有CD40之功能;以及包含編碼如下多肽之DNA之基因,該多肽包含與包含序列編號5、基因庫登錄號NM_001250、序列編號7或基因庫登錄號XM_011766922所示之鹼基序列之DNA在嚴格條件下進行雜交之DNA,且具有CD40之功能;等。
作為在嚴格條件下進行雜交之DNA係指藉由如下方法所獲得之能夠進行雜交之DNA:例如將具有序列編號5、基因庫登錄號NM_001250、序列編號7或基因庫登錄號XM_011766922所示之鹼基序列之DNA用於探針之菌落雜交法、噬菌斑雜交法、南方墨點雜交法、或DNA微陣列法等。具體而言,可列舉:使用固定有源自經雜交之菌落或噬菌斑之DNA、或者具有該鹼基序列之PCR產物或寡DNA的過濾器或載玻片,於0.7~1.0 mol/L之氯化鈉存在下,於65℃下進行雜交法[Molecular Cloning, A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press (1989)、Current Protocols in Molecular Biology, John Wiley & Sons (1987-1997)、DNA Cloning 1: Core Techniques, A Practical Approach, Second Edition, Oxford University (1995)]後,使用0.1~2倍濃度之SSC溶液(1倍濃度之SSC溶液之組成包含150 mmol/L氯化鈉、15 mmol/L檸檬酸鈉),於65℃條件下將過濾器或載玻片洗淨,藉此能夠進行鑑定之DNA。作為能夠進行雜交可能之DNA,例如可列舉:與序列編號5、基因庫登錄號NM_001250、序列編號7或基因庫登錄號XM_011766922所示之鹼基序列具有較佳為60%以上之同源性之DNA、更佳為具有80%以上之同源性之DNA、進而較佳為具有95%以上之同源性之DNA。
關於編碼真核生物之蛋白質之基因之鹼基序列,經常可見基因之多態性。本發明之編碼CD40之基因中亦包括本發明中所使用之基因中由於此種多態性而於鹼基序列中產生小規模突變而成之基因。
關於本發明中之同源性之數值,除特別標明之情形以外,可為使用業者公知之同源性檢索程式所算出之數值,關於鹼基序列,可列舉:BLAST[J. Mol. Biol., 215, 403 (1990)]中使用預設之參數所算出之數值等,關於胺基酸序列,可列舉:於BLAST2[Nucleic Acids Research,25, 3389 (1997)、Genome Research, 7, 649 (1997)、http:// www.ncbi.nlm.nih.gov/ Education/BLASTinfo/information3.html]中使用預設之參數所算出之數值等。
作為預設之參數,G(起始空位罰分(Cost to open gap))於鹼基序列之情形時為5,於胺基酸序列之情形時為11,-E(空位延伸罰分(Cost to extend gap))於鹼基序列之情形時為2,於胺基酸序列之情形時為1,-q(核苷酸錯配罰分(Penalty for nucleotide mismatch))為-3,-r(核苷酸匹配獎分(reward for nucleotide match))為1,-e(期望值(expect value))為10,-W(序列長度(wordsize))於鹼基序列之情形時為11個殘基,於胺基酸序列之情形時為3個殘基,-y[以位數表示之非空位延伸下降值(Dropoff(X)for blast extensions in bits)]於blastn之情形時為20,於blastn以外之程式中為7,-X(以位數表示之空位比對下降值(X dropoff value for gapped alignment in bits))為15,且-Z(以位數表示之最終空位比對下降值(final X dropoff value for gapped alignment in bits))於blastn之情形時為50,於blastn以外之程式中為25(http://www.ncbi.nlm.nih.gov/blast/html/blastcgihelp.html)。
包含CD40之胺基酸序列之部分序列之多肽可藉由業者公知之方法進行製作,例如可藉由如下方式進行製作:使編碼序列編號6、基因庫登錄號NP_001241、基因庫登錄號XP_005569274、或序列編號8所示之胺基酸序列之DNA之一部分缺失,導入包含其之表現載體而獲得轉形體,將該轉形體進行培養。又,基於藉由上述方法所製作之多肽或DNA,並藉由與上述相同之方法,例如可獲得具有於序列編號6、基因庫登錄號NP_001241、基因庫登錄號XP_005569274、或序列編號8所示之胺基酸序列之部分序列中缺失、置換或附加1個以上之胺基酸而成之胺基酸序列的多肽。進而,包含CD40之胺基酸序列之部分序列之多肽、或具有於CD40之胺基酸序列之部分序列中缺失、置換或附加1個以上之胺基酸而成之胺基酸序列之多肽亦可藉由茀基甲氧基羰基(Fmoc)法、第三丁氧基羰基(tBoc)法等化學合成法進行製造。
作為本發明中之CD40之細胞外區,例如可列舉:使用公知之跨膜區預測程式SOSUI(http:// sosui.proteome.bio.tuat.ac.jp/sosuiframe0.html)、TMHMM ver.2(http:// www.cbs.dtu.dk/services/TMHMM-2.0/)或ExPASy Proteomics Server(http://Ca.expasy.org/)等對基因庫登錄號NP_001241所示之人類CD40之胺基酸序列進行預測所得之區域等。具體而言,可列舉:序列編號6或基因庫登錄號NP_001241之第21號~第194號所示之胺基酸序列、序列編號2所示之胺基酸序列。
作為CD40之功能,可列舉:於結合有CD40配位體或促效劑時誘導CD40訊息而引起各種作用。例如若對癌細胞誘導CD40訊息,則引起該癌細胞之細胞死亡或增生阻礙等。若對B淋巴細胞誘導CD40訊息,則例如引起該B淋巴細胞之活化、CD95之表現促進、類型轉變重組及體細胞超突變(somatic hypermutation)等,而誘導抗原高親和性抗體產生等。若對樹狀細胞誘導CD40訊息,則例如引起該樹狀細胞之成熟化或IL-12產生。若對巨噬細胞誘導CD40訊息,則例如引起M2巨噬細胞之表面標記物之減少、及M1巨噬細胞之表面標記物之表現之誘導或促炎性(pro-inflammatory)細胞激素產生。
所謂CD40訊息,係指經由CD40傳遞至細胞內之訊息。
本發明中之FAP係以與FAPA、SIMP、DPPIV、FAPalpha、分離酶(Separase)相同之含義使用。
作為FAP,例如可列舉:包含基因庫登錄號NP_004451或序列編號14所示之胺基酸序列之人類FAP或者包含基因庫登錄號NP_032012或序列編號16所示之胺基酸序列之小鼠FAP等。又,例如可列舉:包含於序列編號14或16、基因庫登錄號NP_004451或基因庫登錄號NP_032012所示之胺基酸序列中缺失、置換或附加1個以上之胺基酸而成之胺基酸序列且具有FAP之功能的多肽。
本發明之FAP亦包括如下多肽:包含與序列編號14或16、基因庫登錄號NP_004451或基因庫登錄號NP_032012所示之胺基酸序列具有較佳為70%以上、更佳為80%以上、進而較佳為90%以上之同源性之胺基酸序列的多肽;最佳為包含具有95%、96%、97%、98%及99%以上之同源性之胺基酸序列且具有FAP之功能之多肽。
關於具有於序列編號14或16、基因庫登錄號NP_004451或基因庫登錄號NP_032012所示之胺基酸序列中缺失、置換或附加1個以上之胺基酸殘基而成之胺基酸序列的多肽,可藉由使用上述定點突變導入法等,對例如編碼序列編號14或16、基因庫登錄號NP_004451或基因庫登錄號NP_032012所示之胺基酸序列之DNA導入定點突變而獲得。缺失、置換或附加之胺基酸之數量並無特別限定,較佳為1個~數十個、例如1~20個、更佳為1個~數個、例如1~5個胺基酸。
作為本發明中之編碼FAP之基因,例如可列舉:包含基因庫登錄號NM_004460或序列編號13所示之鹼基序列之人類FAP的基因;或者包含基因庫登錄號NM_007986或序列編號15所示之鹼基序列之小鼠FAP的基因。
又,本發明之編碼FAP之基因例如亦包括:包含編碼如下多肽之DNA之基因,該多肽包含於序列編號13或15、基因庫登錄號NM_004460或基因庫登錄號NM_007986所示之鹼基序列中缺失、置換或附加1個以上之鹼基而成之鹼基序列且具有FAP之功能;包含編碼如下多肽之DNA之基因,該多肽包含與序列編號13或15、基因庫登錄號NM_004460或基因庫登錄號NM_007986所示之鹼基序列具有60%以上之同源性之鹼基序列、較佳為具有80%以上之同源性之鹼基序列、進而較佳為具有95%以上之同源性之鹼基序列且具有FAP之功能;及包含編碼如下多肽之DNA之基因,該多肽包含與包含序列編號13或15、基因庫登錄號NM_004460或基因庫登錄號NM_007986所示之鹼基序列之DNA在嚴格條件下進行雜交之DNA且具有FAP之功能;等。
作為本發明中之FAP之細胞外區,例如可列舉:使用公知之跨膜區預測程式SOSUI(http:// sosui.proteome.bio.tuat.ac.jp/sosuiframe0.html)、TMHMM ver.2(http:// www.cbs.dtu.dk/services/TMHMM-2.0/)或ExPASy Proteomics Server(http://Ca.expasy.org/)等對基因庫登錄號NP_004451所示之人類FAP之胺基酸序列進行預測所得之區域等。具體而言,可列舉:序列編號14或基因庫登錄號NP_004451之第27號~第760號所示之胺基酸序列、序列編號10所示之胺基酸序列。
作為FAP之功能,例如可列舉:藉由該分子所具有之蛋白酶活性而產生之細胞外基質之重建等。
所謂抗體,係源自編碼構成免疫球蛋白之重鏈之可變區及重鏈之恆定區、以及輕鏈之可變區及輕鏈之恆定區的全部或一部分之基因(稱為「抗體基因」)的蛋白質。本發明之抗體亦包括具有任一免疫球蛋白類別及亞型之抗體或抗體片段。
重鏈(H鏈)係指構成免疫球蛋白分子之2種多肽中分子量較大之多肽。重鏈係決定抗體之類別與亞型。IgA、IgD、IgE、IgG及IgM之特徵在於:分別具有α鏈、δ鏈、ε鏈、γ鏈及μ鏈作為重鏈,且重鏈之恆定區為不同之胺基酸序列。輕鏈(L鏈)係指構成免疫球蛋白分子之2種多肽中分子量較小之多肽。於人類抗體之情形時,輕鏈存在κ鏈與λ鏈2種。
可變區(V區)通常係指免疫球蛋白之N末端側之胺基酸序列內所存在之富有多樣性的區域。可變區以外之部分由於採用多樣性較少之結構,故稱為恆定區(C區)。重鏈與輕鏈之各可變區締合而形成抗原結合部位,並決定抗體對抗原之結合特性。以下,將重鏈可變區或輕鏈可變區簡記為VH或VL。
抗原結合部位係抗體中識別抗原並進行結合之部位,且係指與抗原決定基(表位)形成互補立體結構之部位。抗原結合部位在與抗原決定基之間產生較強之分子間相互作用。抗原結合部位係由包含至少3個互補決定區(CDR)之VH及VL所構成。於人類抗體之情形時,VH及VL分別具有3個CDR。將該等CDR自N末端側起依序分別稱為CDR1、CDR2及CDR3。
恆定區中,重鏈恆定區或輕鏈恆定區分別記載為CH或CL。CH係根據作為重鏈之亞型之α鏈、δ鏈、ε鏈、γ鏈及μ鏈來分類。CH係由自N末端側依序排列之CH1區、鉸鏈區、CH2區、CH3區所構成,將CH2區與CH3區一併稱為Fc區。另一方面,CL係分為稱為Cλ鏈及Cκ鏈之2個亞型。
於人類之IgG之情形時,重鏈中,可變區相當於Kabat等人之EU索引(Kabat et al., Sequences of proteins of immunological interest, 1991 Fifth edition)中之第1號直至第117號之胺基酸序列,恆定區相當於第118號以後之胺基酸序列。於人類抗體之輕鏈中,Kabat等人所編號(Kabat編號)之第1號直至第107號之胺基酸序列相當於可變區,第108號以後之胺基酸序列相當於恆定區。
於人類之IgG之情形時,CH1係指具有EU索引中所示之第118號至第215號之胺基酸序列之區域。同樣地,CH2係指具有Kabat等人之EU索引中所示之第231號至第340號之胺基酸序列的區域,CH3係指具有Kabat等人之EU索引中所示之第341號至第447號之胺基酸序列的區域。在CH1與CH2之間存在稱為鉸鏈(hinge)區(以下亦有時記載鉸鏈)之富有柔軟性之胺基酸區域。鉸鏈區係指具有Kabat等人之EU索引中所示之第216號至第230號之胺基酸序列的區域。
CL於人類抗體之κ鏈之情形時,係指具有Kabat編號中所示之第108號至第214號之胺基酸序列之區域,於λ鏈之情形時,係指具有第108號至第215號之胺基酸序列之區域。
單株抗體係保持單一性(monoclonality)之抗體產生細胞所分泌之抗體,且識別單一之表位(亦稱為抗原決定基)。單株抗體分子彼此具有一致之胺基酸序列(1次結構)且採用單一之結構。多株抗體係指不同純系之抗體產生細胞所分泌之抗體分子之集群。寡株抗體係指將複數個不同之單株抗體混合而成之抗體分子之集群。
表位係抗體所識別、結合之抗原之結構部位。作為表位,例如可列舉:單株抗體所識別、結合之單一胺基酸序列、包含胺基酸序列之立體結構、糖鏈所結合之胺基酸序列及包含糖鏈所結合之胺基酸序列之立體結構等。
作為本發明中之單株抗體,可列舉:由融合瘤產生之抗體、及由經包含抗體基因之表現載體轉形而成之轉形體產生之基因重組抗體。
融合瘤例如可藉由如下方式進行製備:製備抗原,自經該抗原免疫之動物取得具有抗原特異性之抗體產生細胞,進而使該抗體產生細胞與骨髓瘤細胞融合。培養該融合瘤、或者將該融合瘤投予至動物中並使該融合瘤腹水癌化,對該培養液或腹水進行分離、純化,藉此可取得所需之單株抗體。作為用抗原進行免疫之動物,只要能夠製作融合瘤,則則可使用任何動物,可適宜地使用小鼠、大鼠、倉鼠及兔等。又,亦可自此種被免疫動物取得具有抗體產生能力之細胞,於在活體外對該細胞實施免疫後,使之與骨髓瘤細胞融合而製作融合瘤。
作為本發明中之基因重組抗體,例如可列舉:重組小鼠抗體、重組大鼠抗體、重組倉鼠抗體、重組兔抗體、人型嵌合抗體(亦稱為嵌合抗體)、人類化抗體(亦稱為CDR移植抗體)及人類抗體等藉由基因重組技術所製造之抗體。基因重組抗體中,可視作為對象之動物種類或目的來決定應用源自何種動物種類之重鏈及輕鏈之可變區以及恆定區。例如於作為對象之動物種類為人類之情形時,可將可變區設為源自人類或小鼠等非人類動物,將恆定區及連接子設為源自人類。
所謂嵌合抗體,係指包含人類以外之動物(非人類動物)之抗體之VH及VL、與人類抗體之CH及CL的抗體。作為非人類動物,只要為小鼠、大鼠、倉鼠及兔等能夠製作融合瘤之動物,則可使用任何動物。嵌合抗體可藉由如下方式製造:自產生單株抗體之源自非人類動物之融合瘤取得編碼VH及VL之cDNA,分別插入至具有編碼人類抗體之CH及CL之DNA的動物細胞用表現載體中而構建嵌合抗體表現載體,將該表現載體導入至動物細胞使之表現。
所謂人類化抗體,係指將非人類動物抗體之VH及VL之CDR移植至人類抗體之VH及VL所對應之CDR所得的抗體。VH及VL之CDR以外之區域稱為架構區(以下,記載為FR)。人類化抗體可藉由如下方式製造:構建編碼包含非人類動物抗體之VH之CDR之胺基酸序列、與任意之人類抗體之VH之FR之胺基酸序列的VH之胺基酸序列之cDNA、及編碼包含非人類動物抗體之VL之CDR之胺基酸序列、與任意之人類抗體之VL之FR之胺基酸序列的VL之胺基酸序列之cDNA,分別插入至具有編碼人類抗體之CH及CL之DNA之動物細胞用表現載體中而構建人類化抗體表現載體,導入至動物細胞使之表現。
人類抗體本來係指天然地存在於人類體內之抗體,但亦包括藉由最近之基因工程、細胞工程、發育工程之技術進步所製作之自人類抗體噬菌體庫及人類抗體產生基因轉殖動物獲得之抗體等。
關於天然地存在於人類體內之抗體,例如可藉由如下方式取得:使EB病毒等感染人類末梢血淋巴細胞以使之永生化,進行選殖,藉此培養產生該抗體之淋巴細胞,自該培養上清液純化該抗體。
人類抗體噬菌體庫係藉由將自人類B細胞製備之抗體基因插入至噬菌體基因,而於噬菌體表面表現Fab、scFv等抗體片段之庫。可自該庫以對於固定有抗原之基質之結合活性作為指標,回收於表面表現具有所需抗原結合活性之抗體片段之噬菌體。該抗體片段可進而藉由基因工程方法,轉化為包含兩條完整之H鏈及兩條完整之L鏈之人類抗體分子。
人類抗體產生基因轉殖動物係指細胞內組入有人類抗體基因之動物。具體而言,例如向小鼠ES細胞導入人類抗體基因,將該ES細胞向小鼠之早期胚胎移植後使個體發育,藉此可製作人類抗體產生基因轉殖小鼠。源自人類抗體產生基因轉殖動物之人類抗體可藉由如下方式製備:使用通常於非人類動物中進行之融合瘤製作法來取得融合瘤,加以培養而使抗體於培養上清液中產生並蓄積。
本發明中,所謂抗CD40抗體,係指特異性地識別CD40之細胞外區且進行結合之單株抗體。又,於本發明中,所謂抗FAP抗體,係指特異性地識別FAP之細胞外區且進行結合之單株抗體。
本發明中,所謂雙專一性抗體,係指分別與2種表位特異性結合之抗體。本發明之雙專一性抗體具有如下結構:含有包含第一抗原結合域之IgG部分及第二抗原結合域,且該IgG部分之重鏈之C末端直接或經由連接子與該第二抗原結合域結合。
本發明中所謂IgG部分,係指包含IgG或Fc部分經改型之IgG的雙專一性抗體之部分結構,具有包含一條輕鏈及一條重鏈之異源二聚物締合2個而成之異源四聚物結構。
上述IgG之重鏈恆定區可為IgG1、IgG2、IgG3、IgG4之任一亞型。又,亦可缺失、附加、置換、及/或插入該等胺基酸序列之一部分。又,可將包含IgG之重鏈之CH1、鉸鏈、CH2及CH3之胺基酸序列之全部或一部分片段適當組合加以使用。又,亦可使該等胺基酸序列部分地缺失、或改換順序加以使用。又,IgG部分所使用之IgG之亞型並無特別限定,較佳為分別將IgG4、IgG4之重鏈恆定區之第228號之Ser殘基置換為Pro,將第235號之Leu殘基置換為Asn而成之IgG4變異體(以下,記載為IgG4PE);或者分別將IgG4之重鏈恆定區之第228號之Ser殘基置換為Pro,將第235號之Leu殘基置換為Asn,且將第409號之Arg殘基置換為Lys而成之IgG4變異體(以下,記載為IgG4PE R409K)。
例如,重鏈之恆定區(自N末端側依序為CH1-鉸鏈-CH2-CH3)較佳為含有包含序列編號61所表示之胺基酸序列之IgG4PE R409K的IgG部分。
本發明中所謂抗原結合域,係具有特異性地識別抗原並進行結合之功能之部分結構。作為抗原結合域,例如可列舉:利用抗體或該抗體片段、配位體或受體等具有對於抗原之結合能力之蛋白質的重組蛋白質或多肽。
本發明中,重組蛋白質、多肽、抗體或該抗體片段或者雙專一性抗體等與CD40或FAP之任一者結合例如可藉由如下方式進行確認:使用公知之免疫學測定法、較佳為螢光細胞染色法等,對表現CD40或FAP之細胞、與重組蛋白質、多肽、抗體或該抗體片段或者雙專一性抗體之結合性進行測定。又,亦可組合使用公知之免疫學測定法[Monoclonal Antibodies - Principles and Practice, Third Edition, Academic Press(1996)、Antibodies - A Laboratory Manual, Cold Spring Harbor Laboratory (1988)、單純系抗體實驗指南、Kodansha Scientific(1987)]等。
本發明之雙專一性抗體中,第一抗原結合域係構成IgG部分之一部分之部分結構。又,第二抗原結合域係與IgG部分之重鏈C末端側直接或經由連接子結合之部分結構。第一及第二抗原結合域之任一者與CD40結合,另一者與FAP結合。本發明之雙專一性抗體中,將第一抗原結合域及第二抗原結合域分別與CD40及FAP結合者稱為CD40-FAP雙專一性抗體。又,將第一抗原結合域及第二抗原結合域分別與FAP及CD40結合者稱為FAP-CD40雙專一性抗體。
本發明之雙專一性抗體可為CD40-FAP雙專一性抗體、FAP-CD40雙專一性抗體中之任一者,較佳為CD40-FAP雙專一性抗體。
本發明中,與FAP結合之抗原結合域只要為分別特異性地識別FAP並進行結合者,則可為任何者。與FAP結合之抗原結合域例如可為如下任一種形態:抗FAP抗體、FAP之配位體及天然存在之相互作用分子等能夠藉由基因重組技術來製作之多肽、蛋白質分子及其片段、以及該蛋白質分子與低分子或天然物之偶聯物等。
作為與FAP結合之抗原結合域之具體例,可列舉:包含抗FAP抗體之CDR之多肽、抗FAP抗體之抗體片段、包含抗FAP抗體之可變區之多肽或者包含與FAP之配位體之結合區之多肽等。作為與FAP結合之抗原結合域,較佳為包含抗FAP抗體之可變區之多肽,進而較佳為抗FAP抗體之Fab。亦有時將抗FAP抗體之Fab稱為抗FAP Fab。
本發明中,與CD40結合之抗原結合域只要為分別特異性地識別CD40並進行結合者,則可為任何者。
作為與CD40結合之抗原結合域之具體例,可列舉:包含抗CD40抗體之CDR之多肽、抗CD40抗體之抗體片段、包含抗CD40抗體之可變區之多肽或者包含與CD40結合之配位體之結合區之多肽等。作為與CD40結合之抗原結合域,較佳為包含抗CD40抗體之可變區之多肽,進而較佳為抗CD40抗體之Fab。亦有時將抗CD40抗體之Fab稱為抗CD40 Fab。
本發明中,作為與CD40結合之抗原結合域之具體例,亦可列舉:源自非促效性抗CD40抗體且分別特異性地識別CD40並進行結合者。
所謂非促效性抗CD40抗體,係指特異性地識別CD40並進行結合,但不具有促效劑活性之抗體。
作為源自非促效性抗CD40抗體且與CD40結合之抗原結合域,例如可列舉:包含非促效性抗CD40抗體之CDR之多肽、非促效性抗CD40抗體之抗體片段、包含非促效性抗CD40抗體之可變區之多肽等。
作為對於CD40之抗原結合域,於使用非促效性抗CD40抗體之情形時,較佳為包含非促效性抗CD40抗體之可變區之多肽,進而較佳為非促效性抗CD40抗體之Fab。
本發明之雙專一性抗體中,於第二抗原結合域為Fab之情形時,IgG部分之重鏈C末端可直接或經由連接子與該Fab之重鏈N末端結合,亦可與輕鏈N末端結合,但較佳為與該Fab之重鏈N末端結合。
於本發明之雙專一性抗體中,IgG部分之重鏈之C末端可與第二抗原結合域直接結合,亦可經由連接子結合。本發明中,所謂連接子,係指使IgG部分之重鏈之C末端與第二抗原結合域結合之化學結構,較佳為多肽。作為連接子,例如可列舉單鏈肽,作為具體例,可列舉:胺基酸序列包含ES、ESKYG、ESKYGPP、GGGGS、或GGGGS之重複序列之單鏈肽。
本發明之雙專一性抗體具有例如圖1所示之結構、即自N末端側依序結合VH1-CH1-鉸鏈-CH2-CH3-VH2-CH1而成之兩條多肽鏈(亦稱為雙專一性抗體之重鏈)及自N末端側結合VL-CL而成之四條多肽鏈(亦稱為雙專一性抗體之輕鏈)。此處,VH1表示重鏈之N末端側之VH,VH2表示C末端側之VH。
雙專一性抗體之兩條重鏈係藉由鉸鏈區之半胱胺酸殘基進行二硫醚鍵結而締合。雙專一性抗體之重鏈與輕鏈係藉由CH1及CL之半胱胺酸殘基進行二硫醚鍵結而締合。雙專一性抗體之兩條重鏈可相同亦可不同,較佳為相同。又,雙專一性抗體之四條輕鏈相互可相同亦可不同,較佳為四條全部相同。又,CL可為Cλ鏈及Cκ鏈之任一者。
作為本發明之雙專一性抗體,例如可列舉:具有CD40促效劑活性之雙專一性抗體。作為本發明之雙專一性抗體,較佳為於不存在FAP分子或表現FAP之細胞(亦稱為FAP表現細胞)之情況下不顯示CD40促效劑活性,於存在FAP分子或表現FAP之細胞之情況下僅顯示CD40促效劑活性之雙專一性抗體。此種雙專一性抗體於如下方面上較佳:由於在存在表現FAP之細胞之癌等病變部或淋巴結中使CD40活化,故不會產生全身性之伴隨CD40之活化之副作用。
人類中,作為表現FAP之細胞,可列舉:腫瘤局部或淋巴結局部之纖維母細胞或腫瘤細胞等。
本發明之雙專一性抗體所具有之CD40促效劑活性係指誘導抗原呈現細胞活化之活性、誘導腫瘤細胞之細胞死亡之活性等,該等活性係藉由雙專一性抗體與細胞上之CD40結合而引起經由該CD40之訊息從而產生。
本發明中,抗體或雙專一性抗體是否具有CD40促效劑活性例如藉由如下方式進行確認:依據Lagresle et al., J Immunol 1995; 154:5746-5756所記載之方法,對B細胞或人類伯奇氏淋巴瘤細胞株Ramos細胞(JCRB細胞庫、細胞編號:JCRB9119)等表現CD40之細胞上的CD95、CD80、CD83、CD86、及/或HLA-ABC之表現量增加進行評價。
本發明中,抗體或雙專一性抗體與表現CD40之細胞結合時,若與陰性對照相比,使CD95、CD80、CD83、CD86、及/或HLA-ABC等之表現量增加,則判定該抗體或該雙專一性抗體具有促效劑活性。另一方面,抗體或雙專一性抗體與表現CD40之細胞結合時,若與陰性對照相比,未使CD95、CD80、CD83、CD86、及/或HLA-ABC等之表現量增加,則判定該抗體或該雙專一性抗體不具有促效劑活性。
作為本發明之雙專一性抗體,具體而言,可列舉:於在表現FAP之細胞之存在下與FAP及CD40結合時,誘導表現CD40之抗原呈現細胞之活化及/或腫瘤細胞之細胞死亡之雙專一性抗體等。
本發明中,CD40拮抗劑活性係指抑制由CD40配位體或CD40促效劑引起之CD40之活化的活性等。例如係指抑制由CD40配位體或CD40促效劑與CD40結合引起之訊息誘導之活性等。
將與CD40結合但不具有CD40拮抗劑活性之抗體稱為非拮抗劑性抗CD40抗體。
抗體之CD40拮抗劑活性例如可藉由如下方式進行確認:由CD40配位體引起之向Ramos細胞等表現CD40之細胞之CD95表現誘導因添加抗體而得到抑制。
本發明之雙專一性抗體可與在同一細胞上表現之CD40及FAP結合,亦可與在兩個不同之細胞上表現之CD40及FAP結合。
本發明中,將一分子之雙專一性抗體所具有之與某一抗原之結合區的數稱為結合價數。例如於本發明中,於一分子之雙專一性抗體具有與CD40結合之抗原結合域及與FAP結合之抗原結合域各兩個之情形時,可謂該雙專一性抗體以二價分別與CD40及FAP結合。
本發明之雙專一性抗體可藉由既有之製作技術([Nature Protocols, 9, 2450-2463 (2014)]、國際公開第1998/050431號、國際公開第2001/7734號、國際公開第2002/002773號及國際公開第2009/131239號)等進行製作。
本發明之雙專一性抗體可於IgG部分所包含之2個重鏈各自之C末端各具有1個第二抗原結合域,亦可僅於一個重鏈之C末端具有1個第二抗原結合域,但較佳為於2個重鏈各自之C末端各具有1個第二抗原結合域。於2個重鏈各自之C末端各具有1個第二抗原結合域之情形時,該等可相同亦可不同,但較佳為相同之第二抗原結合域。
本發明之雙專一性抗體亦包括:於構成本發明之雙專一性抗體之胺基酸序列中缺失、附加、置換或插入1個以上之胺基酸殘基且具有與上述雙專一性抗體相同之活性的雙專一性抗體。
供缺失、置換、插入及/或附加之胺基酸之數為1個以上,其數量並無特別限定,係藉由Molecular Cloning, The Second Edition, Cold Spring Harbor Laboratory Press (1989)、Current Protocols in Molecular Biology, John Wiley & Sons (1987-1997)、Nucleic Acids Research, 10, 6487 (1982)、Proc. Natl. Acad. Sci., USA, 79, 6409 (1982)、Gene, 34, 315 (1985)、Nucleic Acids Research, 13, 4431 (1985)、Proc. Natl. Acad. Sci USA, 82, 488 (1985)等所記載之定點突變導入法等周知之技術,可缺失、置換、插入或附加之程度之數。例如通常為1~數十個、較佳為1~20個、更佳為1~10個、進而較佳為1~5個。
上述所謂於本發明之雙專一性抗體之胺基酸序列中缺失、附加、置換或插入1個以上之胺基酸殘基係表示如下情況。意指於同一序列中之任意且1個或複數個胺基酸序列中缺失、置換、插入或附加有1個或複數個胺基酸殘基。又,亦存在缺失、置換、插入或附加同時產生之情形,且亦存在供置換、插入或附加之胺基酸殘基為天然型與非天然型中之任一者之情形。
作為天然型胺基酸殘基,例如可列舉:L-丙胺酸、L-天冬醯胺、L-天冬胺酸、L-麩醯胺、L-麩胺酸、甘胺酸、L-組胺酸、L-異白胺酸、L-白胺酸、L-離胺酸、L-精胺酸、L-甲硫胺酸、L-苯基丙胺酸、L-脯胺酸、L-絲胺酸、L-蘇胺酸、L-色胺酸、L-酪胺酸、L-纈胺酸及L-半胱胺酸等。
於以下表示能夠相互置換之胺基酸殘基之較佳例。同一群中所含有之胺基酸殘基能夠相互置換。
A群:白胺酸、異白胺酸、甘白胺酸、纈胺酸、正纈胺酸、丙胺酸、2-胺基丁酸、甲硫胺酸、O-甲基絲胺酸、第三丁基甘胺酸、第三丁基丙胺酸、環己基丙胺酸
B群:天冬胺酸、麩胺酸、異天冬胺酸、異麩胺酸、2-胺基己二酸、2-胺基辛二酸
C群:天冬醯胺、麩醯胺
D群:離胺酸、精胺酸、鳥胺酸、2,4-二胺基丁酸、2,3-二胺基丙酸
E群:脯胺酸、3-羥基脯胺酸、4-羥基脯胺酸
F群:絲胺酸、蘇胺酸、高絲胺酸
G群:苯基丙胺酸、酪胺酸
本發明之雙專一性抗體亦含有包含經轉譯後修飾之任何胺基酸殘基的抗體。作為轉譯後修飾,例如可列舉:多肽之N末端中之麩醯胺殘基向焦麩胺(pyroGlu)之置換等[Beck et al, Analytical Chemistry, 85, 715-736(2013)]。
作為本發明之雙專一性抗體,例如亦可列舉:第一抗原結合域及第二抗原結合域之任一者為源自非促效性抗CD40抗體之抗CD40 Fab的雙專一性抗體、第一抗原結合域及上述第二抗原結合域之任一者為抗FAP Fab之雙專一性抗體。
又,作為本發明之雙專一性抗體,例如可列舉:第一抗原結合域為抗CD40 Fab且上述第二抗原結合域為抗FAP Fab之雙專一性抗體、第一抗原結合域為抗FAP Fab且第二抗原結合域為抗CD40 Fab之雙專一性抗體。
作為本發明之雙專一性抗體,具體而言,例如可列舉下述(1)~(4)所記載之雙專一性抗體等。
(1)包含源自抗CD40抗體之抗CD40 Fab之IgG部分之重鏈之C末端直接與抗FAP Fab結合而成的與人類CD40及人類FAP結合之雙專一性抗體。
(2)包含抗FAP Fab之IgG部分之重鏈之C末端直接與源自抗CD40抗體之抗CD40 Fab結合而成的與人類CD40及人類FAP結合之雙專一性抗體。
(3)包含源自非促效性抗CD40抗體之抗CD40 Fab之IgG部分之重鏈之C末端直接與抗FAP Fab結合而成的與人類CD40及人類FAP結合之雙專一性抗體。
(4)包含抗FAP Fab之IgG部分之重鏈之C末端直接與源自非促效性抗CD40抗體之抗CD40 Fab結合而成的與人類CD40及人類FAP結合之雙專一性抗體。
此處,IgG部分之重鏈之恆定區(自N末端側依序為CH1-鉸鏈-CH2-CH3)較佳為包含序列編號61所表示之胺基酸序列。又,作為本發明之雙專一性抗體或該雙專一性抗體片段,較佳為上述(1)及(3)。
上述(1)~(4)之雙專一性抗體中,較佳為包含如下抗CD40 Fab之雙專一性抗體,該抗CD40 Fab包括含有分別包含序列編號24~26所表示之胺基酸序列之互補決定區(CDR)1~3之VH及含有分別包含序列編號19~21所表示之胺基酸序列之CDR1~3之VL,更佳為包含如下抗CD40 Fab之雙專一性抗體,該抗CD40 Fab含有包含序列編號23所表示之胺基酸序列之VH及包含序列編號18所表示之胺基酸序列之VL。
又,上述(1)~(4)之雙專一性抗體中,較佳為包含如下抗FAP Fab之雙專一性抗體,該抗FAP Fab含有包含選自以下(i)~(v)中之任一個、較佳為選自(iii)~(v)中之任一個、更佳為(iii)之重鏈CDR1~3之VH、及含有分別包含序列編號19~21所表示之胺基酸序列之CDR1~3之VL。
(i)分別包含序列編號33~35所表示之胺基酸序列之重鏈CDR1~3、
(ii)分別包含序列編號38~40所表示之胺基酸序列之重鏈CDR1~3、
(iii)分別包含序列編號43~45所表示之胺基酸序列之重鏈CDR1~3、
(iv)分別包含序列編號48~50所表示之胺基酸序列之重鏈CDR1~3及
(v)分別包含序列編號53~55所表示之胺基酸序列之重鏈CDR1~3。
進而,上述(1)~(4)之雙專一性抗體中,更佳為包含如下抗FAP Fab之雙專一性抗體,該抗FAP Fab含有包含選自序列編號32、37、42、47、52、75、77、79、81及83、較佳為選自序列編號42、47及75之任一者所表示之胺基酸序列、更佳為序列編號42所表示之胺基酸序列之VH、及包含序列編號18所表示之胺基酸序列之VL。
上述(1)~(4)之雙專一性抗體中,進而較佳為包含如下抗CD40 Fab以及如下抗FAP Fab之雙專一性抗體,該抗CD40 Fab包括含有分別包含序列編號24~26所表示之胺基酸序列之CDR1~3之VH、及含有分別包含序列編號19~21所表示之胺基酸序列之CDR1~3之VL,該抗FAP Fab含有包含選自以下(i)~(v)中之任一個、較佳為選自(iii)~(v)中之任一個、更佳為(iii)之重鏈CDR1~3之VH、及含有分別包含序列編號19~21所表示之胺基酸序列之CDR1~3之VL。
(i)分別包含序列編號33~35所表示之胺基酸序列之重鏈CDR1~3、
(ii)分別包含序列編號38~40所表示之胺基酸序列之重鏈CDR1~3、
(iii)分別包含序列編號43~45所表示之胺基酸序列之重鏈CDR1~3、
(iv)分別包含序列編號48~50所表示之胺基酸序列之重鏈CDR1~3及
(v)分別包含序列編號53~55所表示之胺基酸序列之重鏈CDR1~3。
上述(1)~(4)之雙專一性抗體中,進一步較佳為包含如下抗CD40 Fab以及如下抗FAP Fab之雙專一性抗體,該抗CD40 Fab含有包含序列編號23所表示之胺基酸序列之VH、及包含序列編號18所表示之胺基酸序列之VL,該抗FAP Fab含有包含選自序列編號32、37、42、47、52、75、77、79、81及83、較佳為選自序列編號42、47及75之任一者所表示之胺基酸序列、更佳為序列編號42所表示之胺基酸序列之VH、及包含序列編號18所表示之胺基酸序列之VL。
又,作為本發明之雙專一性抗體,可列舉包含2條重鏈及4條輕鏈之雙專一性抗體,該重鏈包含選自序列編號65、67、69、71、73、85、87、89、91、93、95、97、99、101、103、105、107、109及111、較佳為選自序列編號69、71及85之任一者所表示之胺基酸序列、更佳為序列編號69所表示之胺基酸序列,該輕鏈含有包含序列編號18所表示之胺基酸序列之VL。
又,本發明之雙專一性抗體亦包含如下雙專一性抗體,該雙專一性抗體包含重鏈及4條輕鏈,該重鏈包含與選自序列編號65、67、69、71、73、85、87、89、91、93、95、97、99、101、103、105、107、109及111之任一者所表示之胺基酸序列分別至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、或至少99%同源之胺基酸序列,該輕鏈包含與序列編號18所表示之胺基酸序列至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、或至少99%同源之胺基酸序列。
作為本發明之雙專一性抗體之例,可列舉:Ct-R1090-Fa018、Ct-R1090-Fa1007、Ct-R1090-Fa1013、Ct-R1090-Fa1014、Ct-R1090-Fa1046、Ct-R1090-Fa1046-GTS、Ct-R1090-Fa1046-FTS、Ct-R1090-Fa1046-KTS、Ct-R1090-Fa1046-WTS、Ct-R1090-Fa1046-TTS、Ct-Fa018-R1090、Ct-Fa1007-R1090、Ct-Fa1013-R1090、Ct-Fa1014-R1090、Ct-Fa1046-GTS-R1090、Ct-Fa1046-FTS-R1090、Ct-Fa1046-KTS-R1090、Ct-Fa1046-WTS-R1090及Ct-Fa1046-TTS-R1090。其中,較佳為Ct-R1090-Fa1013、Ct-R1090-Fa1014及Ct-R1090-Fa1046-GTS,更佳為Ct-R1090-Fa1013。
本發明之雙專一性抗體亦包含具有效應物活性之抗體。
所謂效應物活性,係指經由抗體之Fc區所引起之抗體依賴性之細胞毒殺活性,例如可列舉:抗體依賴性細胞毒殺活性(Antibody-Dependent Cellular Cytotoxicity activity;ADCC活性)、補體依賴性細胞毒殺活性(Complement-Dependent Cytotoxicity activity;CDC活性)、利用巨噬細胞或樹狀細胞等吞噬細胞之抗體依賴性巨噬活性(Antibody-dependent cellular phagocytosis activity;ADCP活性)及助噬作用效果等。
本發明中,ADCC活性及CDC活性可使用公知之測定方法[Cancer Immunol. Immunother., 36, 373(1993)]進行測定。
所謂ADCC活性,係指與靶細胞上之抗原結合之抗體經由抗體之Fc區與免疫細胞之Fc受體結合,藉此使免疫細胞(自然殺手細胞等)活化而殺傷靶細胞之活性。
Fc受體(FcR)係與抗體之Fc區結合之受體,藉由抗體之結合而誘發各種效應物活性。各FcR係與抗體之亞型對應,IgG、IgE、IgA、IgM分別與FcγR、FcεR、FcαR、FcμR特異性地結合。進而於FcγR中存在FcγRI(CD64)、FcγRII(CD32)及FcγRIII(CD16)之亞型,各亞型存在FcγRIA、FcγRIB、FcγRIC、FcγRIIA、FcγRIIB、FcγRIIC、FcγRIIIA及FcγRIIIB之同功異型物。該等不同之FcγR存在於不同之細胞上[Annu. Rev. Immunol. 9:457-492 (1991)]。於人類中,FcγRIIIB係於嗜中性球中特異性地表現,、FcγRIIIA係於單核球、自然殺手細胞(NK細胞)、巨噬細胞及一部分之T細胞中表現。經由抗體向FcγRIIIA之結合而誘發NK細胞依賴性之ADCC活性。
所謂CDC活性,係指與靶細胞上之抗原結合之抗體使血液中之包含補體相關蛋白質群之一系列級聯(補體活化途徑)活化而殺傷靶細胞的活性。又,藉由利用補體之活化而產生之蛋白質片段,而誘發免疫細胞之趨化及活化。CDC活性之級聯係藉由首先使C1q與Fc區結合,繼而與2個絲胺酸蛋白酶即C1r及C1s結合,而形成C1複合體從而開始。
本發明之雙專一性抗體對於抗原表現細胞之CDC活性、或ADCC活性可藉由公知之測定方法[Cancer Immunol. Immunother., 36, 373 (1993)]進行評價。
作為控制本發明之雙專一性抗體之效應物活性之方法,已知有:控制α-1,6結合至N-結合複合型糖鏈之還原末端所存在之N-乙醯葡糖胺(GlcNAc)之海藻糖(亦稱為核心海藻糖)之量的方法,該N-結合複合型糖鏈與IgG抗體部分之Fc區(包含CH2及CH3區之恆定區)之第297號之天冬醯胺(Asn)結合(國際公開第2005/035586號、國際公開第2002/31140號及國際公開第00/61739號);或者藉由改變抗體之Fc區之胺基酸殘基來進行控制之方法(國際公開第00/42072號)等。
藉由控制附加於雙專一性抗體之海藻糖之量,可增加或降低抗體之ADCC活性。例如,作為使結合至與抗體之Fc結合之N-結合複合型糖鏈之海藻糖的含量降低之方法,藉由使用缺失有α1,6-海藻糖轉移酶基因之宿主細胞來使雙專一性抗體表現,可取得具有較高之ADCC活性之雙專一性抗體。另一方面,作為使結合至與雙專一性抗體之Fc結合之N-結合複合型糖鏈之海藻糖的含量增加之方法,藉由使用導入有α1,6-海藻糖轉移酶基因之宿主細胞來使抗體表現,可取得具有較低之ADCC活性之雙專一性抗體。
又,藉由改型雙專一性抗體之Fc區之胺基酸殘基,可增加或降低ADCC活性或CDC活性。例如藉由使用美國專利申請公開第2007/0148165號說明書中記載之Fc區之胺基酸序列,可增加雙專一性抗體之CDC活性。又,藉由進行美國專利第6,737,056號說明書、美國專利第7,297,775號說明書或美國專利第7,317,091號說明書等中所記載之胺基酸改變,可增加ADCC活性或CDC活性,亦可降低ADCC活性或CDC活性。
進而,藉由將上述方法組合,亦可取得效應物活性得到控制之雙專一性抗體。
本發明之雙專一性抗體之穩定性可藉由測定純化過程中或在一定條件下保存之樣品中所形成之凝集體(寡聚物)量來進行評價。在與對照組相比,於本發明之雙專一性抗體中,相同條件下之凝集體量降低之情形時,評價為本發明之雙專一性抗體之穩定性高於對照組。凝集體量可藉由使用包含凝膠過濾層析法之適當層析法將所凝集之抗體與未凝集之抗體進行分離而進行測定。
本發明之雙專一性抗體之生產細胞之生產性可藉由對培養液中自該抗體產生細胞產生之抗體量進行測定來評價。更具體而言,可藉由利用HPLC法或ELISA法等適當之方法對自培養液中去除產生細胞後所得之培養上清液中所含之抗體量進行測定來評價。
本發明中,所謂抗體片段,係免疫球蛋白之片段,且係包含抗原結合部位並具有對於抗原之抗原結合活性之蛋白質。例如可列舉:Fab、Fab'、F(ab')2
、scFv、雙功能抗體、dsFv、VHH或包含CDR之肽等。
Fab係利用蛋白分解酵素木瓜酶對IgG抗體進行處理所獲得之片段中(在H鏈之第224號之胺基酸殘基被切斷)、H鏈之N末端側約一半與L鏈整體以雙硫鍵(S-S鍵)結合之分子量約5萬的具有抗原結合活性之抗體片段。將包含VH及CH1之Fab之H鏈記載為VH-CH1。又,將包含VL及CL之Fab之L鏈記載為VL-CL。將源自抗CD40抗體之Fab記載為抗CD40 Fab,將源自抗FAP抗體之Fab記載為抗FAP Fab。
F(ab')2
係利用蛋白分解酵素胃蛋白酶對IgG進行處理所獲得之片段中(在H鏈之第234號之胺基酸殘基被切斷),較經由鉸鏈區之S-S鍵而結合有Fab者稍大、分子量約10萬之具有抗原結合活性的抗體片段。
Fab'係上述F(ab')2
之鉸鏈區之S-S鍵被切斷之分子量約5萬之具有抗原結合活性的抗體片段。
scFv係使用12個殘基以上之適當之肽連接子(P)將一條VH與一條VL連結而成之VH-P-VL或VL-P-VH多肽,且係具有抗原結合活性之抗體片段。
雙功能抗體係抗原結合特異性相同或不同之scFv形成了二聚物之抗體片段,且係具有對於相同抗原之2價抗原結合活性、或對於不同抗原分別具有特異性抗原結合活性之抗體片段。
dsFv係指使將VH及VL中之各1個胺基酸殘基置換為半胱胺酸殘基而成之多肽經由該半胱胺酸殘基間之S-S鍵進行結合而成者。
VHH(亦稱為奈米抗體)係指重鏈抗體之重鏈之可變區(variable domain of heavy chain of heavy chain antibody)(VHH抗體)中之重鏈可變區,且可於不存在其他多肽之情況下與抗原結合。
VHH抗體係存在於羊駝等駝科動物及鯊魚等軟骨魚中之抗體,無輕鏈及CH1,僅包含重鏈。
包含CDR之肽包含VH或VL之CDR之至少1個區域。包含複數個CDR之肽可藉由使CDR彼此直接或經由適當之肽連接子結合而製作。包含CDR之肽可藉由如下方式表現並製造:構建編碼本發明之雙專一性抗體之VH及VL之CDR之DNA,將該DNA插入至原核生物用表現載體或真核生物用表現載體,將該表現載體向原核生物或真核生物導入。又,包含CDR之肽亦可藉由Fmoc法或tBoc法等化學合成法來製造。
本發明之雙專一性抗體亦可使用包含胺基酸殘基改變之Fc區,該胺基酸殘基改變係以抗體之效應物活性之增強或缺損、抗體之穩定化、及血中半衰期之控制為目的。
本發明之雙專一性抗體包含使放射性同位素、低分子之藥劑、高分子之藥劑、蛋白質或抗體醫藥等以化學或基因工程方式與本發明之雙專一性抗體結合而成的雙專一性抗體之衍生物。
本發明中之雙專一性抗體之衍生物可藉由如下方式進行製造:利用化學手法[抗體工程入門,地人書館(1994)]使放射性同位素、低分子之藥劑、高分子之藥劑、免疫活化劑、蛋白質或抗體醫藥等與本發明之雙專一性抗體之H鏈或L鏈之N末端側或C末端側、該雙專一性抗體中之糖鏈等結合。
又,本發明中之雙專一性抗體之衍生物可藉由如下基因工程方法進行製造:使編碼本發明之雙專一性抗體之DNA、與編碼所需蛋白質或抗體醫藥之DNA連結並插入至表現載體,將該表現載體導入至適當之宿主細胞使之表現。
作為放射性同位素,例如可列舉:111
In、131
I、125
I、90
Y、64
Cu、99
Tc、77
Lu或211
At等。放射性同位素可藉由氯胺T法等直接結合至抗體。又,亦可使抗體與使放射性同位素螯合之物質結合。作為螯合劑,例如可列舉:1-異硫氰酸酯基苄基-3-甲基二伸乙基三胺五乙酸(MX-DTPA)等。
作為低分子之藥劑,例如可列舉:烷基化劑、亞硝基脲劑、代謝拮抗劑、抗生素、植物鹼、拓樸異構酶抑制劑、激素治療劑、激素拮抗劑、芳香酶抑制劑、P糖蛋白抑制劑、鉑錯合物衍生物、M期抑制劑或激酶抑制劑等抗癌劑[臨床腫瘤學,癌與化學療法社(1996)];氫化可體松或潑尼松等類固醇劑;阿司匹林或吲哚美辛等非類固醇劑;硫代蘋果酸金或青黴胺等免疫調節劑;環磷醯胺或硫唑嘌呤等免疫抑制劑;或者如馬來酸氯苯那敏或氯苯苄咯(Tavegil)之抗組織胺劑等抗炎症劑[炎症與抗炎症療法,醫齒藥出版股份有限公司(1982)]等。
作為抗癌劑,例如可列舉:阿密磷定(阿米福汀(Ethyol))、順鉑、達卡巴(DTIC)、放線菌素、二氯甲基二乙胺(氮芥)、鏈佐星、環磷醯胺、異環磷醯胺、卡莫司汀(BCNU)、環己亞硝脲(CCNU)、多柔比星(阿德力黴素)、表柔比星、吉西他濱(鹽酸吉西他濱)、道諾紅菌素、甲基苄肼)、絲裂黴素、阿糖胞苷、依託泊苷、5-氟尿嘧啶、氟尿嘧啶、長春花鹼、長春新鹼、博萊黴素、道諾黴素、培洛黴素、雌莫司汀、紫杉醇(泰克索)、歐洲紫杉醇(多西他賽)、阿地白介素、門冬醯胺酶、白消安、卡鉑、奧沙利鉑、奈達帕汀、克拉屈濱、喜樹鹼、10-羥基-7-乙基-喜樹鹼(SN38)、氟尿苷、氟達拉濱、羥基脲、依達比星、巰乙磺酸鈉、伊立替康(CPT-11)、拓樸替康、米托蒽醌、托泊替康、亮丙瑞林、甲地孕酮、美法侖、巰嘌呤、羥基脲、光輝黴素、密妥坦、培門冬酶、噴司他丁、哌泊溴烷、鏈佐星、他莫昔芬、戈舍瑞林、亮丙瑞林、氟他胺、替尼泊甙、睾內脂、硫鳥嘌呤、噻替派、尿嘧啶氮芥、長春瑞濱、氯芥苯丁酸、氫化可體松、潑尼松龍、甲基潑尼松龍、長春地辛、尼莫司汀、司莫司汀、卡培他濱、拓優得、阿紮胞苷、UFT、奧沙利鉑、吉非替尼(易瑞沙)、伊馬替尼(STI571)、厄洛替尼、類FMS酪胺酸激酶(FMS-like tyrosine kinase 3)(Flt3)抑制劑、血管內皮生長因子受體(vascular endothelial growth factor receptor)(VEGFR)抑制劑、纖維母細胞生長因子受體(fibroblast growth factor receptor)(FGFR)抑制劑、埃羅替尼(Tarceva)等表皮生長因子受體(epidermal growth factor receptor)(EGFR)抑制劑、根赤殼菌素、17-烯丙基胺基-17-去甲氧基格爾德黴素、雷帕黴素、安吖啶、全反維他命A酸、沙利竇邁、來那度胺、阿那曲唑、法倔唑、來曲唑、依西美坦、硫代蘋果酸金、D-青黴胺、布西拉明、硫唑嘌呤、咪唑立賓、環孢靈、雷帕黴素、氫化可體松、蓓薩羅丁(貝沙羅汀)、他莫昔芬、地塞米松、助孕素類、雌激素類、阿那曲唑(阿納托唑)、亮丙瑞林、阿司匹林、吲哚美辛、塞來昔布、硫唑嘌呤、青黴胺、硫代蘋果酸金、馬來酸氯苯那敏、馬來酸氯苯吡胺、氯雷他定、維生素A酸、蓓薩羅丁、砷、硼替佐米、別嘌呤醇、卡奇黴素、替伊莫單抗、貝沙羅汀、Ozogamine、克拉紅黴素、甲醯四氫葉酸、酮康唑、氨魯米特、蘇拉明、美登素類化合物或其衍生物等。
作為使低分子之藥劑與本發明之雙專一性抗體鍵結之方法,例如可列舉:經由戊二醛使藥劑與該抗體之胺基間鍵結之方法、或者經由水溶性碳二醯亞胺使藥劑之胺基與該抗體之羧基鍵結之方法等。
作為高分子之藥劑,例如可列舉:聚乙二醇(PEG)、白蛋白、葡聚糖、聚氧乙烯、苯乙烯馬來酸共聚物、聚乙烯吡咯啶酮、吡喃共聚物、或羥丙基甲基丙烯醯胺等。藉由使該等高分子化合物與本發明之雙專一性抗體結合,而期待(1)對於化學、物理或生物上之各種因子之穩定性之提高、(2)血中半衰期之顯著延長、或者(3)免疫原性之消失或抗體產生之抑制等效果[Bioconjugates醫藥品,廣川書店(1993)]。
例如,作為使PEG與本發明之雙專一性抗體結合之方法,可列舉:與PEG化修飾試劑進行反應之方法等[Bioconjugates醫藥品,廣川書店(1993)]。作為PEG化修飾試劑,可列舉:對於離胺酸之ε-胺基之修飾劑(日本專利特開昭61-178926號公報)、對於天冬胺酸及麩胺酸之羧基之修飾劑(日本專利特開昭56-23587號公報)、或者對於精胺酸之胍基之修飾劑(日本專利特開平2-117920號公報)等。
作為免疫活化劑,可為作為免疫佐劑為人所知之天然物,作為具體例,使免疫亢進之藥劑可列舉:β(1→3)葡聚糖(例如,香菇多糖或裂褶菌多糖)、或α半乳糖苷基神經醯胺(KRN7000)等。
作為蛋白質,例如可列舉:使NK細胞、巨噬細胞或嗜中性球等免疫活性(適格)細胞活化之細胞激素或生長因子或者毒素蛋白質等。
作為細胞激素或生長因子,例如可列舉:干擾素(以下,記為IFN)-α、IFN-β、IFN-γ、介白素(以下,記為IL)-2、IL-12、IL-15、IL-18、IL-21、IL-23、顆粒球菌落刺激因子(G-CSF)、顆粒球/巨噬細胞菌落刺激因子(GM-CSF)或巨噬細胞菌落刺激因子(M-CSF)等。
作為毒素蛋白質,例如可列舉:離胺酸、白喉毒素或ONTAK等,亦包括為了調節毒性而向蛋白質導入突變而成之蛋白質毒素。
與蛋白質或抗體醫藥之融合抗體可藉由如下方式表現並製造:使編碼本發明之雙專一性抗體之cDNA與編碼蛋白質之cDNA連結而構建編碼融合抗體之DNA,將該DNA插入至原核生物或真核生物用表現載體,將該表現載體向原核生物或真核生物導入。
於將上述雙專一性抗體之衍生物用作檢測方法、測定方法、檢測用試劑、測定用試劑或診斷藥的情形時,作為與本發明之雙專一性抗體結合之藥劑,可列舉:通常之免疫學檢測或測定方法中所使用之標記物。作為標記物,例如可列舉:鹼性磷酸酶、過氧化酶或螢光素酶等酶;吖啶鎓酯或咯吩等發光物質;或者螢光異硫氰酸鹽(FITC)或異硫氰酸四甲基羅丹明(RITC)、Alexa(註冊商標)Fluor 488、R-紅藻素(R-PE)等螢光物質等。
本發明中包含具有CDC活性或ADCC活性等細胞毒殺活性之雙專一性抗體。本發明之雙專一性抗體對於抗原表現細胞之CDC活性或ADCC活性可藉由公知之測定方法[Cancer Immunol. Immunother., 36, 373 (1993)]進行評價。
又,本發明係關於一種包含特異性地識別CD40及FAP並進行結合之雙專一性抗體之組合物、或者含有該雙專一性抗體作為有效成分之與CD40及FAP之至少一者相關之疾病之治療藥。
作為與CD40及FAP之至少一者相關之疾病,只要為與CD40及FAP之至少一者有關之疾病,則可為任何者,例如可列舉惡性腫瘤及癌症等。
本發明中,作為惡性腫瘤及癌症,例如可列舉:大腸癌、結腸直腸癌、肺癌、乳癌、神經膠質瘤、惡性黑色素瘤(黑色素瘤)、甲狀腺癌、腎細胞癌、白血病、淋巴瘤、T細胞淋巴瘤、胃癌、胰腺癌、子宮頸癌、子宮內膜癌、卵巢癌、膽管癌、食道癌、肝臟癌、頭頸癌、皮膚癌、尿路癌、膀胱癌、前列腺癌、絨毛膜癌、咽頭癌、喉頭癌、間皮瘤、胸膜瘤、雄胚瘤、子宮內膜增生、子宮內膜症、胚組織瘤、纖維肉瘤、卡波西肉瘤、血管瘤、海綿狀血管瘤、血管母細胞瘤、視網膜胚細胞瘤、星狀細胞瘤、神經纖維瘤、少突膠質細胞瘤、神經管胚細胞瘤、神經胚細胞瘤、神經膠質瘤、橫紋肌肉瘤、神經膠母細胞瘤、骨原性肉瘤、平滑肌肉瘤及威爾姆斯瘤等。
含有本發明之雙專一性抗體、或該等之衍生物之治療藥可為僅包含作為有效成分之該雙專一性抗體、或其衍生物者,較佳為通常與藥理學上所容許之1種以上之載體一起混合,而以藉由製劑學之技術領域中所公知之任意方法所製造之醫藥製劑的形式提供。
投予路徑較佳為使用治療時最具效果者,例如可列舉:經口投予、或者口腔內、呼吸道內、直腸內、皮下、肌內或靜脈內等非經口投予。其中,較佳為靜脈內投予。
作為投予形態,例如可列舉:噴霧劑、膠囊劑、錠劑、散劑、顆粒劑、糖漿劑、乳劑、栓劑、注射劑、軟膏或貼劑等。
投予量或投予次數係根據設為目標之治療效果、投予方法、治療期間、年齡、及體重等而不同,通常成人每天為10 μg/kg~10 mg/kg。
進而,本發明係關於一種含有本發明之雙專一性抗體之CD40及FAP之至少一者的免疫學檢測用或測定用試劑、或者與CD40及FAP之至少一者相關之疾病之診斷藥。又,本發明係關於一種使用本發明之雙專一性抗體之CD40及FAP之至少一者的免疫學檢測用或測定用方法;與CD40及FAP之至少一者相關之疾病之治療方法;或者與CD40及FAP之至少一者相關之疾病、較佳為CD40及FAP之表現細胞所參與之疾病之診斷方法。
作為本發明中檢測或測定CD40及FAP之至少一者之量之方法,可列舉任意之公知方法。例如可列舉:免疫學檢測或測定方法等。
所謂免疫學檢測或測定方法,係使用施加了標記之抗原或抗體,對抗體量或抗原量進行檢測或測定之方法。作為免疫學檢測或測定方法,例如可列舉:放射免疫測定法(RIA)、酶免疫測定法(EIA或ELISA)、螢光免疫測定法(FIA)、發光免疫測定法(luminescent immunoassay)、西方點墨法或物理化學手法等。
藉由使用本發明之雙專一性抗體檢測或測定表現CD40及FAP之至少一者之細胞,可診斷與CD40及FAP之至少一者相關之疾病、較佳為CD40及FAP之表現細胞所參與之疾病。
於檢測表現CD40及FAP之至少一者之細胞時,可使用公知之免疫學檢測法。作為公知之免疫學檢測法,可列舉:免疫沈澱法、免疫細胞染色法、免疫組織染色法或螢光抗體染色法等。又,螢光抗體染色法例如可使用FMAT8100HTS系統(Applied Biosystems公司製造)等。
本發明中作為成為檢測或測定CD40及FAP之至少一者之對象的生物樣本,例如為組織細胞、血液、血漿、血清、胰液、尿、糞便、組織液或培養液等,只要有可能包含表現CD40及FAP之至少一者之細胞,則無特別限定。
含有本發明之雙專一性抗體、或該等之衍生物之診斷藥亦可視目標之診斷法而含有用以進行抗原抗體反應之試劑、該反應之檢測用試劑。作為用以進行抗原抗體反應之試劑,例如可列舉緩衝劑、鹽等。
作為檢測用試劑,例如可列舉:與上述雙專一性抗體、或該等之衍生物結合之經標記之二次抗體、或者與標記對應之受質等通常之免疫學檢測或測定方法所使用之試劑。
以下,關於本發明之雙專一性抗體之製作方法、該雙專一性抗體之活性評價方法、以及使用該雙專一性抗體之疾病之治療方法及診斷方法,具體地進行記載。
1.單株抗體之製作方法
本發明中之單株抗體之製造方法包括下述作業步驟。即,(1)純化用作免疫原之抗原及製作抗原於細胞表面過度表現之細胞之至少一步驟;(2)用抗原免疫動物後,採集血液對其抗體效價進行檢驗以決定摘除脾臟等之時期,從而製備抗體產生細胞之步驟;(3)製備骨髓瘤細胞(myeloma);(4)使抗體產生細胞與骨髓瘤進行細胞融合;(5)篩選產生目標抗體之融合瘤群;(6)自融合瘤群分離(選殖)單純系(monoclonal)細胞;(7)視情形培養用以大量製造單株抗體之融合瘤、或飼養移植有融合瘤之動物;(8)對如此製造之單株抗體之生理活性及其抗原結合特異性進行研究、或者檢驗作為標記試劑之特性;等。
以下,按照上述步驟,對用以製作本發明中之與CD40及FAP結合之雙專一性抗體的與CD40結合之單株抗體及與FAP結合之單株抗體的製作方法進行詳述。該抗體之製作方法並不限於此,例如亦可使用脾臟細胞以外之抗體產生細胞及骨髓瘤。
(1)抗原之純化
表現CD40或FAP之細胞可藉由將包含編碼CD40或FAP全長或其部分長度之cDNA的表現載體導入至大腸桿菌、酵母、昆蟲細胞或動物細胞等中而獲得。又,可自大量地表現CD40及FAP之至少一者之各種人類腫瘤培養細胞或人類組織等純化CD40及FAP之至少一者並用作抗原。又,亦可將該腫瘤培養細胞或該組織等直接用作抗原。進而,亦可藉由Fmoc法或tBoc法等化學合成法製備具有CD40或FAP之部分序列之合成肽用於抗原。
本發明中所使用之CD40及FAP可使用Molecular Cloning, A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press (1989)或Current Protocols In Molecular Biology, John Wiley & Sons (1987-1997)等所記載之方法等,例如藉由以下之方法使編碼CD40或FAP之DNA於宿主細胞中表現而進行製造。
藉由將包含編碼CD40或FAP之部分之全長cDNA插入至適當之表現載體之啟動子之下游,從而製作重組載體。亦可使用基於全長cDNA所製備之包含編碼多肽之部分之適當長度之DNA片段代替上述全長cDNA。繼而,將所獲得之該重組載體導入至適於該表現載體之宿主細胞,藉此可獲得生產CD40或FAP之轉形體。
作為表現載體,只要可實現於使用之宿主細胞中自主複製或向染色體中組入,且於可轉錄編碼CD40或FAP之DNA之位置含有適當啟動子,則可使用任一種。
作為宿主細胞,例如為大腸桿菌等屬於艾氏菌屬等之微生物、酵母、昆蟲細胞或動物細胞等,只要可表現目標之基因,則可使用任一種。
於使用大腸桿菌等原核生物作為宿主細胞之情形時,重組載體較佳為於原核生物中能夠自主複製,並且含有啟動子、核糖體結合序列、包含編碼CD40或FAP之部分之DNA、及轉錄終止序列的載體。又,對於該重組載體,轉錄終止序列並非必需,但較佳為於結構基因之正下方配置轉錄終止序列。進而,該重組載體亦可含有控制啟動子之基因。
作為該重組載體,較佳為使用將作為核糖體結合序列之夏因-達爾加諾序列(Shine-Dalgarno sequence)與起始密碼子之間調節為適當距離(例如6~18個鹼基)的質體。
又,作為編碼該CD40或FAP之DNA之鹼基序列,可以成為最適合於宿主內表現之密碼子之方式進行鹼基置換,藉此,可提高目標之CD40或FAP之生產率。
作為表現載體,只要為能夠於所使用之宿主細胞中發揮功能者,則可使用任一種,例如可列舉:pBTrp2、pBTac1、pBTac2(以上為Roche-Diagnostics公司製造)、pKK233-2(Pharmacia公司製造)、pSE280(Invitrogen公司製造)、pGEMEX-1(Promega公司製造)、pQE-8(Qiagen公司製造)、pKYP10(日本專利特開昭58-110600號公報)、pKYP200[Agricultural Biological Chemistry, 48, 669 (1984)]、pLSA1[Agric. Biol. Chem., 53, 277 (1989)]、pGEL1[Proc. Natl. Acad. Sci. USA, 82, 4306 (1985)]、pBluescript II SK(-)(Stratagene公司製造)、pTrs30[由大腸桿菌JM109/pTrS30(FERM BP-5407)製備]、pTrs32[由大腸桿菌JM109/pTrS32(FERM BP-5408)製備]、pGHA2[由大腸桿菌IGHA2(FERM BP-400)製造,日本專利特開昭60-221091號公報]、pGKA2[由大腸桿菌IGKA2(FERM BP-6798)製備,日本專利特開昭60-221091號公報]、pTerm2(美國專利第4,686,191號說明書,美國專利第4,939,094號說明書,美國專利第5,160,735號說明書)、pSupex、pUB110、pTP5、pC194、pEG400[J. Bacteriol., 172, 2392 (1990)]、pGEX(Pharmacia公司製造)、pET系統(Novagen公司製造)或pME18SFL3(東洋紡織公司製造)等。
作為啟動子,只要為於所使用之宿主細胞中發揮功能者,則可為任何者。例如可列舉:trp啟動子(Ptrp)、lac啟動子、PL啟動子、PR啟動子或T7啟動子等源自大腸桿菌或噬菌體等之啟動子。又,例如亦可列舉:使2個Ptrp串聯而成之串聯啟動子、tac啟動子、lacT7啟動子或let I啟動子等經人工設計改變之啟動子等。
作為宿主細胞,例如可列舉:大腸桿菌XL1-Blue、大腸桿菌XL2-Blue、大腸桿菌DH1、大腸桿菌MC1000、大腸桿菌KY3276、大腸桿菌W1485、大腸桿菌JM109、大腸桿菌HB101、大腸桿菌No.49、大腸桿菌W3110、大腸桿菌NY49、或大腸桿菌DH5α等。
作為向宿主細胞導入重組載體之方法,只要為向所使用之宿主細胞導入DNA之方法,則可使用任一種方法,例如可列舉:使用鈣離子之方法[Proc. Natl. Acad. Sci. USA, 69, 2110 (1972)、Gene, 17, 107 (1982)、Molecular & General Genetics, 168, 111 (1979)]。
於使用動物細胞作為宿主之情形時,作為表現載體,只要為於動物細胞中發揮功能者,則可使用任一種,例如可列舉:pcDNAI(Invitrogen公司製造)、pcDM8(舟越公司製造)、pAGE107[日本專利特開平3-22979號公報;Cytotechnology, 3, 133 (1990)]、pAS3-3(日本專利特開平2-227075號公報)、pCDM8[Nature, 329, 840 (1987)]、pcDNAI/Amp(Invitrogen公司製造)、pcDNA3.1(Invitrogen公司製造)、pREP4(Invitrogen公司製造)、pAGE103[J. Biochemistry, 101, 1307 (1987)]、pAGE210、pME18SFL3或pKANTEX93(國際公開第97/10354號)等。
作為啟動子,只要為於動物細胞中能夠發揮功能者,則可使用任一種,例如可列舉:巨細胞病毒(CMV)之即刻早期(IE)基因之啟動子、SV40之初期啟動子、反轉錄病毒之啟動子、金屬硫蛋白啟動子、熱休克啟動子、SRα啟動子或來源於小鼠白血病病毒(Molony Murine Leukemia Virus)之啟動子或促進子。又,亦可將人類CMV之IE基因之促進子與啟動子一併使用。
作為宿主細胞,例如可列舉:人類伯奇氏淋巴瘤細胞Namalwa、源自非洲綠猴腎臟之細胞COS、源自中國倉鼠卵巢之細胞CHO、或人類白血病細胞HBT5637(日本專利特開昭63-000299號公報)等。
作為向宿主細胞導入重組載體之方法,只要為向動物細胞導入DNA之方法,則可使用任一種方法,例如可列舉:電穿孔法[Cytotechnology, 3, 133 (1990)]、磷酸鈣法(日本專利特開平2-227075號公報)、或脂轉染法[Proc. Natl. Acad. Sci. USA, 84, 7413(1987)]等。
將源自保有組入有以上述方式獲得之編碼CD40或FAP之DNA之重組載體的微生物或動物細胞等的轉形體於培養基中進行培養,使該CD40及FAP之至少一者於培養物中生成蓄積,自該培養物中進行採集,藉此可製造CD40或FAP。將該轉形體於培養基中進行培養之方法可依據宿主之培養所使用之通常方法進行。
於利用源自真核生物之細胞來表現之情形時,可獲得附加有糖或糖鏈之CD40或FAP。
於培養經使用誘導性之啟動子之重組載體轉形之微生物時,亦可視需要將誘導劑(inducer)添加至培養基中。例如,於培養經使用lac啟動子之重組載體轉形之微生物之情形時,亦可將異丙基-β-D-硫代半乳糖苷等添加至培養基中,於培養經使用trp啟動子之重組載體轉形之微生物之情形時,亦可將吲哚丙烯酸等添加至培養基中。
作為培養將動物細胞作為宿主所獲得之轉形體之培養基,例如可列舉:通常所使用之RPMI1640培養基[The Journal of the American Medical Association, 199, 519 (1967)]、Eagle之MEM培養基[Science, 122, 501 (1952)]、杜爾貝科改良MEM培養基[Virology, 8, 396(1959)]、199培養基[Proc. Soc. Exp. Biol. Med., 73, 1 (1950)]、Iscove's Modified Dulbecco's Medium(IMDM)培養基、或於該等培養基中添加有胎牛血清(FBS)等之培養基等。培養係通常於pH值6~8、30~40℃、5%CO2
存在下等條件下進行1~7天。又,於培養中亦可視需要,將康黴素或青黴素等抗生素添加至培養基中。
作為編碼CD40或FAP之基因之表現方法,除直接表現以外,亦可使用分泌生產或融合蛋白質表現等方法[Molecular Cloning, A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press (1989)]。作為CD40或FAP之生產方法,例如可列舉:於宿主細胞內生產CD40或FAP之方法;使CD40或FAP分泌至宿主細胞外之方法;或於宿主細胞外膜上生產CD40或FAP之方法,且可藉由改變所使用之宿主細胞或所生產之CD40或FAP之結構而選擇適當之方法。
例如製作於編碼細胞外區之胺基酸序列之DNA上連結編碼抗體之Fc區之DNA、編碼麩胱甘肽S-轉移酶(GST)之DNA、或編碼FLAG標籤之DNA或者編碼組胺酸標籤之DNA等而成的DNA,使之表現並進行純化,藉此可製作抗原融合蛋白質。具體而言,例如可列舉:使CD40或FAP之細胞外區與人類IgG之Fc區結合而成之Fc融合蛋白質、CD40或FAP之細胞外區與麩胱甘肽S-轉移酶(GST)之融合蛋白質。
於在宿主細胞內或宿主細胞外膜上產生CD40或FAP之情形時,可藉由使用Paulson等人之方法[J. Biol. Chem., 264, 17619 (1989)]、LOW等人之方法[Proc. Natl. Acad. Sci., USA, 86, 8227 (1989)、Genes Develop., 4, 1288 (1990)]、日本專利特開平05-336963號公報或國際公開第94/23021號等所記載之方法,使CD40或FAP積極地分泌至宿主細胞外。又,亦可利用使用二氫葉酸還原酶基因等之基因擴增系統(日本專利特開平2-227075號公報)而使CD40或FAP之產生量上升。
所產生之CD40或FAP例如可以下述方式進行單離、純化。
於CD40或FAP以溶解狀態在細胞內表現之情形時,於培養結束後藉由離心分離而回收細胞,於水系緩衝液中懸浮後,使用超音波破碎機、法式壓碎機、Manton-Gaulin均質機或Dyno磨機等將細胞破碎,而獲得無細胞萃取液。可藉由將該無細胞萃取液進行離心分離而獲得上清液,將通常之蛋白質之單離純化法、即溶劑萃取法、利用硫酸銨等之鹽析法、除鹽法、利用有機溶劑之沈澱法、使用二乙胺基乙基(DEAE)-瓊脂糖凝膠、DIAION HPA-75(三菱化學公司製造)等樹脂之陰離子交換層析法、使用S-瓊脂糖凝膠(S-Sepharose)FF(Pharmacia公司製造)等樹脂之陽離子交換層析法、使用丁基瓊脂糖凝膠(Butyl Sepharose)、苯基瓊脂糖凝膠(Phenyl Sepharose)等樹脂之疏水性層析法、使用分子篩之凝膠過濾法、親和層析法、層析聚焦(Chromatofocusing)法、或等電點電泳等電泳法等方法單獨或組合使用,而自所獲得之上清液獲得純化蛋白質。
於CD40或FAP在細胞內形成不溶體而表現之情形時,與上述同樣地回收細胞後將之破碎,進行離心分離,藉此以沈澱組分回收該CD40或FAP之不溶體。利用蛋白質改性劑將所回收之該CD40或FAP之不溶體進行可溶化。可藉由對該可溶化液進行稀釋或透析而使該CD40或FAP恢復到正常之立體結構後,利用與上述相同之單離純化法而獲得多肽之純化蛋白質。
於CD40或FAP、或者其糖修飾體等衍生物被分泌至細胞外之情形時,可於培養上清液中回收該CD40或FAP、或者其糖修飾體等衍生物。可藉由與上述同樣地使用離心分離等方法對該培養上清液進行處理而取得可溶性組分,藉由使用與上述相同之單離純化法而自可溶性組分獲得純化蛋白質。
又,本發明中所使用之CD40或FAP亦可藉由Fmoc法或tBoc法等化學合成法進行製造。具體而言,例如可利用Advanced ChemTech公司製造、PerkinElmer公司製造、Pharmacia公司製造、Protein Technology Instrument公司製造、Synthecell-Vega公司製造、Perceptive公司製造或島津製作所公司製造等之肽合成機進行化學合成。
(2)抗體產生細胞之製備步驟
用(1)中所獲得之抗原來免疫小鼠、大鼠、倉鼠、兔、牛、或羊駝等動物,採取該動物之脾臟、淋巴結或末梢血中之抗體產生細胞。又,作為動物,例如可列舉富塚等人之文獻[Tomizuka. et al., Proc Natl Acad Sci USA., 97, 722, (2000)]中所記載之產生源自人類之抗體的基因轉殖小鼠作為被免疫動物,且為了提高免疫原性,可列舉CD40或FAP之調節劑剔除小鼠等作為被免疫動物。
免疫係藉由將抗原與弗氏完全佐劑、或氫氧化鋁凝膠與百日咳菌疫苗等適當之佐劑一併投予來進行。小鼠免疫時之免疫原投予法可為皮下注射、腹腔內注射、靜脈內注射、皮內注射、肌內注射或足底注射等之任一種,較佳為腹腔內注射、足底注射或靜脈內注射。於抗原為部分肽之情形時,與牛血清白蛋白(BSA)或匙孔螺血氰蛋白(Keyhole Limpet hemocyanin,KLH)等載體蛋白製作偶聯物,將其用作免疫原。
抗原之投予係於第1次之投予後,每隔1~2週進行5~10次。於各投予後第3~7天自眼底靜脈叢進行採血,使用酵素免疫測定法[Antibodies-A Laboratory Manual, Cold Spring Harbor Laboratory (1988)]等,對其血清之抗體效價進行測定。若將對於用於免疫之抗原,上述血清顯示出充分之抗體效價之動物作為融合用抗體產生細胞之供給源,則可提高以後之操作之效果。
於抗原之最終投予後第3~7天,自經免疫之動物摘除脾臟等包含抗體產生細胞之組織,採集抗體產生細胞。抗體產生細胞係漿細胞及作為其前驅細胞之淋巴細胞,其可自個體之任意部位獲得,一般而言,可自脾臟、淋巴結、骨髄、扁桃體、末梢血、或適當組合有該等者等中獲得,最常使用脾臟細胞。於使用脾臟細胞之情形時,將脾臟切碎、解開後,進行離心分離,進而去除紅血球而取得融合用抗體產生細胞。
(3)骨髓瘤之製備步驟
作為骨髓瘤,可使用源自小鼠、大鼠、豚鼠、倉鼠、兔或人類等哺乳動物之無自我抗體產生能力之細胞,一般而言,使用自小鼠獲得之培養細胞株、例如使用8-氮鳥嘌呤耐性小鼠(源自BALB/c)骨髓瘤細胞株P3-X63Ag8-U1(P3-U1)[Current Topics in Microbiology and Immunology, 18, 1 (1978)]、P3-NS1/1-Ag41(NS-1)[European J. Immunology, 6, 511 (1976)]、SP2/0-Ag14(SP-2)[Nature, 276, 269 (1978)]、P3-X63-Ag8653 (653)[J. Immunology, 123, 1548 (1979)]、或P3-X63-Ag8(X63)[Nature, 256, 495 (1975)]等。該細胞株係於適當之培養基、例如8-氮鳥嘌呤培養基[添加有麩醯胺、2-巰基乙醇、慶大黴素、FCS及8-氮鳥嘌呤之RPMI-1640培養基]、Iscove改良達爾伯克培養基(Iscove's Modified Dulbecco's Medium;以下稱為「IMDM」)、或達爾伯克改良伊格爾培養基(Dulbecco's Modified Eagle Medium;以下稱為「DMEM」)等培養基中繼代培養。於細胞融合之3~4天前將上述細胞株於正常培養基(例如,包含10% FCS之DMEM培養基)中進行繼代培養,確保融合當天2×107
個以上之細胞數。
(4)細胞融合
利用最低基礎培養基(Minimum Essential Medium,MEM)或磷酸緩衝生理鹽水(PBS;磷酸二鈉1.83 g、磷酸一鉀0.21 g、食鹽7.65 g、蒸餾水1升、pH值7.2)將(2)中所獲得之融合用抗體產生細胞與(3)中所獲得之骨髓瘤細胞充分洗淨,並以成為融合用抗體產生細胞:骨髓瘤細胞=5:1~10:1之方式加以混合,進行離心分離後,將上清液去除。將沈澱之細胞群充分地解開後,一面加入聚乙二醇-1000(PEG-1000)、MEM培養基及二甲基亞碸之混液一面於37℃下進行攪拌。進而每1~2分鐘添加MEM培養基1~2 mL數次後,添加MEM培養基以使總量成為50 mL。遠離心分離後,將上清液去除。將沈澱之細胞群緩慢地解開後,於HAT培養基[添加有次黃嘌呤、胸苷、及胺基喋呤之正常培養基]中緩慢地使細胞懸浮。將該懸浮液於5%CO2
培養箱中以37℃培養7~14天。
又,亦可藉由以下之方法進行細胞融合。將脾臟細胞與骨髓瘤細胞利用無血清培養基(例如DMEM)、或磷酸緩衝生理鹽水(以下稱為「磷酸緩衝液」)充分地洗淨,並以脾臟細胞與骨髓瘤細胞之細胞數之比成為5:1~10:1左右的方式加以混合,進行離心分離。將上清液去除,將沈澱之細胞群充分地解開後,一面滴加1 mL之包含50%(w/v) 聚乙二醇(分子量1000~4000)之無血清培養基一面進行攪拌。其後,緩慢地加入10 L之無血清培養基後,進行離心分離。再次捨棄上清液,使沈澱之細胞於包含適量之次黃嘌呤-胺基喋呤-胸苷(HAT)液及人類介白素-2(IL-2)之正常培養基(以下,稱為HAT培養基)中懸浮,分注至培養用培養板(以下,稱為培養板)之各孔中,於5%二氧化碳存在下,於37℃下培養2週左右。途中適當補充HAT培養基。
(5)融合瘤群之選擇
於用於融合之骨髓瘤細胞為8-氮鳥嘌呤耐性株之情形時、即為次黃嘌呤-鳥嘌呤-磷酸核糖轉移酶(HGPRT)缺損株之情形時,未融合之骨髓瘤細胞及骨髓瘤細胞彼此之融合細胞於HAT培養基中無法生存。另一方面,抗體產生細胞彼此之融合細胞及抗體產生細胞與骨髓瘤細胞之融合瘤可於HAT培養基中生存,但抗體產生細胞彼此之融合細胞不久會達到壽命。因此,藉由於HAT培養基中繼續培養,而僅抗體產生細胞與骨髓瘤細胞之融合瘤存活,結果可取得融合瘤。
針對生長成群落狀之融合瘤,將培養基更換為自HAT培養基中去除胺基喋呤後之培養基(以下,稱為HT培養基)。其後,可採集培養上清液之一部分,並使用下述之抗體效價測定法來選擇產生抗體之融合瘤。作為抗體效價之測定方法,例如可列舉:放射性同位素免疫測定法(RIA法)、固相酵素免疫測定法(ELISA法)、螢光抗體法及被動血球凝集反應法等各種之公知技術,就檢測感度、迅速性、準確性及操作之自動化之可能性等觀點而言,較佳為RIA法或ELISA法。
將藉由測定抗體效價而判明了產生所需抗體之融合瘤移至另一培養板來進行選殖。作為該選殖法,例如可列舉:以於培養板之1個孔中包含1個細胞之方式進行稀釋並進行培養之限制稀釋法;於軟瓊脂培養基中進行培養並回收群落之軟瓊脂法;藉由顯微操作儀單離1個細胞之方法;藉由細胞分選儀單離1個細胞之方法等。
關於確認到抗體效價之孔,例如選擇將利用限制稀釋法之選殖反覆進行2~4次而穩定地確認到抗體效價者作為產生對於CD40或FAP之單株抗體之融合瘤株。
(6)單株抗體之製備
對於經姥鮫烷處理[將2,6,10,14-四甲基十五烷(Pristane)0.5 mL進行腹腔內投予,飼養2週]之8~10週齡之小鼠或裸小鼠,將(5)中所獲得之單株抗體產生融合瘤注射至腹腔內。於10~21天融合瘤發生腹水癌化。自該小鼠採集腹水,進行離心分離,將固形物成分去除後,利用40~50%硫酸銨進行鹽析,利用辛酸沈澱法、DEAE-瓊脂糖凝膠管柱、蛋白A-管柱或凝膠過濾管柱進行純化,收集IgG或IgM組分,作為純化單株抗體。又,藉由於同一系統之小鼠(例如,BALB/c)或Nu/Nu小鼠、大鼠、豚鼠、倉鼠或兔等之腹腔內使該融合瘤增生,可獲得包含大量與CD40或FAP結合之單株抗體之腹水。
將(5)中所獲得之單株抗體產生融合瘤於添加有10%FBS之RPMI1640培養基等中進行培養後,藉由離心分離去除上清液,於GIT培養基、或添加有5%Daigo GF21之Hybridoma-SFM培養基等中懸浮,藉由燒瓶培養、旋轉培養或袋裝培養等來培養3~7天。將所獲得之細胞懸浮液進行離心分離,自所獲得之上清液藉由蛋白A管柱或蛋白G管柱進行純化,收集IgG組分,亦可獲得純化單株抗體。作為純化之簡便方法,亦可利用市售之單株抗體純化套組(例如,MAbTrap GII套組;Amersham Pharmacia Biotech公司製造)等。
抗體之亞型之確定係使用亞型分型套組(subtype typing kit),藉由酵素免疫測定法進行。蛋白量之測定係可藉由勞立法(Lowry method)及自280 nm下之吸光度[1.4(OD280
)=免疫球蛋白1 mg/mL]算出之方法來進行。
(7)單株抗體對於CD40或FAP之結合分析
單株抗體對於CD40或FAP之結合活性可藉由歐氏(雙向)擴散法(Ouchterlony)法、ELISA法、RIA法、流式細胞分析法(FCM)或表面電漿子共振法(SPR)等結合分析系統來進行測定。
歐氏(雙向)擴散法雖簡便,但於抗體濃度較低之情形時需要濃縮操作。另一方面,於使用ELISA法或RIA法之情形時,使培養上清液直接與抗原吸附固相進行反應,進而使用各種免疫球蛋白同型、與亞型對應之抗體作為二次抗體,藉此能夠一面鑑定抗體之同型、亞型,一面測定抗體之結合活性。
作為程序之具體例,使經純化或部分純化之重組CD40或FAP吸附於ELISA用96孔培養板等之固相表面,進而藉由與抗原無關係之蛋白質、例如牛血清白蛋白(BSA)對未吸附抗原之固相表面進行阻斷。利用包含磷酸緩衝生理鹽水(PBS)及0.05% Tween20之PBS(Tween-PBS)等將ELISA培養板洗淨後,使經階段稀釋之第1抗體(例如小鼠血清、培養上清液等)進行反應,使抗體與固定在培養板之抗原結合。繼而,分注經作為第2抗體之生物素、酵素(辣根過氧化酶;HRP、鹼性磷酸酶;ALP等)、化學發光物質或放射線化合物等標記之抗免疫球蛋白抗體,使結合至培養板之第1抗體與第2抗體反應。利用Tween-PBS充分洗淨後,進行與第2抗體之標記物質相應之反應,選擇與靶抗原特異性地反應之單株抗體。
FCM法中,可測定抗體對於抗原表現細胞之結合活性[Cancer Immunol. Immunother., 36, 373 (1993)]。抗體與在細胞膜上表現之膜蛋白質抗原結合意指該抗體識別天然存在之抗原之立體結構並進行結合。
作為SPR法,可列舉利用表面電漿子共振(Biacore)之動力學解析。例如使用Biacore T100,測定抗原與受驗物之間之結合之動力學,並利用機器附帶之解析軟體對其結果進行解析。作為程序之具體例,藉由胺偶合法將抗小鼠IgG抗體固定至感測器晶片CM5後,使融合瘤培養上清液或純化單株抗體等受驗物質流過以結合適當量,進而使濃度既知之複數種濃度之抗原流過,測定結合及解離。繼而,對於所獲得之資料,使用機器附帶之軟體,進行基於1:1結合模型之動力學解析,取得各種參數。或者,藉由例如胺偶合法將CD40或FAP固定至感測器晶片上後,使濃度既知之複數種濃度之純化單株抗體流過,對結合及解離進行測定。對於所獲得之資料,使用機器附帶之軟體,進行基於二價結合(bivalent binding)模型之動力學解析,取得各種參數。
又,於本發明中,與對於CD40或FAP之抗體競爭性地與CD40或FAP結合之抗體可藉由使受驗抗體共存於上述結合分析系統中並進行反應而選擇。即,對添加有受驗抗體時與抗原之結合會受到抑制之抗體進行篩選,藉此可獲得與上述中所取得之抗體競爭性地與CD40或FAP結合之抗體。
(8)對於CD40或FAP之單株抗體之表位之鑑定
本發明中,抗體所識別並結合之表位之鑑定可以如下方式進行。
例如,製作抗原之部分缺損體、改變了種間不同之胺基酸殘基之變異體、或改變了特定區域之變異體,若抗體對於該缺損體或變異體之反應性降低,則明確缺損部位或胺基酸改變部位為該抗體之表位。此種抗原之部分缺損體及變異體可使用適當之宿主細胞、例如大腸桿菌、酵母、植物細胞或哺乳動物細胞等,以分泌蛋白質之形式取得,亦可於宿主細胞之細胞膜上表現而以抗原表現細胞之形式製備。於膜型抗原之情形時,為了在保持抗原之立體結構之狀態下進行表現,較佳為於宿主細胞之膜上進行表現。又,亦可製作模仿抗原之一次結構或立體結構之合成肽,而確認抗體之反應性。關於合成肽,可列舉使用公知之肽合成技術而製作分子之各種部分肽之方法等。
例如,對於人類及小鼠之CD40或FAP之細胞外區,製作適當組合構成各區域之區而成之嵌合蛋白質,確定抗體對於該蛋白質之反應性,藉此可鑑定抗體之表位。其後,進一步細緻地,使用業者所周知之寡肽合成技術,合成各種其對應部分之寡肽、或該肽之變異體等,確定抗體對於該肽之反應性,藉此可特定表位。作為用以獲得多種寡肽之簡便方法,亦可利用市售之套組[例如,SPOTs套組(GENOSYS BIOTECHNOLOGIES公司製造)、使用多中心合成法之一系列多中心-肽合成套組(Chiron公司製造)等]。
與和與CD40或FAP結合之抗體所結合之表位相同之表位結合的抗體可藉由鑑定上述結合分析系統中所獲得之抗體之表位,製作該表位之部分合成肽、擬態為該表位之立體結構之合成肽、或該表位之重組體等並進行免疫而取得。
例如若表位為膜蛋白質,則可藉由使全細胞外區或一部分細胞外域與適當之標籤、例如FLAG標籤、組胺酸標籤、GST蛋白質或抗體Fc區域等連結而製作重組融合蛋白質,用該重組蛋白質進行免疫而更有效率地製作該表位特異性抗體。
2.基因重組抗體之製作
作為基因重組抗體之製作例,雖於P. J. Delves., ANTIBODY PRODUCTION ESSENTIAL TECHNIQUES., 1997 WILEY、P. Shepherd and C. Dean. Monoclonal Antibodies., 2000 OXFORD UNIVERSITY PRESS及J. W. Goding., Monoclonal Antibodies : principles and practice., 1993 ACADEMIC PRESS等有概論,但於以下表示嵌合抗體、人類化抗體及人類抗體之製作方法。又,關於基因重組小鼠、大鼠、倉鼠及兔子抗體,亦可藉由相同之方法進行製作。
(1)自融合瘤取得編碼單株抗體之V區之cDNA
編碼單株抗體之VH及VL之cDNA之取得例如可以如下方式進行。
首先,自產生單株抗體之融合瘤提取mRNA而合成cDNA。繼而,將所合成之cDNA選殖至噬菌體或質體等載體而製作cDNA庫。使用編碼抗體之C區部分或V區部分之DNA作為探針,自該庫分別單離具有編碼VH或VL之cDNA之重組噬菌體或重組質體。確定所單離之重組噬菌體或重組質體內之VH或VL之完整鹼基序列,根據該鹼基序列推測VH或VL之完整胺基酸序列。
作為製作融合瘤時所使用之非人類動物,使用小鼠、大鼠、倉鼠或兔等,但只要能夠製作融合瘤,則可使用任何動物。
於自融合瘤製備總RNA時,使用硫氰酸胍-三氟乙酸銫法[Methods in Enzymol., 154, 3 (1987)]、或RNA簡易套組(Qiagen公司製造)等套組等。
於自總RNA製備mRNA時,使用寡(dT)固定化纖維素管柱法[Molecular Cloning, A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press (1989)]、或Oligo-dT30<Super>mRNA Purification Kit(TAKARA BIO公司製造)等套組等。又,亦可使用Fast Track mRNA Isolation Kit(Invitrogen公司製造)、或QuickPrep mRNA Purification Kit(Pharmacia公司製造)等套組,而製備mRNA。
於cDNA之合成及cDNA庫之製作時,使用公知之方法[Molecular Cloning, A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press(1989)、Current Protocols in Molecular Biology, Supplement 1, John Wiley & Sons (1987-1997)]或SuperScript Plasmid System for cDNA Synthesis and Plasmid Cloning(Invitrogen公司製造)或ZAP-cDNA Synthesis Kit(Stratagene公司製造)等套組等。
於製作cDNA庫時,作為供組入以自融合瘤提取之mRNA為模板所合成之cDNA的載體,只要為供組入該cDNA之載體,則可使用任何者。
例如使用ZAP Express[Strategies, 5, 58(1992)]、pBluescript II SK(+)[Nucleic Acids Research, 17, 9494 (1989)]、λZAPII(Stratagene公司製造)、λgt10、λgt11[DNA Cloning: A Practical Approach, I, 49 (1985)]、Lambda BlueMid(Clontech公司製造)、λExCell、pT7T3-18U(Pharmacia公司製造)、pcD2[Mol. Cell. Biol., 3, 280 (1983)]或pUC18[Gene, 33, 103 (1985)]等。
關於供導入由噬菌體或質體載體構建之cDNA庫之大腸桿菌,只要為可導入、表現及維持該cDNA庫者,則可使用任何者。例如使用XL1-Blue MRF'[Strategies, 5, 81 (1992)]、C600[Genetics, 39, 440 (1954)]、Y1088、Y1090[Science, 222, 778 (1983)]、NM522[J. Mol. Biol., 166, 1 (1983)]、K802[J. Mol. Biol., 16, 118 (1966)]、或JM105[Gene, 38, 275 (1985)]等。
自cDNA庫選擇編碼非人類抗體之VH或VL之cDNA純系時,使用利用同位素或經螢光標記之探針之菌落雜交法、或者噬菌斑雜交法[Molecular Cloning, A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press(1989)]等。
又,製備引子,以自mRNA合成之cDNA或cDNA庫為模板而進行聚合酶鏈反應法[以下,記載為PCR法,Molecular Cloning, A Laboratory Manual, Second Edition , Cold Spring Harbor Laboratory Press (1989)、Current Protocols in Molecular Biology, Supplement 1, John Wiley & Sons (1987-1997)],藉此亦可製備編碼VH或VL之cDNA。
利用適當之限制酶等切斷所選擇之cDNA後,選殖至pBluescript SK(-)(Stratagene公司製造)等質體,藉由通常所使用之鹼基序列分析方法等而確定該cDNA之鹼基序列。例如進行雙脫氧法[Proc. Natl. Acad. Sci. USA, 74, 5463 (1977)]等反應後,使用A.L.F.DNA核酸定序(Pharmacia公司製造)等鹼基序列自動分析裝置等來進行解析。
根據所確定之完整鹼基序列分別推測VH及VL之完整胺基酸序列,與既知之抗體之VH及VL之完整胺基酸序列[Sequences of Proteins of Immunological Interest, US Dept. Health and Human Services (1991)]進行比較,藉此確認所取得之cDNA是否編碼包括分泌訊息序列在內之抗體之VH及VL各自之完整胺基酸序列。
關於包含分泌訊息序列之抗體之VH及VL各自之完整胺基酸序列,與既知之抗體之VH及VL之完整胺基酸序列[Sequences of Proteins of Immunological Interest, US Dept. Health and Human Services (1991)]進行比較,藉此可推測分泌訊息序列之長度及N末端胺基酸序列,進而可鑑定該等所屬之亞群。
又,關於VH及VL之各CDR之胺基酸序列,可藉由與既知之抗體之VH及VL之胺基酸序列[Sequences of Proteins of Immunological Interest, US Dept. Health and Human Services (1991)]進行比較而推測。
又,對於所獲得之VH及VL之完整胺基酸序列,例如使用SWISS-PROT或PIR-Protein等任意之資料庫,進行基於BLAST法[J. Mol. Biol., 215, 403 (1990)]等之同源性檢索,藉此可確認該VH及VL之完整胺基酸序列是否為新穎者。
(2)基因重組抗體表現用載體之構建
基因重組抗體表現用載體可藉由將編碼人類抗體之CH及CL之至少一者之DNA選殖至動物細胞用表現載體而構建。
作為人類抗體之C區,可使用任意之人類抗體之CH及CL,例如可使用人類抗體之γ1亞型之CH及κ型之CL等。作為編碼人類抗體之CH及CL之DNA,使用cDNA,但亦可使用包含外顯子與內含子之染色體DNA。
關於動物細胞用表現載體,只要為可組入編碼人類抗體之C區之基因並表現者,則可使用任何者,例如可使用pAGE107[Cytotechnol., 3, 133 (1990)]、pAGE103[J. Biochem., 101, 1307 (1987)]、pHSG274[Gene, 27, 223 (1984)]、pKCR[Proc. Natl. Acad. Sci. USA, 78, 1527 (1981)]、pSG1bd2-4[Cytotechnol., 4, 173 (1990)]、或pSE1UK1Sed1-3[Cytotechnol., 13, 79 (1993)]、INPEP4(Biogen-IDEC公司製造)、N5KG1val(美國專利第6,001,358號說明書)、N5KG4PE R409K(國際公開第2006/033386號所記載)、N5KG2載體(國際公開第2003/033538號所記載)、轉位子載體(國際公開第2010/143698號)等。
作為動物細胞用表現載體之啟動子與促進子,可使用SV40之初期啟動子[J. Biochem., 101, 1307 (1987)]、來源於小鼠白血病病毒LTR[Biochem. Biophys. Res. Commun., 149, 960 (1987)]、CMV啟動子(美國專利第5,168,062號說明書)或免疫球蛋白H鏈之啟動子[Cell, 41, 479 (1985)]及促進子[Cell, 33, 717 (1983)]等。
為了表現基因重組抗體,就載體之構建容易性、向動物細胞之導入容易性、細胞內之抗體H鏈及L鏈之表現量之均衡性等觀點而言,使用搭載有抗體H鏈及L鏈之兩基因之載體(串聯型載體)[J. Immunol. Methods, 167, 271 (1994)],但亦可將分別搭載有抗體H鏈與L鏈之各基因之複數個載體(分離型載體)組合使用。
作為串聯型之基因重組抗體表現用載體,使用pKANTEX93(國際公開第97/10354號)、pEE18[Hybridoma, 17, 559 (1998)]、N5KG1val(美國專利第6,001,358號說明書)、N5KG4PE R409K(國際公開第2006/033386號所記載)、N5KG2載體(國際公開第2003/033538號所記載)、Tol2轉位子載體(國際公開第2010/143698號)等。
(3)嵌合抗體表現載體之構建
將編碼(1)中所獲得之非人類抗體之VH或VL之cDNA分別選殖至(2)中所獲得之基因重組抗體表現用載體內的編碼人類抗體之CH或CL之各基因的上游,藉此可構建嵌合抗體表現載體。
首先,為了將編碼非人類抗體之VH或VL之cDNA之3'末端側、與人類抗體之CH或CL之5'末端側連結,而製作以編碼連結部分之鹼基序列適當之胺基酸,且成為適當之限制酶識別序列之方式所設計的VH及VL之cDNA。繼而,將所製作之VH及VL之cDNA以其等以適當形式表現的方式分別選殖至(2)中所獲得之基因重組抗體表現用載體內的編碼人類抗體之CH或CL之各基因之上游,而構建嵌合抗體表現載體。
又,亦可使用於兩端具有適當限制酶之識別序列之合成DNA,藉由PCR法分別擴增編碼非人類抗體之VH或VL的cDNA,並選殖至(2)中所獲得之基因重組抗體表現用載體,藉此構建嵌合抗體表現載體。
(4)編碼人類化抗體之V區之cDNA之製作
編碼人類化抗體之VH或VL之cDNA可以如下方式進行製作。首先,分別選擇供移植(1)中所獲得之非人類抗體之VH或VL之CDR之胺基酸序列的人類抗體之VH或VL之架構區(以下,記載為FR)之胺基酸序列。
關於所選擇之FR之胺基酸序列,只要為源自人類抗體者,則可使用任一者。例如使用登陸於蛋白質資料庫(Protein Data Bank)等資料庫中之人類抗體之FR之胺基酸序列、或者人類抗體之FR之各亞群之共通胺基酸序列[Sequences of Proteins of Immunological Interest, US Dept. Health and Human Services (1991)]等。為了抑制抗體之結合活性之降低,而選擇與原本之非人類抗體之VH或VL之FR之胺基酸序列具有儘可能高之同源性(60%以上)的人類FR之胺基酸序列。
繼而,將原本之非人類抗體之CDR之胺基酸序列分別移植至所選擇的人類抗體之VH或VL之FR之胺基酸序列,而分別設計人類化抗體之VH或VL之胺基酸序列。考慮抗體基因之鹼基序列中可見之密碼子之使用頻度[Sequences of Proteins of Immunological Interest, US Dept. Health and Human Services(1991)],將所設計之胺基酸序列轉化為DNA序列,藉此分別設計人類化抗體之VH或VL之cDNA序列。
基於所設計之cDNA序列,合成包含100~150個鹼基左右長度之數條合成DNA,使用其等進行PCR。於該情形時,就PCR中之反應效率及能夠合成之DNA長度之觀點而言,較佳為對於H鏈及L鏈各設計4~6條合成DNA。又,亦可合成可變區全長之合成DNA來使用。
進而,將適當之限制酶之識別序列導入至位於兩端之合成DNA之5'末端,藉此可容易地將編碼人類化抗體之VH或VL之cDNA選殖至(2)中所獲得之基因重組抗體表現用載體。PCR後,將擴增產物分別選殖至pBluescript SK(-)(Stratagene公司製造)等質體,藉由與(1)中所記載之方法相同之方法來確定鹼基序列,取得具有所需之編碼人類化抗體之VH或VL之胺基酸序列之DNA序列的質體。
(5)人類化抗體之V區之胺基酸序列之改變
人類化抗體若僅將非人類抗體之VH及VL之CDR移植至人類抗體之VH及VL之FR,則其抗原結合活性較原本之非人類抗體降低[BIO/TECHNOLOGY, 9, 266 (1991)]。因此,對人類抗體之VH及VL之FR之胺基酸序列中直接參與與抗原之結合之胺基酸殘基、與CDR之胺基酸殘基相互作用之胺基酸殘基、及維持抗體之立體結構且間接地參與與抗原之結合之胺基酸殘基進行鑑定,將該等胺基酸殘基置換為原本之非人類抗體之胺基酸殘基,藉此可使已降低之人類化抗體之抗原結合活性上升。
為了鑑定與抗原結合活性相關之FR之胺基酸殘基,使用X射線結晶解析[J. Mol. Biol., 112, 535 (1977)]或電腦建模[Protein Engineering, 7, 1501 (1994)]等,藉此可進行抗體之立體結構之構建及解析。又,關於各抗體,製作數種改變體,反覆研究各自與抗原結合活性之相關性,並進行試誤,藉此可取得具有所需之抗原結合活性之改變人類化抗體。
人類抗體之VH及VL之FR之胺基酸殘基可藉由使用改變用合成DNA來進行(4)中所記載之PCR而進行改變。關於PCR後之擴增產物,藉由(1)中所記載之方法來確定鹼基序列以確認實施過目標改變。
(6)人類化抗體表現載體之構建
將所構建之編碼人類化抗體之VH或VL之cDNA分別選殖至(2)中所獲得之基因重組抗體表現用載體之編碼人類抗體之CH或CL的各基因之上游,而可構建人類化抗體表現載體。
例如,將適當之限制酶之識別序列導入至構建(4)及(5)中所獲得之人類化抗體之VH或VL時所使用之合成DNA中位於兩端之合成DNA的5'末端,藉此以其等以適當之形式進行表現之形式分別選殖至(2)中所獲得之基因重組抗體表現用載體內之編碼人類抗體之CH或CL的各基因之上游。
(7)人類抗體表現載體之構建
於使用產生人類抗體之動物作為被免疫動物,構建產生單株抗體之融合瘤的情形時,於(1)中可獲得人類抗體之VH及VL之胺基酸序列及cDNA序列。因此,將編碼(1)中所獲得之人類抗體之VH或VL之基因分別選殖至(2)中所獲得之基因重組抗體表現用載體之編碼人類抗體之CH或CL之各基因的上游,藉此可構建人類抗體表現載體。
(8)基因重組抗體之短暫性表現
可使用(3)、(6)及(7)中所獲得之基因重組抗體表現用載體、或改變其等而成之表現載體,使基因重組抗體短暫性表現,而有效率地評價所獲得之多種基因重組抗體之抗原結合活性。
關於供導入表現載體之宿主細胞,只要為可表現基因重組抗體之宿主細胞,則可使用任何細胞,例如使用COS-7細胞[American Type Culture Collection(ATCC)編號:CRL1651]。向COS-7細胞導入表現載體時,使用DEAE-葡聚糖法[Methods in Nucleic Acids Res., CRC press (1991)]、或脂轉染法[Proc. Natl. Acad. Sci. USA, 84, 7413 (1987)]等。
導入表現載體後,使用酵素免疫抗體法[Monoclonal Antibodies-Principles and practice, Third Edition, Academic Press (1996)、Antibodies - A Laboratory Manual, Cold Spring Harbor Laboratory (1988)、單純系抗體實驗指南、Kodansha Scientific (1987)]等,對培養上清液中之基因重組抗體之表現量及抗原結合活性進行測定。
(9)基因重組抗體之穩定表現株之取得與基因重組抗體之製備
將(3)、(6)及(7)中所獲得之基因重組抗體表現用載體導入至適當之宿主細胞,藉此可獲得穩定地表現基因重組抗體之轉形株。
關於表現載體向宿主細胞之導入,例如可列舉:電穿孔法[日本專利特開平2-257891號公報、Cytotechnology, 3, 133 (1990)]、鈣離子方法、電穿孔法、球形質體法、乙酸鋰法、磷酸鈣法、脂轉染法等。又,作為下述向動物導入基因之方法,例如可列舉:顯微注射法、使用電穿孔法或脂轉染法向ES細胞導入基因之方法、及核移植法等。
作為供導入基因重組抗體表現用載體之宿主細胞,只要為可表現基因重組抗體之宿主細胞,則可使用任何細胞。例如使用小鼠SP2/0-Ag14細胞(ATCC CRL1581)、小鼠P3X63-Ag8.653細胞(ATCC CRL1580)、中國倉鼠CHO-K1細胞(ATCC CCL-61)、DUKXB11(ATCC CCL-9096)、Pro-5細胞(ATCC CCL-1781)、CHO-S細胞(Life Technologies、Cat No. 11619)、缺失有二氫葉酸還原酶基因(dhfr)之CHO細胞(CHO/DG44細胞)[Proc. Natl. Acad. Sci. USA, 77, 4216 (1980)]、獲得了凝集素耐性之Lec13細胞[Somatic Cell and Molecular genetics, 12, 55 (1986)]、缺失有α1,6-海藻糖轉移酶基因之CHO細胞(國際公開第2005/035586號、國際公開第02/31140號)、大鼠YB2/3HL.P2.G11.16Ag.20細胞(ATCC編號:CRL1662)等。
又,亦可使用:與細胞內糖核苷酸GDP-海藻糖之合成相關之酶等蛋白質、與海藻糖之1位α結合至N-糖苷結合複合型糖鏈之還原末端之N-乙醯胺基葡萄糖之6位的糖鏈修飾相關的酶等蛋白質、或者與細胞內糖核苷酸GDP-海藻糖向高爾基體的輸送相關之蛋白質等之活性降低或缺失之宿主細胞、例如缺失有α1,6-海藻糖基轉移酶基因之CHO細胞(國際公開第2005/035586號、國際公開第02/31140號)等。
藉由於導入表現載體後,將穩定地表現基因重組抗體之轉形株於包含G418硫酸鹽(以下,記載為G418)等藥劑之動物細胞培養用培養基中進行培養而選擇(日本專利特開平2-257891號公報)。
關於動物細胞培養用培養基,使用RPMI1640培養基(Invitrogen公司製造)、GIT培養基(日本製藥公司製造)、EX-CELL301培養基(JRH公司製造)、EX-CELL302培養基(JRH公司製造)、EX-CELL325培養基(JRH公司製造)、IMDM培養基(Invitrogen公司製造)或融合瘤-SFM培養基(Invitrogen公司製造)、或者於該等培養基中添加有FBS等各種添加物之培養基等。將所獲得之轉形株於培養基中進行培養,藉此使基因重組抗體於培養上清液中表現並蓄積。培養上清液中之基因重組抗體之表現量及抗原結合活性可藉由ELISA法等進行測定。又,可利用DHFR擴增系統(日本專利特開平2-257891號公報)等,使轉形株所產生之基因重組抗體之表現量提昇。
基因重組抗體係使用蛋白A-管柱,自轉形株之培養上清液進行純化[Monoclonal Antibodies - Principles and practice, Third Edition, Academic Press (1996)、Antibodies - A Laboratory Manual, Cold Spring Harbor Laboratory (1988)]。又,亦可將凝膠過濾、離子交換層析法及超過濾等蛋白質之純化中所使用之方法組合來進行純化。
經純化之基因重組抗體之H鏈、L鏈或抗體分子整體之分子量可使用聚丙烯醯胺凝膠電泳法[Nature, 227, 680(1970)]、或西方墨點法[Monoclonal Antibodies - Principles and practice, Third Edition, Academic Press (1996)、Antibodies - A Laboratory Manual, Cold Spring Harbor Laboratory (1988)]等進行測定。
3.雙專一性抗體之設計
本發明之雙專一性抗體可藉由如下方式製作:分別設計第二抗原結合域與IgG部分,設計連結有其等之雙專一性抗體。
3-1.IgG部分之設計
IgG部分可藉由如下方式獲得:使用上述1.中所記載之方法取得單株抗體,使用上述2.中所記載之方法來確定各抗體之CDR及可變區之cDNA序列,設計包含抗體之CDR或可變區之IgG部分。
3-2.第二抗原結合域之設計
於第二抗原結合域包含抗體之CDR或可變區之情形時,可藉由如下方式進行製作:利用上述1.及2.中所記載之方法來確定抗體之CDR或可變區之DNA序列,設計包含其等之第二抗原結合域。作為此種第二抗原結合域,亦可使用如scFv等般使VH與VL直接或經由適當之連接子結合而成之單鏈者、如Fab及dsFv等般以於雙鏈中表現並於表現後進行雙硫鍵結之方式設計者、或者VHH等。又,作為第二抗原結合域,亦可使用抗原蛋白質之配位體、或者包含該配位體之與抗原蛋白質結合之部分之肽片段。第二抗原結合域之抗原結合活性係利用上述方法進行評價,可選擇保持有抗原結合活性者。
4.雙專一性抗體之製作
抗原結合域除上述1.中所記載之使用融合瘤之方法以外,亦可藉由噬菌體呈現(Phage Display)法、酵母呈現(Yeast display)法等技術進行單離並取得[Emmanuelle Laffy et al., Human Antibodies 14, 33-55, (2005)]。
又,於製作包含複數個VH與單一VL之雙專一性抗體之情形時,以雙專一性抗體中所包含之各抗原結合域分別特異性地與抗原進行反應的方式,使用噬菌體呈現法等進行篩選,選擇最適合單一VL之各VH。
具體而言,首先,使用上述1.中所記載之方法,用第1抗原免疫動物,自該動物之脾臟製作融合瘤,選殖編碼第1抗原結合域之DNA序列。繼而,用第2抗原免疫動物,自該動物之脾臟製備cDNA庫,自該庫,藉由PCR取得編碼VH之胺基酸序列之DNA。
繼而,製作表現連結第2抗原之免疫中所獲得之VH與第1抗原結合域之VL而成之scFv的噬菌體庫,使用該噬菌體庫進行篩選,藉此選擇呈現與第2抗原特異性地結合之scFv之噬菌體。根據所選擇之噬菌體,對編碼第2抗原結合域之VH之胺基酸序列之DNA序列進行選殖。
進而,於第二抗原結合域為Fab之情形時,設計編碼藉由重鏈恆定區將第1抗原結合域之VH與第2抗原結合域之VH連結而成之多肽之胺基酸序列的DNA序列,將編碼單一VL之胺基酸序列之DNA序列與該DNA序列一起插入至例如上述2.(2)中所記載之基因重組抗體表現用載體,藉此可構建本發明之雙專一性抗體之表現載體。
5.本發明之雙專一性抗體之活性評價
經純化之雙專一性抗體之活性評價可以如下方式進行。
本發明之雙專一性抗體對於表現CD40及FAP之至少一者之細胞株的結合活性可使用上述1.(7)記載之結合分析系統來進行測定。
對於表現CD40及FAP之至少一者之細胞之CDC活性、或ADCC活性可藉由公知之測定方法[Cancer Immunol. Immunother., 36, 373 (1993)]進行測定。
本發明之雙專一性抗體之細胞死亡誘導活性(亦稱為抗細胞活性)可藉由以下之方法進行測定。例如將細胞接種至96孔培養板中,添加抗體並培養一定期間後,使WST-8試劑(Dojindo公司製造)進行反應,藉由讀板儀測定450 nm之吸光度,藉此測定細胞之存活率。
自CD40或FAP向細胞內之訊息傳遞可藉由西方墨點法等檢測細胞內之蛋白質之磷酸化而進行評價。
6.使用本發明之雙專一性抗體之疾病之治療方法
本發明之雙專一性抗體可用於治療與CD40及FAP之至少一者相關之疾病、較佳為表現CD40或FAP之細胞所參與之疾病。作為與CD40及FAP之至少一者相關之疾病,例如可列舉:惡性腫瘤及癌症等。
作為惡性腫瘤及癌症,例如可列舉:大腸癌、結腸直腸癌、肺癌、乳癌、神經膠質瘤、惡性黑色素瘤(黑色素瘤)、甲狀腺癌、腎細胞癌、白血病、淋巴瘤、T細胞淋巴瘤、胃癌、胰腺癌、子宮頸癌、子宮內膜癌、卵巢癌、膽管癌、食道癌、肝臟癌、頭頸癌、鱗狀細胞癌、皮膚癌、尿路癌、膀胱癌、前列腺癌、絨毛膜癌、咽頭癌、喉頭癌、胸膜瘤、雄胚瘤、子宮內膜增生、子宮內膜症、胚組織瘤、纖維肉瘤、卡波西肉瘤、血管瘤、海綿狀血管瘤、血管母細胞瘤、視網膜胚細胞瘤、星狀細胞瘤、神經纖維瘤、少突膠質細胞瘤、神經管胚細胞瘤、神經胚細胞瘤、神經膠質瘤、橫紋肌肉瘤、神經膠母細胞瘤、骨原性肉瘤、平滑肌肉瘤及威爾姆斯瘤等。
含有本發明之雙專一性抗體或其衍生物之治療藥可為僅包含作為有效成分之該雙專一性抗體、或其衍生物者,較佳為通常與藥理學上所容許之1種以上之載體一起混合,而以藉由製劑學之技術領域中所公知之任意方法所製造之醫藥製劑的形式提供。
作為投予路徑,例如可列舉:經口投予、或者口腔內、呼吸道內、直腸內、皮下、肌內或靜脈內等非經口投予。作為投予形態,例如可列舉:噴霧劑、膠囊劑、錠劑、散劑、顆粒劑、糖漿劑、乳劑、栓劑、注射劑、軟膏或貼劑等。各種製劑可使用通常使用之賦形劑、增量劑、結合劑、浸潤劑、崩解劑、表面活性劑、潤滑劑、分散劑、緩衝劑、保存劑、增溶劑、防腐劑、著色料、香味劑、及穩定劑等,藉由常規方法進行製造。
作為賦形劑,例如可列舉:乳糖、果糖、葡萄糖、玉米澱粉、山梨糖醇、結晶纖維素、殺菌水、乙醇、甘油、生理鹽水及緩衝液等。作為崩解劑,例如可列舉:澱粉、海藻酸鈉、明膠、碳酸鈣、檸檬酸鈣、糊精、碳酸鎂及合成矽酸鎂等。
作為結合劑,例如可列舉:甲基纖維素或其鹽、乙基纖維素、阿拉伯膠、明膠、羥丙基纖維素及聚乙烯吡咯啶酮等。作為潤滑劑,例如可列舉:滑石、硬脂酸鎂、聚乙二醇及氫化植物油等。
作為穩定劑,例如可列舉:精胺酸、組胺酸、離胺酸、甲硫胺酸等胺基酸、人類血清白蛋白、明膠、葡聚糖40、甲基纖維素、亞硫酸鈉、偏亞硫酸鈉等。
作為其他添加劑,例如可列舉:糖漿、凡士林、甘油、乙醇、丙二醇、檸檬酸、氯化鈉、亞硝酸鈉及磷酸鈉等。
作為適合經口投予之製劑,例如可列舉:乳劑、糖漿劑、膠囊劑、錠劑、散劑或顆粒劑等。
如乳劑或糖漿劑之液體製備物係使用水、蔗糖、山梨糖醇或果糖等糖類;聚乙二醇或丙二醇等二醇類;芝麻油、橄欖油或大豆油等油類;對羥基苯甲酸酯類等防腐劑;或者草莓香料或胡椒薄荷等香料類等作為添加劑來進行製造。
膠囊劑、錠劑、散劑或顆粒劑等係使用乳糖、葡萄糖、蔗糖或甘露醇等賦形劑;澱粉或海藻酸鈉等崩解劑;硬脂酸鎂或滑石等潤滑劑;聚乙烯醇、羥丙基纖維素或明膠等結合劑;脂肪酸酯等界面活性劑或者甘油等塑化劑等作為添加劑來進行製造。
作為適合非經口投予之製劑,例如可列舉注射劑、栓劑或噴霧劑等。注射劑係使用包含鹽溶液、葡萄糖溶液、或者上述兩者之混合物之載體等來進行製造。
栓劑係使用可可脂、氫化脂肪或羧酸等載體來進行製造。噴霧劑係使用不會刺激接受者之口腔及呼吸道黏膜,且使本發明之單株抗體以微細粒子之形式分散而使吸收變得容易的載體等來進行製造。作為載體,例如可列舉乳糖或甘油等。又,亦可以霧劑或乾粉之形式進行製造。進而,於上述非經口劑中,亦可添加於適合經口投予之製劑中作為添加劑所例示之成分。
關於以本發明之雙專一性抗體之有效量與適當之稀釋劑及藥理學上可使用之載子的組合所投予之有效量,每次體重每1 kg為0.0001 mg~100 mg,且以2天至8週之間隔來投予。
7.使用本發明之雙專一性抗體之疾病之診斷方法
使用本發明之雙專一性抗體,對表現CD40及FAP之至少一者之細胞進行檢測或測定,藉此可診斷與CD40及FAP之至少一者相關之疾病、較佳為CD40或FAP之表現細胞所參與之疾病。
與CD40及FAP之至少一者相關之疾病即惡性腫瘤或癌症之診斷例如可以下述方式對CD40及FAP之至少一者進行檢測或測定來進行。
首先,對於自複數個健康者之生物體採集到之生物樣本,使用本發明之雙專一性抗體或者其等之衍生物,並使用下述免疫學方法,進行CD40及FAP之至少一者之檢測或測定,調查健康者之生物樣本中之CD40及FAP之至少一者之存在量。
繼而,於受驗者之生物樣本中,同樣地調查CD40及FAP之至少一者之存在量,將其存在量與健康者之存在量進行比較。於受驗者之CD40及FAP之至少一者之存在量較健康者增加之情形時,該受驗者被診斷為癌症。關於其他與CD40及FAP之至少一者相關之疾病之診斷,亦可藉由相同之方法進行診斷。
所謂免疫學方法,係使用施加有標記之抗原或抗體等,對抗體量或抗原量進行檢測或測定之方法。例如可列舉:放射性物質標記免疫抗體法、酵素免疫測定法、螢光免疫測定法、發光免疫測定法、西方墨點法或物理化學方法等。
作為放射性物質標記免疫抗體法,例如可列舉如下方法:使抗原或表現抗原之細胞等與本發明之雙專一性抗體進行反應,進而使施加過放射線標記之抗免疫球蛋白抗體或結合片段進行反應後,利用閃爍計數器等進行測定。
作為酵素免疫測定法,例如可列舉如下方法:使抗原或表現抗原之細胞等與本發明之雙專一性抗體進行反應,進而使施加過標記之抗免疫球蛋白抗體或結合片段進行反應後,利用吸光光度計對顯色色素進行測定。例如可列舉夾層ELISA法等。
作為酵素免疫測定法中所使用之標記體,可使用公知之酶標記[酵素免疫測定法,醫學書院(1987)]。例如使用鹼性磷酸酶標記、過氧化酶標記、螢光素酶標記、或生物素標記等。
夾層ELISA法係如下方法:使抗體與固相結合後,捕獲作為檢測或測定對象之抗原,使所捕獲之抗原與第2抗體進行反應。該ELISA法中,準備與欲檢測或測定之抗原結合之抗體或抗體片段且抗原結合部位不同之2種抗體,之後,預先使第1抗體或抗體片段吸附於培養板(例如96孔培養板),繼而預先利用FITC等螢光物質、過氧化酶等酵素、或者生物素等來標記第2抗體或抗體片段。與上述吸附有抗體之培養板中,使自體內分離出之細胞或其破碎液、組織或其破碎液、細胞培養上清液、血清、胸水、腹水、或眼液等反應後,使經標記之抗體或抗體片段反應,進行與標記物質對應之檢測反應。根據階段性地稀釋濃度已知之抗原所製作之校準曲線,而算出受驗樣品中之抗原濃度。
作為用於夾層ELISA法之抗體,可使用多株抗體或單株抗體中之任一種,亦可使用Fab、Fab'、或F(ab)2
等抗體片段。作為夾層ELISA法中所使用之2種抗體之組合,可為與不同表位結合之單株抗體或該抗體片段之組合,亦可為多株抗體與單株抗體或其抗體片段之組合。
作為螢光免疫測定法,例如利用文獻[Monoclonal Antibodies-Principles and practice, Third Edition, Academic Press (1996)、單純系抗體實驗指南、Kodansha Scientific (1987)]等中所記載之方法進行測定。作為螢光免疫測定法中所使用之標記體,可使用公知之螢光標記[螢光抗體法、Soft science公司(1983)]。例如使用FITC或RITC等。
作為發光免疫測定法,例如利用文獻[生物發光與化學發光 臨床檢查42、廣川書店(1998)]等所記載之方法進行測定。作為發光免疫測定法中所使用之標記體,可列舉公知之發光體標記,例如使用吖啶鎓酯或咯吩等。
作為西方墨點法,係如下方式進行測定:利用SDS(十二烷基硫酸鈉)-PAGE(聚丙烯醯胺凝膠)[Antibodies - A Laboratory Manual Cold Spring Harbor Laboratory (1988)]區分抗原或表現抗原之細胞等後,將該凝膠印漬至聚偏二氟乙烯(PVDF)膜或硝化纖維素膜,使該膜和與抗原結合之抗體或抗體片段進行反應,進而使施加有FITC等螢光物質、過氧化酶等酵素標記、或生物素標記等之抗IgG抗體或其抗體片段反應後,將該標記進行可視化。將一例示於以下。
首先,使表現具有所需胺基酸序列之多肽之細胞或組織溶解,於還原條件下,藉由SDS-PAGE法,使以每區帶之蛋白質量計0.1~30 μg進行泳動。繼而,將泳動之蛋白質轉移至PVDF膜,於室溫下與包含1~10%BSA之PBS(以下,記載為BSA-PBS)反應30分鐘而進行阻斷操作。此處,使本發明之雙專一性抗體進行反應,利用包含0.05~0.1%之Tween-20之PBS(Tween-PBS)洗淨,於室溫下,使經過氧化酶標記之山羊抗小鼠IgG反應2小時。利用Tween-PBS洗淨,使用ECL西方墨點法檢測試劑(Amersham公司製造)等,檢測該抗體所結合之帶,藉此檢測抗原。作為西方墨點法中之檢測所使用之抗體,可使用可與未保持天然型立體結構之多肽結合之抗體。
作為物理化學方法,例如藉由使作為抗原之CD40及FAP之至少一者與本發明之雙專一性抗體結合而形成凝集體,對該凝集體進行檢測。作為除此以外之物理化學方法,亦可使用毛細管法、單向免疫擴散法(single immunodiffusion)、免疫比濁法或乳膠免疫比濁法[臨床檢查法摘要,金原出版(1998)]等。
關於乳膠免疫比濁法,係使用使抗體或抗原致敏之粒徑0.1~1 μm左右之聚苯乙烯乳膠等載體,若藉由相對應之抗原或抗體而引起抗原抗體反應,則反應液中之散射光增加,而透過光減少。將該變化作為吸光度或積分球濁度進行檢測,藉此測定受驗樣品中之抗原濃度等。
另一方面,於檢測或測定表現CD40及FAP之至少一者之細胞時,可使用公知之免疫學檢測法,較佳為使用免疫沈澱法、免疫細胞染色法、免疫組織染色法或螢光抗體染色法等。
作為免疫沈澱法,係使表現CD40及FAP之至少一者之細胞等與本發明之雙專一性抗體反應後,添加對於蛋白G-瓊脂糖凝膠等免疫球蛋白具有特異性結合能力之載體,使抗原抗體複合體沈降。
或者,亦可藉由如以下之方法對表現CD40及FAP之至少一者之細胞進行檢測或測定。首先,將本發明之雙專一性抗體固相化至ELISA用96孔培養板後,藉由BSA-PBS進行阻斷。繼而,捨棄BSA-PBS,利用PBS充分洗淨後,使表現CD40及FAP之至少一者之細胞或組織之溶解液進行反應。利用SDS-PAGE用樣品緩衝液,自充分洗淨後之培養板提取免疫沈澱物,藉由上述西方墨點法進行檢測。
免疫細胞染色法或免疫組織染色法係如下方法:針對表現抗原之細胞或組織等,視情形為了使抗體之通過性變得良好而利用界面活性劑或甲醇等進行處理後,與本發明之雙專一性抗體進行反應,進而與施加有FITC等螢光標記、過氧化酶等酵素標記或生物素標記等之抗免疫球蛋白抗體或其結合片段進行反應,之後將該標記進行可視化,並利用顯微鏡進行觀察。又,可藉由使螢光標記之抗體與細胞進行反應並利用流式細胞儀進行解析之螢光抗體染色法[Monoclonal Antibodies-Principles and practice, Third edition, Academic Press (1996)、單純系抗體實驗指南、Kodansha Scientific (1987)]來進行檢測。尤其是本發明之雙專一性抗體可藉由螢光抗體染色法而檢測出於細胞膜上表現之CD40及FAP之至少一者。
又,螢光抗體染色法中,於使用FMAT8100HTS系統(Applied Biosystems公司製造)等之情形時,可不將形成之抗體-抗原複合體、與不參與形成抗體-抗原複合體之游離之抗體或抗原分離而測定抗原量或抗體量。
實施例
以下,藉由實施例對本發明具體地進行說明,但本發明並不限定於下述實施例。
[實施例1]可溶性人類及猴CD40抗原以及可溶性人類FAP抗原之製備
1.人類CD40及猴CD40之可溶性抗原之製備
藉由以下所記載之方法分別製作於C末端附加有FLAG-Fc之人類及猴CD40之細胞外域蛋白質。將編碼人類CD40細胞外域之鹼基序列示於序列編號1,將自該鹼基序列推測之胺基酸序列示於序列編號2,將編碼猴CD40細胞外域之鹼基序列示於序列編號3,將自該鹼基序列推測之胺基酸序列示於序列編號4。
(1)人類及猴CD40-FLAG-Fc載體之製作
基於人類CD40基因之鹼基序列(基因庫登錄號:NM_
001250、序列編號5;將該基因所編碼之胺基酸序列示於序列編號6),製作包含序列編號1所示之鹼基序列之人類CD40之細胞外域之基因片段。
利用限制酶KpnI及XbaI來消化包含FLAG-tag及人類IgG之Fc區之INPEP4載體(Biogen-IDEC公司製造),加上包含序列編號1所示之鹼基序列之第1~60號鹼基序列的人類CD40訊息序列編碼區,將所得之該細胞外域之基因片段插入至適當部位,製作人類CD40-FLAG-Fc表現載體。
藉由相同之方法,基於自猴末梢血液單核球(PBMC)選殖之猴CD40基因之鹼基序列(序列編號7;將該基因所編碼之胺基酸序列示於序列編號8),製作含有包含序列編號3所示之鹼基序列之猴CD40之細胞外域之基因片段的猴CD40-FLAG-Fc表現載體。
(2)人類及猴CD40-FLAG-Fc蛋白質之製作
使用FreeStyle(商標)293表現系統(Thermo Fisher公司製造),將1-(1)中所製作之人類CD40-FLAG-Fc表現載體導入至HEK293細胞並進行培養,利用短暫性表現系統使蛋白質表現。於載體導入5天後回收培養上清液,利用孔徑0.22 μm之膜濾器(MILLIPORE公司製造)進行過濾。
使用蛋白A(ProteinA)樹脂(MabSelect、GE healthcare公司製造)對該培養上清液進行親和純化。利用達爾伯克磷酸緩衝生理鹽水[D-PBS(-) 無Ca及Mg,液體;以下記載為D-PBS(-);Nacalai Tesque公司製造]將吸附於蛋白A樹脂之抗體洗淨,藉由20 mM之檸檬酸鈉、50 mM之NaCl緩衝液(pH值3.4)溶出,回收至包含1 M磷酸鈉緩衝液(pH值7.0)之試管中。
繼而,藉由使用VIVASPIN(Sartrius stealin公司製造)之超過濾,將緩衝液置換為D-PBS(-)後,利用孔徑0.22 μm之膜濾器Millex-Gv(Millipore公司製造)進行過濾殺菌,製作人類CD40-FLAG-Fc蛋白質。利用相同之方法,使用1-(1)中所製作之猴CD40-FLAG-Fc表現載體製作猴CD40-FLAG-Fc蛋白質。所取得之蛋白質之濃度係測定波長280 nm之吸光度,使用自各蛋白質之胺基酸序列推測之吸光係數而算出。
(3)人類及猴CD40-GST載體之製作
利用限制酶BglII及KpnI來消化包含GST區之N5載體(Biogen-IDEC公司製造),將包含1-(1)記載之序列編號1所表示之鹼基序列之人類CD40之細胞外域的基因片段插入至適當之部位,而製作人類CD40-GST表現載體。藉由相同之方法製作含有包含序列編號3所示之鹼基序列之細胞外域之基因片段的猴CD40-GST表現載體。
(4)人類及猴CD40-GST蛋白質之製作
將1-(3)中所製作之人類CD40-GST載體藉由與1.(2)相同之方法導入至HEK293細胞,培養該細胞後,將培養上清液利用膜濾器進行過濾。使該培養上清液與麩胱甘肽Sepharose 4B(GE healthcare Bioscience公司製造)進行反應,利用D-PBS(-)洗淨,使用含10 mM麩胱甘肽之50 mM三羥甲基胺基甲烷鹽酸鹽(pH值8.0)作為溶出緩衝液來進行親和純化。
對於所溶出之融合蛋白溶液,藉由與1-(2)相同之方法,利用超過濾及膜濾器進行過濾殺菌,而獲得人類CD40-GST蛋白質。又,使用猴CD40-GST載體,藉由相同之方法獲得猴CD40-GST蛋白質。所取得之蛋白質之濃度係測定波長280 nm之吸光度,使用自各蛋白質之胺基酸序列推測之吸光係數而算出。
2.人類及小鼠FAP之可溶性抗原之製備
藉由以下所記載之方法製作附加有FLAG-Fc或GST之人類及小鼠FAP之細胞外域蛋白質。將編碼人類FAP細胞外域之鹼基序列示於序列編號9,將自該鹼基序列推測之胺基酸序列示於序列編號10。將編碼小鼠FAP細胞外域之鹼基序列示於序列編號11,將自該鹼基序列推測之胺基酸序列示於序列編號12。
基於序列編號13所示之人類FAP基因(基因庫登錄號:NM_004460、將該基因所編碼之胺基酸序列示於序列編號14)及序列編號15所示之小鼠FAP基因(基因庫登錄號:NM_007986、將該基因所編碼之胺基酸序列示於序列編號16),以與1-(1)及(2)中所記載之方法相同之方式,分別獲得人類及小鼠FAP-FLAG-Fc蛋白質。
又,以與1-(3)及(4)中所記載之方法相同之方式,分別獲得人類及小鼠FAP-GST蛋白質。所取得之蛋白質之濃度係測定波長280 nm之吸光度,使用自各蛋白質之胺基酸序列推測之吸光係數而算出。
[實施例2]人類FAP表現HEK293細胞之製作
使用In-Fusion HD選殖套組(TAKARA BIO公司製造),將序列編號9所示之人類FAP之基因插入至pUC載體(TAKARA BIO公司製造),獲得目標之FAP表現載體Tn-pMug-Hygro-人類FAP。
使用FuGENE HD轉染試劑(Promega公司製造),將所取得之人類FAP表現載體Tn-pMug-Hygro-人類FAP導入至HEK293細胞。為了挑選表現載體導入細胞,更換為含有1 mg/mL 勻黴素之培養基,取得藥劑耐性細胞。
使用細胞分選儀SH800(SONY公司製造),自所取得之藥劑挑選細胞,將與抗FAP抗體西羅珠單抗結合之FAP高表現組分進行單細胞分離。藉由FACS Canto(商標)II(BD Bioscience)確認到所取得之純系之FAP表現。以下,將所獲得之人類FAP表現細胞記載為人類FAP-HEK293。
[實施例3]抗CD40單株抗體及表現載體之取得
1.CD40免疫人類抗體M13噬菌體庫之製作
向人類抗體產生小鼠[Ishida & Lonberg, IBC's 11th Antibody Engineering, Abstract 2000; Ishida, I. et al., Cloning & Stem Cells 4, 85-96 (2002)及石田功(2002)實驗醫學20, 6, 846-851]之腹腔內投予作為免疫原之實施例1中所製作的人類CD40-FLAG-Fc計4次。僅於初次免疫時添加作為佐劑之明礬凝膠(2 mg/隻)及百日咳菌疫苗(1×109
個/隻)。
於初次免疫後之2週後進行第2次免疫,於第2次免疫之1週後進行第3次免疫,於第3次免疫後之10天後進行最終免疫,在最終免疫後之4天後解剖小鼠,外科摘除脾臟。將所摘除之脾臟置於細胞濾網(Falcon公司製造)上,一面利用矽棒輕輕地壓碎一面將細胞移至試管中,進行離心分離後,於冰中使所沈澱之細胞與紅血球去除試劑(Sigma-Aldrich公司製造)反應3分鐘,進而進行離心分離。
使用RNeasy微型套組(QIAGEN公司製造),自所獲得之脾臟細胞提取RNA,利用SMARTer RACE cDNA擴增套組(Clontech公司製造)擴增cDNA,進而利用PCR擴增VH基因片段。將該VH基因片段、與包含為人類之抗體胚系序列且編碼L6序列之胺基酸序列(VL)的序列編號17所示之鹼基序列之VL基因片段一起插入至噬菌粒pCANTAB 5E(Amersham Pharmacia公司製造),將大腸桿菌TG1(Lucigen公司製造)轉形而獲得質體。
再者,L6序列編碼包含序列編號18所示之胺基酸序列之人類抗體之輕鏈可變區(VL),且該VL之CDR1、2及3(分別表示為LCDR1、2及3)之胺基酸序列分別以序列編號19、20及21表示。
使VCSM13抗干擾輔助噬菌體(Agilent Technologies公司製造)感染所獲得之質體,藉此獲得具有包含L6序列之VL基因,且VH基因得到庫化之CD40免疫人類抗體M13噬菌體庫。
2.抗CD40單株抗體之取得
使用該CD40免疫人類抗體M13噬菌體庫,藉由下述噬菌體呈現法,取得具有包含L6胺基酸序列之VL與包含所需胺基酸序列之VH的抗CD40單株抗體。將實施例1中所取得之人類CD40-GST固相化,使人類抗體M13噬菌體庫於室溫下與使用SuperBlock阻斷緩衝液(Thermo Fisher公司製造)阻斷了未結合人類CD40-GST之部位的MAXISORP STARTUBE(NUNC公司製造)反應1~2小時,利用D-PBS(-)及含有0.1% Tween20之PBS(以下記載為PBS-T;和光純藥工業公司製造)分別洗淨3次後,利用0.1 M之甘胺酸-鹽酸鹽緩衝液(Gly-HCl)(pH值2.2)使噬菌體溶出。
使所溶出之噬菌體感染TG1勝任細胞,擴增噬菌體,再次與固相化至MAXISORP STARTUBE之人類CD40-GST進行反應,利用D-PBS(-)及PBS-T分別洗淨5次後,利用0.1 M之甘胺酸-鹽酸鹽緩衝液(Gly-HCl)(pH值2.2)使噬菌體溶出。
將該操作反覆2次或3次,將呈現與人類CD40特異性地結合之scFv之噬菌體進行濃縮。使經濃縮之噬菌體感染TG1,接種至SOBAG培養板(2.0%胰蛋白腖、0.5%酵母萃取液、0.05% NaCl、2.0%葡萄糖、10 mM MgCl2
、100 μg/mL安比西林、1.5%瓊脂)而形成群落。
對群落進行植菌並培養後,使VCSM13抗干擾輔助噬菌體感染其,再次進行培養,藉此獲得單純系之噬菌體。使用所獲得之單純系之噬菌體,利用ELISA選擇與人類及猴CD40-GST均結合之純系。
於ELISA中,將實施例1之人類或猴CD40-GST固相化至各孔,使用利用SuperBlock阻斷緩衝液(Thermo Fisher公司製造)阻斷了未結合人類或猴CD40-GST之部位的MAXISORP(NUNC公司製造)。於各孔中添加各噬菌體純系,於室溫下反應30分鐘後,將各孔利用PBS-T洗淨3次。
繼而,利用含有10%Block Ace(大日本製藥股份有限公司製造)之PBS-T將經辣根過氧化酶標記之抗M13抗體(GE healthcare Bioscience公司製造)稀釋至5000倍,於各孔中各添加50 μL,於室溫下保溫30分鐘。利用PBS-T將微量培養板洗淨4次後,於各孔中添加TMB顯色基質液(DAKO公司製造)各50 μL,於室溫下保溫10分鐘。於各孔中添加2 mol/L之HCl溶液(50 μL/well)以停止顯色反應,利用讀板儀(Emax;Molecular Devices公司)對波長450 nm(參照波長570 nm)下之吸光度進行測定。
針對與人類及猴CD40均結合之純系,進行序列分析,取得具有包含L6之胺基酸序列之VL之抗CD40抗體R1090S55A。將編碼所取得之CD40抗體R1090S55A之VH之完整鹼基序列示於序列編號22,將自該鹼基序列推測之胺基酸序列示於序列編號23,將VH之CDR1~3(以下亦有時記載為HCDR1~3)之胺基酸序列分別示於序列編號24、25、26。
3.抗CD40單株抗體表現載體之取得
製作組入有所取得之抗CD40抗體R1090S55A之基因之可溶性IgG的表現載體。首先,將編碼R1090S55A之VL之L6基因次選殖至N5KG4PE R409K載體(國際公開第2006/033386號中所記載)之BglII-BsiWI位點。
其後,將R1090S55A之VH基因次選殖至N5KG4PE R409K載體之SalI-NheI位點,獲得具有人類IgG4PE R409K之恆定區之抗CD40單株抗體R1090S55A之表現載體即N5KG4PE R409K_R1090S55A。
4.陽性對照抗體表現載體之取得
為了將國際公開第2003/040170號中所記載之抗CD40單株抗體21.4.1(CP870,893)作為抗CD40抗體之陽性對照抗體來製作,而製作表現載體。將21.4.1之VH之鹼基序列示於序列編號27,將自該序列推測之VH之胺基酸序列示於序列編號28。又,將21.4.1之VL之鹼基序列示於序列編號29,將自該序列推測之VL之胺基酸序列示於序列編號30。
合成編碼21.4.1之VH及VL之基因,分別次選殖至N5KG2載體(國際公開第2003/033538號中所記載)之SalI-NheI及BglII-BsiWI位點,獲得具有人類IgG2之恆定區之抗CD40單株抗體21.4.1的表現載體N5KG2_21.4.1。
[實施例4]抗FAP單株抗體及表現載體之取得
1.人類未活化抗體M13噬菌體庫之製作
自源自人類PBMC之cDNA,利用PCR擴增VH基因片段。將該VH基因片段與包含序列編號17所示之L6之鹼基序列之VL基因片段一起插入至將噬菌粒pCANTAB 5E(Amersham Pharmacia公司製造)之標籤序列變更為FLAG-His標籤及胰蛋白酶識別序列[Nature Biotechnoloy 2001, 19, 75-78]的載體,將大腸桿菌TG1(Lucigen公司製造)轉形而獲得質體。
使VCSM13抗干擾輔助噬菌體(Agilent Technologies公司製造)感染所獲得之質體,藉此獲得具有包含L6鹼基序列之VL基因且VH基因得到庫化之人類未活化抗體M13噬菌體庫。
2.FAP免疫人類抗體M13噬菌體庫之製作
向人類抗體產生小鼠[Ishida & Lonberg, IBC's 11th Antibody Engineering, Abstract 2000; Ishida, I. et al., Cloning & Stem Cells 4, 85-96 (2002)及石田功(2002)實驗醫學20, 6, 846-851]投予20 μg/隻或50 μg/隻之作為免疫原之實施例1中所製作之GST-人類FAP、GST-小鼠FAP、或Fc-人類FAP計4次。僅於初次免疫時添加作為佐劑之明礬凝膠(0.25 mg/隻或2 mg/隻)及不活化百日咳菌懸浮液(Nacalai Tesque公司製造)(1×109
個/隻)。
於初次免疫後之2週後進行第2次免疫,於第2次免疫之1週後進行第3次免疫,於第3次免疫後之10天後進行最終免疫,在最終免疫後之4天後解剖小鼠,外科摘除淋巴結或脾臟。將所摘除之淋巴結或脾臟均質化後,通過細胞濾網(Falcon公司製造)而將細胞移至試管中,進行離心分離而使細胞沈澱。所獲得之細胞係與紅血球去除試劑(Sigma-Aldrich公司製造)混合,於37℃之熱水浴中反應1分鐘後,利用DMEM培養基(Sigma-Aldrich公司製造)進行稀釋,進而進行離心分離。
使用RNeasy微型Plus套組(QIAGEN公司製造),自所獲得之淋巴結細胞或脾臟細胞提取RNA,利用SMARTer RACE cDNA擴增套組(Clontech公司製造)擴增cDNA,進而利用PCR擴增VH基因片段。將該VH基因片段與包含序列編號17所示之L6之鹼基序列之VL基因片段一起插入至將噬菌粒pCANTAB 5E(Amersham Pharmacia公司製造)之標籤序列變更為FLAG-His標籤及胰蛋白酶識別序列之載體,將大腸桿菌TG1(Lucigen公司製造)轉形而獲得質體。
使VCSM13抗干擾輔助噬菌體(Agilent Technologies公司製造)感染所獲得之質體,藉此獲得具有包含L6序列之VL基因且VH基因得到庫化之FAP免疫人類抗體M13噬菌體庫。
3.抗FAP單株抗體之取得
使用人類未活化抗體M13噬菌體庫及FAP免疫人類抗體M13噬菌體庫,藉由下述噬菌體呈現法,取得具有包含L6之胺基酸序列之VL之抗FAP單株抗體。將人類FAP-His(BioLegend公司製造)、小鼠FAP(R&D systems公司製造)、或實施例1中所製作之Fc-人類FAP或Fc-小鼠FAP固相化,使人類未活化抗體M13噬菌體庫或FAP免疫人類抗體M13噬菌體庫於室溫下與使用SuperBlock阻斷緩衝液進行了阻斷之MAXISORP STARTUBE反應1~2小時,利用D-PBS(-)及PBS-T洗淨後,利用0.25%胰蛋白酶(Nacalai Tesque公司製造)使噬菌體溶出。使所溶出之噬菌體感染TG1勝任細胞,擴增噬菌體。其後,再次與固相化至MAXISORP STARTUBE之Fc-人類FAP、Fc-小鼠FAP、人類FAP-His、或小鼠FAP進行反應,實施洗淨及溶出。反覆該操作,將呈現與人類FAP及小鼠FAP特異性地結合之抗體分子之噬菌體進行濃縮。使經濃縮之噬菌體感染TG1,接種至SOBAG培養板(2.0%胰蛋白腖、0.5%酵母萃取液、0.05% NaCl、2.0%葡萄糖、10 mM MgCl2
、100 μg/mL安比西林、1.5%瓊脂)而形成群落。
對群落進行植菌並進行培養後,使之感染VCSM13抗干擾輔助噬菌體,再次進行培養,藉此獲得單純系之噬菌體液。使用所獲得之單純系噬菌體液,利用ELISA來選擇與人類及小鼠FAP結合之純系。
於ELISA中,使用如下FAP MAXISORP(NUNC公司製造),其係將抗GST抗體(Rockland公司製造)固相化至各孔,使用SuperBlock阻斷緩衝液(Thermo Fisher公司製造)進行阻斷後,結合人類FAP-His(BioLegend公司製造)、小鼠FAP(R&D systems公司製造)或實施例1中所製作之GST-人類FAP或GST-小鼠而成。於各孔中添加各噬菌體液,於室溫下反應60分鐘後,將各孔利用PBS-T洗淨3次。
繼而,利用含有10%Block Ace(大日本製藥股份有限公司製造)之PBS-T將經辣根過氧化酶標記之抗M13抗體(GE healthcare Bioscience公司製造)稀釋至1000倍,於各孔中各添加50 μL,於室溫下保溫30分鐘。將微量培養板利用PBS-T洗淨3次後,於各孔中各添加50 μL之TMB顯色基質液(DAKO公司製造),於室溫下保溫10分鐘。於各孔中添加2 N之HCl溶液(50 μL/well)以停止顯色反應,利用讀板儀(EnSpire:PerkinElmer公司製造)對波長450 nm(參照波長570 nm)下之吸光度進行測定。
針對與人類及小鼠FAP結合之純系進行序列分析,取得具有包含L6之胺基酸序列之VL之抗FAP抗體。表1中表示編碼所取得之各抗FAP抗體之VH之完整鹼基序列及自該鹼基序列推測之胺基酸序列、以及HCDR1~3之胺基酸序列。
[表1]
抗FAP抗體VH之序列資訊
| 純系名 | VH 鹼基序列 | VH 胺基酸序列 | HCDR1 胺基酸序列 | HCDR2 胺基酸序列 | HCDR3 胺基酸序列 |
| Fa018 | 序列編號31 | 序列編號32 | 序列編號33 | 序列編號34 | 序列編號35 |
| Fa1007 | 序列編號36 | 序列編號37 | 序列編號38 | 序列編號39 | 序列編號40 |
| Fa1013 | 序列編號41 | 序列編號42 | 序列編號43 | 序列編號44 | 序列編號45 |
| Fa1014 | 序列編號46 | 序列編號47 | 序列編號48 | 序列編號49 | 序列編號50 |
| Fa1046 | 序列編號51 | 序列編號52 | 序列編號53 | 序列編號54 | 序列編號55 |
4.陽性對照抗體表現載體之取得
作為抗FAP抗體之陽性對照抗體,製作具有國際公開第2001/68708號所記載之抗FAP單株抗體西羅珠單抗之可變區之抗體。將西羅珠單抗之VH之鹼基序列示於序列編號56,將自該序列推測之VH之胺基酸序列示於序列編號57。又,將西羅珠單抗之VL之鹼基序列示於序列編號58,將自該序列推測之VL之胺基酸序列示於序列編號59。
合成西羅珠單抗之序列編號56所示之VH之鹼基序列及序列編號58所示之VL之鹼基序列的基因片段,插入至pCI載體(Promega公司)之適當之限制酶位點,而取得西羅珠單抗表現載體。
[實施例5]與CD40及FAP結合之雙專一性抗體之製作
利用以下方法製作具有圖1所記載之結構之與人類及猴CD40以及人類FAP之雙專一性抗體。雙專一性抗體之形狀係採用國際公開第2009/131239號所記載之形狀。
以下之步驟中所製作之雙專一性抗體具有作為IgG部分之重鏈恆定區的IgG4PE R409K之CH(將鹼基序列以序列編號60表示,將胺基酸序列以序列編號61表示)、作為與IgG部分之重鏈C末端結合之Fab之CH1的IgG4之CH1(將鹼基序列以序列編號62表示,將胺基酸序列以序列編號63表示)。又,具有包含L6所編碼之VL之輕鏈。
將該雙專一性抗體之名稱、製作該雙專一性抗體時所使用之抗CD40抗體之純系及抗FAP抗體之純系示於表2。
[表2]
CD40-FAP雙專一性抗體所包含之抗體純系
| 雙專一性抗體之名稱 | VH1之純系 | VH2之純系 |
| Ct-R1090-Fa018 | R1090S55A | Fa018 |
| Ct-R1090-Fa1007 | R1090S55A | Fa1007 |
| Ct-R1090-Fa1013 | R1090S55A | Fa1013 |
| Ct-R1090-Fa1014 | R1090S55A | Fa1014 |
| Ct-R1090-Fa1046 | R1090S55A | Fa1046 |
1.雙專一性抗體之表現載體之製作
關於表2所記載之雙專一性抗體,藉由以下所記載之方法製作表現載體。
基於實施例3中所取得之抗CD40抗體R1090S55A、及表2中所記載之抗FAP抗體純系之共通之VL(L6)的胺基酸序列(序列編號18,進行最適合於哺乳類細胞中表現之密碼子轉化,獲得編碼L6胺基酸序列之鹼基序列(序列編號17)。人工合成序列編號17所示之鹼基序列,選殖至包含訊息序列之pCI載體(Promega公司)之適當之限制酶位點,而獲得輕鏈表現載體。
繼而,包含序列編號22所記載之編碼抗CD40抗體R1090S55A之VH之鹼基序列、序列編號60所記載之編碼IgG4PE R409K之CH之鹼基序列、序列編號62所記載之編碼IgG4之CH1之鹼基序列的人工合成基因及經PCR擴增之編碼表1所記載之抗FAP抗體純系之VH之鹼基序列插入至pCI載體(Promega公司),獲得重鏈表現載體。於表3表示編碼所取得之CD40-FAP雙專一性抗體之重鏈之鹼基序列及自該鹼基序列推測之胺基酸序列。
[表3]
CD40-FAP雙專一性抗體所包含之重鏈鹼基序列及胺基酸序列
| 雙專一性抗體名 | 重鏈 鹼基序列 | 重鏈 胺基酸序列 |
| Ct-R1090-Fa018 | 序列編號64 | 序列編號65 |
| Ct-R1090-Fa1007 | 序列編號66 | 序列編號67 |
| Ct-R1090-Fa1013 | 序列編號68 | 序列編號69 |
| Ct-R1090-Fa1014 | 序列編號70 | 序列編號71 |
| Ct-R1090-Fa1046 | 序列編號72 | 序列編號73 |
2.雙專一性抗體之製備
使用實施例5之1.中所製作之雙專一性抗體之表現載體,利用以下之方法使各雙專一性抗體表現並進行純化。
藉由Expi293F(商標)表現系統(Thermo Fisher公司製造),將CD40-FAP雙專一性抗體之重鏈表現載體及包含編碼L6之鹼基序列之輕鏈表現載體共基因導入至Expi293F細胞,於16小時後添加轉染增強劑(Transfection Enhancer),利用短暫性表現系統使雙專一性抗體表現。
於載體導入4天後回收培養上清液,利用孔徑0.22 μm之膜濾器(MILLIPORE製造)進行過濾後,使用蛋白A樹脂(MabSelect、GE healthcare Bioscience公司製造)對抗體進行親和純化。作為洗淨液,使用D-PBS(-)。藉由20 mM檸檬酸鈉-50 mM NaCl緩衝液(pH值3.0)使蛋白A樹脂所吸附之抗體溶出,並將之回收至包含200 mM磷酸鈉緩衝液(pH值7.0)之試管中。
繼而,藉由使用VIVASPIN(Sartrius stealin公司製造)之超過濾,將緩衝液置換為D-PBS(-)後,利用孔徑0.22 μm之膜濾器Millex-Gv(Millipore公司製造)進行過濾殺菌,取得CD40-FAP雙專一性抗體。測定抗體溶液之波長280 nm之吸光度,使用自各抗體之胺基酸序列推測之吸光係數,算出純化抗體之濃度。
[實施例6]對照抗體之製作
分別使用實施例3-3中所製作之N5KG4PE R409K_R1090S55A、實施例3-4中所製作之N5KG2_21.4.1及實施例4-4中所製作之西羅珠單抗之各表現載體,藉由與實施例5-2相同之方法,製作具有IgG4PE R409K之CH之抗CD40單株抗體R1090S55A及21.4.1、以及抗FAP單株抗體西羅珠單抗。
[實施例7]利用ELISA之雙專一性抗體之FAP結合性評價
依據以下之程序,藉由ELISA來評價實施例5中所取得之CD40-FAP雙專一性抗體對於人類FAP之結合性。
將實施例1中所製作之人類FAP-GST以10 μg/mL之濃度利用D-PBS(-)進行稀釋,以50 μL/well分注至96孔NUNC-IMMUNO PLATE(Thermo Fisher公司製造)中,於4℃下靜置一夜。利用200 μL/well之D-PBS(-)將孔洗淨3次後,為了阻斷而以50 μL/well添加1%(w/v)BSA-PBS(-)pH值7.0(Nacalai Tesque公司製造),於室溫下靜置1小時。
將孔中之溶液捨棄,將稀釋至0.00064、0.0032、0.016、0.08、0.4、2、10或50 μg/mL之CD40-FAP雙專一性抗體分別以50 μL/well添加至孔中。於室溫下靜置2小時後,利用200 μL/well之0.05w/v%-tPBS(1x) without KCl(pH值7.2)(Nacalai Tesque公司製造)洗淨3次,添加利用1%(w/v)BSA-PBS(-) pH值7.0(Nacalai Tesque公司製造)將作為2次抗體之Peroxidase AffiniPure山羊抗人IgG, Fcγ Fragment specific(Jackson ImmunoReseach公司製造)稀釋至1/1000倍所得之溶液50 μL/well,並於室溫下靜置1小時。
利用200 μL/well之0.05w/v%-tPBS(1x) without KCl(pH值7.2)(Nacalai Tesque公司製造)將孔洗淨3次後,將等量混合基質試劑盒(R&D systems公司製造)之A液與B液所得者以100 μL/well進行添加並靜置5分鐘。將停止溶液(R&D systems公司製造)以50 μL/well進行添加而使反應停止,使用EPOCH2微型讀板儀(Biotek公司製造)測定450 nm及570 nm之吸光度,計算自450 nm之吸光度減去570 nm之吸光度所得之值。
再者,關於陰性對照,使用IgG4抗體R409K變異體(人類IgG4抗體中,將第409號之R置換為K之抗體)(以下,記載為抗DNP抗體),其係使用編碼同型對照抗體[Clin Cancer Res 2005, 11(8), 3126-3135記載之抗2,4-二硝基酚(DNP)IgG4抗體]之載體並依據實施例5所記載之方法所製作。
圖2中表示對CD40-FAP雙專一性抗體對於人類FAP之結合性進行測定所得之結果。如圖2所示,確認到實施例5中所製作之CD40-FAP雙專一性抗體均與經固相化之人類FAP-GST結合。
[實施例8]利用流式細胞儀評價雙專一性抗體之FAP結合性
實施例5中所製作之CD40-FAP雙專一性抗體對於人類胚性腎細胞HEK293細胞(ATCC No. CRL-1573)及實施例2中所獲得之人類FAP/HEK293細胞之結合性評價係如下述般實施。
使HEK293細胞或FAP/HEK293細胞以2×106
cells/mL之濃度於染色緩衝液(含有1% BSA及2% FBS之D-PBS(-);亦記載為SB)中懸浮,以50 μL/well分注至96孔圓底培養板(Falcon公司製造)中。離心分離(1500 rpm、4℃、3分鐘)後,去除上清液,向沈澱物以最終濃度0.01、0.1、1或10 μg/mL添加50 μL/well之實施例5中所獲得之抗體並進行懸浮後,於冰溫下靜置30分鐘。
進而進行離心分離(2000 rpm、4℃、2分鐘),去除上清液,利用200 μL/well之SB將沈澱物洗淨3次後,將1 μg/mL之山羊F(ab')2
抗人IgG(γ chain specific) Allophycocyanin(APC) Conjugate(Southern Biotech公司製造)以50 μL/well進行添加,於冰溫下保溫30分鐘。利用SB洗淨2次後,於200 μL/well之SB中進行懸浮,利用流式細胞儀FACSCANTO II(Becton Dickinson公司製造)測定各細胞之螢光強度。陰性對照係使用DNP抗體。
圖3(A)中表示對於HEK293進行測定所得之結果,圖3(B)中表示對於人類FAP/HEK293細胞進行測定所得之結果。根據圖3(A)及圖3(B),受驗抗體均未與HEK293細胞結合、或對於HEK293細胞顯示極弱之結合性。另一方面,對於人類FAP/HEK293細胞,以0.01 μg/mL之低濃度結合。
根據以上之結果表示,實施例5中所製作之CD40-FAP雙專一性抗體均為與FAP特異性地結合之抗體。
[實施例9]藉由利用流式細胞分析之CD95表現量分析來評價CD40單株抗體之CD40訊息誘導活性
以Ramos細胞上之CD95之表現量增加作為指標,藉由如下之FCM法對實施例6中所獲得之CD40單株抗體R1090S55A及21.4.1之CD40訊息誘導活性進行評價。
將2×106
cells/mL之Ramos細胞以50 μL/well接種至U底96孔培養板(Falcon公司製造),添加經包含10%FBS之RPMI1640培養基(Sigma-Aldrich公司製造)稀釋之0.2、2或20 μg/ml(最終濃度0.1、1或10 μg/ml)之受驗抗體50 μg/mL,於37℃、5.0%二氧化碳存在下培養16小時。
於離心分離(2000 rpm、4℃、2分鐘)後,將上清液去除,將沈澱物利用200 μL/well之SB洗淨3次。於離心分離(2000 rpm、4℃、2分鐘)後將上清液去除,向沈澱物添加PE小鼠抗人CD95抗體(Becton Dickinson公司製造)並使之懸浮後,於冰溫下靜置30分鐘。
進一步離心分離(2000 rpm、4℃、2分鐘),去除上清液,將沈澱物利用200 μL/well之SB洗淨3次後,使經100倍稀釋之7-胺基放射菌素(7AAD)(Becton Dickinson公司製造)於200 μL/well之SB中懸浮,利用流式細胞儀FACSCANTO II(Becton Dickinson公司製造)測定Ramos細胞上之CD95之螢光強度。陰性對照係使用抗DNP抗體。
於圖4中表示抗CD40抗體21.4.1及R1090S55A之評價結構。根據圖4,促效性抗CD40單株抗體21.4.1與抗DNP抗體相比,增加了Ramos細胞上之CD95之表現量。另一方面,抗CD40抗體單株R1090S55A係與抗DNP抗體相同程度之CD95表現量。
即,表示僅21.4.1為誘導CD40訊息之促效劑抗體,實施例6中所取得之抗CD40單株抗體R1090S55A係不誘導CD40訊息之非促效劑抗體。又,於CD40配位體之存在下進行同樣之試驗,結果為,抗CD40單株抗體R1090S55A不抑制由CD40配位體引起之Ramos細胞上之CD95之表現量的增加(無資料記載)。即,表示抗CD40單株抗體R1090S55A為不抑制由CD40配位體引起之CD40訊息誘導之非拮抗劑性抗體。
[實施例10]藉由利用流式細胞分析之CD95表現量分析來評價CD40-FAP雙專一性抗體的CD40訊息誘導活性
以Ramos細胞上之CD95之表現量增加作為指標,藉由如下述般之FCM法評價實施例5中所獲得之CD40-FAP雙專一性抗體於FAP陽性或陰性細胞共存下之向Ramos細胞的CD40訊息誘導活性。
將4×106
cells/mL之Ramos細胞以25 μL/well接種至U底96孔培養板(Falcon公司製造),將經包含10%FBS之RPMI1640培養基(Sigma-Aldrich公司製造)稀釋之0.002、0.02、0.2、2或20 μg/mL(最終濃度0.001、0.01、0.1、1或10 μg/mL)之受驗抗體以50 μL/well進行添加,進而將HEK293細胞或人類FAP/HEK293細胞4×106
cells/mL以25 μL/well進行添加,於37℃、5.0%二氧化碳存在下培養16小時。
於離心分離(2000 rpm、4℃、2分鐘)後,將上清液去除,將沈澱物利用200 μL/well之SB洗淨3次。於離心分離(2000 rpm、4℃、2分鐘)後,將上清液去除,向沈澱物添加PE小鼠抗人CD95抗體(Becton Dickinson公司製造)及FITC小鼠抗人CD20(Becton Dickinson公司製造)並使其等懸浮後,於冰溫下靜置30分鐘。
進一步離心分離(2000 rpm、4℃、2分鐘),將上清液去除,將沈澱物利用200 μL/well之SB洗淨3次後,使經100倍稀釋之7AAD(Becton Dickinson公司製造)於200 μL/well之SB中懸浮,利用流式細胞儀FACSCANTO II(Becton Dickinson公司製造)對存在於CD20陽性組分中之Ramos細胞上之CD95的螢光強度進行測定。陰性對照係使用抗DNP抗體,陽性對照係使用21.4.1。
將與HEK293細胞共培養之Ramos細胞之結果示於圖5(A),將與人類FAP/HEK293細胞共培養之Ramos細胞之結果示於圖5(B)。
根據圖5(A)、圖5(B),於所有共培養系之實驗中,陰性對照之抗DNP抗體均未使Ramos細胞上之CD95之表現量增加,但陽性對照之抗CD40抗體21.4.1增加了Ramos細胞上之CD95表現量。
又,於如圖5(A)所示般將FAP陰性之HEK293細胞與Ramos細胞共培養之情形時,所有CD40-FAP雙專一性抗體均未誘導CD40訊息。另一方面,於如圖5(B)所示,顯示於將FAP陽性之人類FAP/HEK293細胞與Ramos細胞共培養之情形時,CD40-FAP雙專一性抗體使Ramos細胞上之CD95之表現量增加,誘導了CD40訊息。
該等結果表示,如圖5(A)所示般CD40-FAP雙專一性抗體於單獨地與Ramos細胞上之CD40結合時不會誘導CD40訊息,而如圖5(B)所示般僅於共存FAP陽性細胞時誘導CD40訊息。
如圖4所示,可知確認到作為親本抗體之R1090S55A不會向Ramos細胞誘導CD40訊息,但CD40-FAP雙專一性抗體具有親本抗體所不具有之CD40訊息誘導活性。
藉此提示有如下情況:本發明之CD40-FAP雙專一性抗體於腫瘤或淋巴組織等表現FAP之細胞所存在之部位特異性地向B細胞等CD40陽性細胞誘導訊息。
[實施例11]對於樹狀細胞之CD40訊息誘導活性之評價
以樹狀細胞上之活化標記物、或樹狀細胞所產生之細胞激素之表現量增加作為指標,藉由FCM法及ELISA法,對實施例5中所獲得之CD40-FAP雙專一性抗體於FAP陽性或陰性細胞共存下之對於樹狀細胞之CD40訊息誘導活性進行評價。陰性對照係使用抗DNP抗體,陽性對照組係使用21.4.1。
1.細胞之製備
將Untouched Frozen NPB-CD14+單核球(AllCells公司製造)解凍後,使之於包含10%血清、青黴素-鏈黴素(Nacalai Tesque公司製造)、100 ng/mL之重組人GM-CSF(R&D公司製造)及100 ng/mL之重組人IL4(R&D公司製造)之RPMI1640(Nacalai Tesque公司製造)中以成為1×106
cells/mL之細胞密度的方式進行懸浮,並以3 mL/well添加至6well Flat Bottom Ultra-Low attachment Surface(CORNING公司製造)。
於培養2天及4天後更換為新鮮培養基。於培養開始後之7天後回收細胞,於1500 rpm、室溫下進行5分鐘離心分離。將沈澱細胞於包含100 ng/mL之IL-4及100 ng/mL之GM-CSF之RPMI1640培養基中以成為2×105
cells/mL之方式進行製備,並以250 μL/well添加至24well Flat Bottom Ultra-Low attachment Surface(CORNING公司製造)之各孔中。進而,將人類FAP/HEK293細胞或HEK293細胞以成為4×105
/mL之方式進行製備,以125 μL/well添加至各孔中。將以最終濃度成為0.01、0.1、1、10 μg/mL之方式所製備之評價抗體以125 μL/well進行添加並進行混合,於37℃、5%CO2
存在下培養2天。
2.利用FCM法之樹狀細胞上之活化標記物之表現測定
將經2天培養之細胞進行離心分離(2000 rpm、4℃、5分鐘)。將上清液去除,添加1%(w/v)BSA-PBS(-)、pH值7.0 without KCl(Nacalai公司製造)(亦記載為FACS緩衝液)各150 μL。添加BD Fc Block Reagent for Human (BD Pharmingen公司製造),使之懸浮,於冰上靜置5分鐘。
其後,於96孔U底培養板(FALCON公司製造)之各孔中添加細胞懸浮液50 μL。將懸浮於FACS緩衝液中之標記抗體Brilliant Violet 421抗人CD80抗體(Biolegend公司製造)、PE人CD83抗體(Beckman Coulter公司製造)、PE小鼠抗人CD86(BD Pharmingen公司製造)、FITC小鼠抗人HLA-ABC(BD Pharmingen公司製造)、或APC抗人CD45(Biolegend公司製造)分別以50 μL/well進行添加,並於4℃下保溫30分鐘。
於離心分離(1200 rpm、4℃、2分鐘)後,將上清液進行傾析。將沈澱之細胞利用200 μL之FACS緩衝液洗淨2次。其後,利用100 μL之FACS緩衝液使細胞再懸浮,利用FACS Canto II(BD Biosciences公司製造)對螢光強度進行測定。分析時,使用資料分析軟體FlowJo9.6.4,針對CD45陽性細胞上之CD80、CD83、CD86及HLA-ABC,根據所結合的抗體之平均螢光強度(MFI)來評價表現量。
3.利用ELISA法之樹狀細胞產生IL-12之表現測定
作為樹狀細胞之活化指標,對所產生之IL-12之表現量變現進行驗證。將實施例11-1中經2天培養後之細胞之培養上清液進行回收,依據推薦方法使用人類IL-12 p70 ELISA Ready-SET-Go!(Affymetrix公司製造),對IL-12之表現量進行評價。
4.評價結果
將與HEK293細胞共培養之樹狀細胞之CD83、CD80、CD86、HLA-ABC及IL-12之表現量增加的結果分別示於圖6(A)、圖7(A)、圖8(A)及圖9(A)、圖10(A)。又,將與人類FAP/HEK293細胞共培養之樹狀細胞之CD83、CD80、CD86、HLA-ABC及IL-12之表現量之結果分別示於圖6(B)、圖7(B)、圖8(B)、圖9(B)及圖10(B)。
所有圖中,作為陽性對照之抗CD40單株抗體21.4.1與陰性對照之抗DNP抗體相比,均增加了樹狀細胞所表現之標記物、細胞激素之表現。
又,如圖6(A)、圖7(A)、圖8(A)、圖9(A)及圖10(A)所示,於FAP陰性之HEK293細胞與樹狀細胞之共培養下,所有CD40-FAP雙專一性抗體均不增加樹狀細胞上之CD83、CD80、CD86及HLA-ABC以及可溶性之IL-12的表現量,不會誘導CD40訊息。另一方面,如圖6(B)、圖7(B)、圖8(B)、圖9(B)及圖10(B)所示,顯示於將FAP陽性之人類FAP/HEK293細胞與Ramos細胞共培養之情形時,CD40-FAP雙專一性抗體使樹狀細胞上之CD83、CD80、CD86及HLA-ABC以及可溶性之IL-12之表現量增加,誘導了CD40訊息。
藉此提示有如下情況:本發明之CD40-FAP雙專一性抗體於腫瘤或淋巴組織等表現FAP之細胞所存在之部位特異性地向樹狀細胞等CD40陽性細胞誘導訊息。與實施例10之結果對照,顯示本發明之CD40-FAP雙專一性抗體無論細胞種類為何,均向CD40陽性之免疫細胞誘導CD40訊息。
[實施例12]置換了附加N型糖鏈之胺基酸殘基之CD40-FAP雙專一性抗體之製作
實施例4中所取得之抗FAP抗體純系中,序列編號52所示之Fa1046之重鏈胺基酸序列由於在架構區具有附加N型糖鏈之胺基酸殘基,故藉由以下之方法製作置換了該胺基酸殘基之改變體。
為了將序列編號52所示之FAP抗體純系Fa1046之VH序列中所包含之附加N型糖鏈的胺基酸殘基置換為替代之胺基酸殘基,而進行最適合於哺乳類細胞中表現之密碼子轉化,獲得相對應之鹼基序列。於表4中記載所製作之改變體之純系名、及編碼各VH之鹼基序列及胺基酸序列。
[表4]
置換了附加N結合糖鏈之胺基酸殘基之FAP抗體純系
| 純系名 | VH鹼基序列 | VH胺基酸序列 |
| Fa1046-GTS | 序列編號74 | 序列編號75 |
| Fa1046-FTS | 序列編號76 | 序列編號77 |
| Fa1046-KTS | 序列編號78 | 序列編號79 |
| Fa1046-WTS | 序列編號80 | 序列編號81 |
| Fa1046-TTS | 序列編號82 | 序列編號83 |
關於表4中所記載之各純系,藉由與實施例5相同之方法製作CD40-FAP雙專一性抗體。將該雙專一性抗體之名稱、製作該雙專一性抗體(以下,亦記載為改型雙專一性抗體)時所使用之抗CD40抗體之純系及抗FAP抗體之純系示於表5。表6中表示編碼包含所取得之各抗FAP抗體之VH之CD40-FAP雙專一性抗體之重鏈的鹼基序列及自該鹼基序列推測之胺基酸序列。
[表5]
| 改型雙專一性抗體所使用之純系 | ||
| 雙專一性抗體名 | VH1之純系 | VH2之純系 |
| Ct-R1090-Fa1046-GTS | R1090S55A | Fa1046-GTS |
| Ct-R1090-Fa1046-FTS | R1090S55A | Fa1046-FTS |
| Ct-R1090-Fa1046-KTS | R1090S55A | Fa1046-KTS |
| Ct-R1090-Fa1046-WTS | R1090S55A | Fa1046-WTS |
| Ct-R1090-Fa1046-TTS | R1090S55A | Fa1046-TTS |
[表6]
| 改型雙專一性抗體重鏈之序列資訊 | ||
| 雙專一性抗體之名稱 | 重鏈 鹼基序列 | 重鏈 胺基酸序列 |
| Ct-R1090-Fa1046-GTS | 序列編號84 | 序列編號85 |
| Ct-R1090-Fa1046-FTS | 序列編號86 | 序列編號87 |
| Ct-R1090-Fa1046-KTS | 序列編號88 | 序列編號89 |
| Ct-R1090-Fa1046-WTS | 序列編號90 | 序列編號91 |
| Ct-R1090-Fa1046-TTS | 序列編號92 | 序列編號93 |
[實施例13]使用流式細胞分析法,對置換了附加N型糖鏈之胺基酸殘基之CD40-FAP雙專一性抗體之人類FAP結合性進行評價
藉由與實施例7相同之方法評價實施例12中所獲得之R1090S55A-Fa1046之改型雙專一性抗體對於人類FAP/HEK293細胞及HEK293細胞的結合性。
圖11(A)表示對於HEK293進行測定所得之結果,圖11(B)表示對於人類FAP/HEK293細胞進行測定所得之結果。如圖11(A)所示,受驗抗體均未與FAP陰性之HEK293細胞結合。另一方面,如圖11(B)所示,均自0.01 μg/mL之低濃度起與FAP陽性之人類FAP/HEK293細胞結合,為與改型前之R1090S55A-Fa1046同等以上之活性。
根據以上之結果顯示,實施例12中所製作之R1090S55A-Fa1046之改型雙專一性抗體均為與FAP特異性地結合之抗體。
[實施例14]對於置換了附加N型糖鏈之胺基酸殘基之CD40-FAP雙專一性抗體,藉由使用流式細胞分析法之CD95表現量分析來評價CD40訊息誘導活性
以Ramos細胞上之CD95之表現量增加作為指標,藉由與實施例10相同之方法評價實施例12中所獲得之CD40-FAP雙專一性抗體於FAP陽性或陰性細胞共存下對於Ramos細胞之CD40訊息誘導活性。陰性對照係使用抗DNP抗體,陽性對照係使用21.4.1。
將與HEK293細胞共培養之Ramos細胞之結果示於圖12(A),將與人類FAP/HEK293細胞共培養之Ramos細胞之結果示於圖12(B)。
如圖12(A)所示,於FAP陰性之HEK293細胞及Ramos細胞之共培養下,所有改型雙專一性抗體之CD95之表現量均與抗DNP抗體為相同程度,未誘導CD40訊息。另一方面,如圖12(B)所示,於將FAP陽性之人類FAP/HEK293細胞及Ramos細胞共培養之情形時,改型雙專一性抗體使CD95之表現量增加至與改型前之R1090S55A-Fa1046同等以上。因此,顯示改型雙專一性抗體誘導與改型前之R1090S55A-Fa1046同等以上之CD40訊息。
藉此提示:實施例12中所製作之改型雙專一性抗體於腫瘤或淋巴組織等表現FAP之細胞所存在之部位特異性地向B細胞、樹狀細胞等CD40陽性細胞誘導訊息。
[實施例15]與CD40及FAP結合之雙專一性抗體之製作
依據實施例5記載之方法,製作具有包含表7所示之VH1、VH2之重鏈及含有包含L6之胺基酸序列之VL之輕鏈的雙專一性抗體。表8中表示編碼各FAP-CD40雙專一性抗體之重鏈之鹼基序列及自該鹼基序列推測之胺基酸序列。
[表7]
| FAP-CD40雙專一性抗體所包含之VH純系 | ||
| 雙專一性抗體之名稱 | VH1之純系 | VH2之純系 |
| Ct-Fa018-R1090 | Fa018 | R1090S55A |
| Ct-Fa1007-R1090 | Fa1007 | R1090S55A |
| Ct-Fa1013-R1090 | Fa1013 | R1090S55A |
| Ct-Fa1014-R1090 | Fa1014 | R1090S55A |
| Ct-Fa1046-GTS-R1090 | Fa1046-GTS | R1090S55A |
| Ct-Fa1046-FTS-R1090 | Fa0146-FTS | R1090S55A |
| Ct-Fa1046-KTS-R1090 | Fa0146-KTS | R1090S55A |
| Ct-Fa1046-WTS-R1090 | Fa0146-WTS | R1090S55A |
| Ct-Fa1046-TTS-R1090 | Fa0146-TTS | R1090S55A |
[表8]
| CD40-FAP雙專一性抗體所包含之鹼基序列及胺基酸序列 | ||
| 雙專一性抗體之名稱 | 重鏈 鹼基序列 | 重鏈 胺基酸序列 |
| Ct-Fa018-R1090 | 序列編號94 | 序列編號95 |
| Ct-Fa1007-R1090 | 序列編號96 | 序列編號97 |
| Ct-Fa1013-R1090 | 序列編號98 | 序列編號99 |
| Ct-Fa1014-R1090 | 序列編號100 | 序列編號101 |
| Ct-Fa1046-GTS-R1090 | 序列編號102 | 序列編號103 |
| Ct-Fa1046-FTS-R1090 | 序列編號104 | 序列編號105 |
| Ct-Fa1046-KTS-R1090 | 序列編號106 | 序列編號107 |
| Ct-Fa1046-WTS-R1090 | 序列編號108 | 序列編號109 |
| Ct-Fa1046-TTS-R1090 | 序列編號110 | 序列編號111 |
[實施例16]藉由利用流式細胞分析之CD95表現量分析來評價CD40-FAP雙專一性抗體之CD40訊息誘導活性
與實施例9同樣地,確認實施例15中所製作之CD40-FAP雙專一性抗體之CD40訊息誘導活性。
[實施例17]藉由利用流式細胞分析之CD95表現量分析來評價CD40-FAP雙專一性抗體之CD40訊息誘導活性
以Ramos細胞上之CD95之表現量增加作為指標,藉由與實施例10相同之方法來評價實施例5中所獲得之CD40-FAP雙專一性抗體(Ct-R1090-Fa1007、Ct-R1090-Fa1013、Ct-R1090-Fa1014及Ct-R1090-Fa1046)於FAP陽性纖維母細胞及FAP陽性癌細胞中向Ramos細胞之CD40訊息誘導活性。
但,受驗抗體係以0.02、0.2、2或20 μg/mL(最終濃度0.01、0.1、1或10 μg/mL)之濃度添加。又,使用FAP陽性纖維母細胞株IMR-90(ATCC No. CCL-186)或FAP陽性癌細胞株RPMI-7951(DSMZ No. ACC 66)來代替實施例10中所使用之HEK293細胞或人類FAP/HEK293細胞。
將與IMR-90細胞共培養之Ramos細胞之結果示於圖13(A),將與RPMI-7951細胞共培養之Ramos細胞之結果示於圖13(B)。
其結果為,於所有共培養系之實驗中,陰性對照之抗DNP抗體均未增加Ramos細胞上之CD95之表現,但陽性對照之抗CD40單株抗體21.4.1增加了Ramos細胞上之CD95表現。
如圖13(A)及圖13(B)所示,顯示於將FAP陽性之纖維母細胞或FAP陽性之癌細胞與Ramos細胞共培養之情形時,CD40-FAP雙專一性抗體使Ramos細胞上之CD95之表現量增加,誘導了CD40訊息。
藉此提示,本發明之CD40-FAP雙專一性抗體於腫瘤或淋巴組織等表現FAP之纖維母細胞或癌細胞所存在之部位特異性地向B細胞等CD40陽性細胞誘導訊息。
[實施例18]藉由利用流式細胞分析之CD83及CD95表現量分析來評價CD40-FAP雙專一性抗體之CD40訊息誘導活性
以Ramos細胞上之CD83及CD95之表現量增加作為指標,藉由與實施例10相同之方法評價實施例5及實施例12中所獲得之CD40-FAP雙專一性抗體(Ct-R1090-Fa1013、Ct-R1090-Fa1014及Ct-R1090-Fa1046-GTS)於人類FAP固相化珠粒中向Ramos細胞之CD40訊息誘導活性。
但,受驗抗體係以2 μg/mL(最終濃度1 μg/mL)進行添加,添加下述方法中所製作之人類FAP固相化珠粒以代替實施例10中所使用之HEK293細胞或人類FAP/HEK293細胞。又,作為CD83檢測用抗體,使用APC小鼠抗人CD83抗體(Biolegend公司製造)。
(FAP固相化珠粒之製作方法)
以如下方式製備固相化有人類FAP之Dynabeads M-280 抗生蛋白鏈菌素(Invitrogen公司製造)。藉由Biotin標記套組NH2(同仁化學公司製造)對實施例1中所獲得之人類FAP-FLAG-Fc進行生物素標記,而獲得人類FAP-FLAG-Fc-Biotin。將1 μg之人類FAP-FLAG-Fc-Biotin階段性地稀釋1.5倍,使各自與6.5×106
之Dynabeads M-280 抗生蛋白鏈菌素結合。藉此,獲得了不同之人類FAP量之經固相化之珠粒。
(固相化於珠粒之FAP之量之測定方法)
固相化於珠粒之FAP之量係利用流式細胞儀FACSCANTO II(Becton Dickinson公司製造)測定陰性對照之抗DNP抗體及抗FAP抗體對於FAP之固相化珠粒的結合,以相對螢光強度值(RFI)來算出。
將Ramos細胞上之CD83之表現量之結果示於圖14(A),將Ramos細胞上之CD95之表現量之結果示於圖14(B)。
其結果為,不論固相化於經共培養之珠粒之人類FAP之量如何,陰性對照之抗DNP抗體均不增加Ramos細胞上之CD83及CD95的表現量,但陽性對照之抗CD40單株抗體21.4.1增加了Ramos細胞上之CD83及CD95表現量。
如圖14(A)及圖14(B)所示,顯示CD40-FAP雙專一性抗體Ct-R1090-Fa1013、Ct-R1090-Fa1014及Ct-R1090-Fa1046-GTS於固相化有FAP之珠粒中,與FAP之量相關聯地使Ramos細胞上之CD83及CD95之表現量增加,誘導了CD40訊息。
藉此提示出如下情況:本發明之CD40-FAP雙專一性抗體不論細胞之種類如何,均於在表面上表現FAP之細胞所存在之部位特異性地向B細胞、樹狀細胞等CD40陽性細胞誘導訊息,且該訊息之強度有與FAP之表現量相關之傾向。
[實施例19]作為對照組之CD40-FAP雙專一性抗體之製作
藉由以下之方法製作國際公開第2018/185045號所記載之CD40-FAP雙專一性抗體。
1.雙專一性抗體之表現載體之製作
國際公開第2018/185045號中所記載之抗體純系P1AA9663係將與FAP結合之抗FAP抗體28H1、與抗CD40抗體SGN-40組合而成之CD40-FAP雙專一性抗體,且報告有向CD40陽性細胞誘導訊息(國際公開第2018/185045號、Beatty GL et al., Expert Rev Anticancer Ther. 2017 Feb;17 (2):175-186.)。基於構成P1AA9663之胺基酸序列(序列編號112、113、114),進行最適合於哺乳類細胞中表現之密碼子轉化,獲得鹼基序列(序列編號115、116、117)。人工合成包含序列編號115、116、117所示之鹼基序列之基因片段,將之選殖至包含編碼訊息序列之鹼基序列之pCI載體(Promega公司)之適當限制酶位點,獲得表現載體。
2.雙專一性抗體之製備
使用實施例19之1.中所製作之雙專一性抗體之表現載體,利用以下之方法使雙專一性抗體表現並進行純化。
藉由Expi293F(商標)表現系統(Thermo Fisher公司製造)將構成雙專一性抗體之分子之表現載體共基因導入至Expi293F細胞,於16小時後添加轉染增強劑(Transfection Enhancer),利用短暫性表現系統使雙專一性抗體表現。
於載體導入4天後回收培養上清液,利用孔徑0.22 μm之膜濾器(MILLIPORE製造)進行過濾後,使用蛋白A樹脂(MabSelect、GE healthcare Bioscience公司製造)對抗體進行親和純化。作為洗淨液,使用D-PBS(-)。藉由20 mM檸檬酸鈉-50 mM NaCl緩衝液(pH值3.0)使蛋白A樹脂所吸附之抗體溶出,並回收至包含200 mM磷酸鈉緩衝液(pH值7.0)之試管中。
繼而,藉由使用VIVASPIN(Sartrius stealin公司製造)之超過濾,將緩衝液置換為D-PBS(-)後,利用孔徑0.22 μm之膜濾器Millex-Gv(Millipore公司製造)進行過濾殺菌,而取得CD40-FAP雙專一性抗體P1AA9663。對抗體溶液之波長280 nm之吸光度進行測定,使用自各抗體之胺基酸序列推測之吸光係數,算出純化抗體之濃度。
[實施例20]藉由利用流式細胞分析之CD83表現量分析來評價CD40-FAP雙專一性抗體之CD40訊息誘導活性
以Ramos細胞上之CD83之表現量增加作為指標,利用與實施例10及實施例19相同之方法來評價實施例5、實施例12及實施例19中所獲得之CD40-FAP雙專一性抗體(Ct-R1090-Fa1013、Ct-R1090-Fa1014、Ct-R1090-Fa1046-GTS及P1AA9663)於FAP陽性細胞存在下對Ramos細胞之CD40訊息誘導活性。
但,受驗抗體係以0.02、0.2、2或20 μg/mL(最終濃度0.01、0.1、1或10 μg/mL)之濃度添加,使用FAP陽性纖維母細胞株SC-00A05(NEUROMICS公司製造)以代替實施例10之HEK293細胞或人類FAP/HEK293細胞。
將Ramos細胞之CD83之表現量變化之結果示於圖15。其結果為,於所有共培養系之實驗中,陰性對照之抗DNP抗體均未增加Ramos細胞上之CD83之表現量,但陽性對照之抗CD40抗體21.4.1增加了Ramos細胞上之CD83之表現量。
顯示CD40-FAP雙專一性抗體Ct-R1090-Fa1013、Ct-R1090-Fa1014及Ct-R1090-Fa1046-GTS均增加了Ramos細胞上之CD83之表現量,誘導了CD40訊息。又,顯示該抗體較對照組之CD40-FAP雙專一性抗體P1AA9663增加CD83之表現量,更強地誘導CD40訊息。
藉此提示了如下情況:本發明之CD40-FAP雙專一性抗體強於既有之CD40-FAP雙專一性抗體,對B細胞等CD40陽性細胞誘導訊息。
[實施例21]CD40-FAP雙專一性抗體對於樹狀細胞之CD40訊息誘導活性之評價
以樹狀細胞之活化標記物即CD83、CD86及HLA-ABC之表現量增加作為指標,藉由與實施例11相同之方法評價實施例5、實施例12及實施例19中所獲得之CD40-FAP雙專一性抗體(Ct-R1090-Fa1013、Ct-R1090-Fa1046-GTS及P1AA9663)於FAP陽性細胞共存下向樹狀細胞之CD40訊息誘導活性。
但,使用FAP陽性纖維母細胞株SC-00A05(NEUROMICS公司製造)以代替實施例11之人類FAP/HEK293細胞或HEK293細胞。又,CD86之檢測用抗體係使用PE-Cy7小鼠抗人CD86(Biolegend公司製造)。
將樹狀細胞上之CD83、CD86、及HLA-ABC之表現量變化之結果分別示於圖16(A)、圖16(B)及圖16(C)。
根據圖16(A)、圖16(B)及圖16(C),不論何種標記物,作為陰性對照之抗DNP抗體均未使表現量增加,相對於此,作為陽性對照之抗CD40單株抗體21.4.1使表現量增加。
又,不論何種標記物,均與P1AA9663相比,Ct-R1090-Fa1013及Ct-R1090-Fa1046-GTS使表現量增加,Ct-R1090-Fa1013及Ct-R1090-Fa1046-GTS與P1AA9663相比,較強地誘導CD40訊息。
藉此揭示有如下情況:本發明之CD40-FAP雙專一性抗體與既有之CD40-FAP雙專一性抗體相比,對樹狀細胞等CD40陽性細胞較強地誘導CD40訊息。
[實施例22]於交聯抗體存在下藉由利用流式細胞分析之CD83及CD95表現量分析來評價CD40-FAP雙專一性抗體的CD40訊息誘導活性
為了確認有無FAP表現細胞非依賴性之基於抗體交聯之CD40訊息誘導活性,利用與實施例10相同之方法,以Ramos細胞上之CD83及CD95之表現量增加作為指標,評價實施例5及實施例19中所獲得之CD40-FAP雙專一性抗體(Ct-R1090-Fa1013及P1AA9663)於交聯抗體存在下對Ramos細胞之CD40訊息誘導活性。
但,實施例10之HEK293細胞或人類FAP/HEK293細胞未使用,添加受驗抗體後,將製備成80 μg/mL(最終濃度20 μg/mL)之交聯抗體抗人IgG(FcγR specific)(Jackson Immunoresearch公司製造)以25 μL/well進行添加。
又,CD83檢測用抗體係使用APC小鼠抗人CD83抗體(Biolegend公司製造)。
將Ramos細胞之CD83之表現量變化之結果示於圖17(A),將CD95之表現量之結果示於圖17(B)。
根據圖17(A)及(B),陰性對照之抗DNP抗體未增加Ramos細胞上之CD83及CD95之表現量。
又,對照組之P1AA9663於觀察到FAP依賴性之促效劑活性之0.1 μg/mL之濃度下,明顯增加了CD83及CD95之表現量,另一方面,Ct-R1090-Fa1013於相同濃度下未明顯地增加CD83及CD95之表現量。因此,顯示於交聯抗體之存在下,P1AA9663較強地誘導FAP表現細胞非依賴性之CD40訊息,另一方面,Ct-R1090-Fa1013未明顯地誘導FAP表現細胞非依賴性之CD40訊息。
根據以上提示有如下情況:於生物體內藉由抗體等而交聯之情形時,既有之CD40-FAP雙專一性抗體即便於表現FAP之細胞不存在之部位亦有可能較強地誘導CD40訊息,另一方面,本發明之CD40-FAP雙專一性抗體於表現FAP之細胞不存在之部位有可能不會誘導CD40訊息。
已使用特定態樣對本發明詳細地進行了說明,但業者明確,能夠不脫離本發明之意圖與範圍而進行各種變更及變化。再者,本申請案係基於2019年5月15日提出申請之日本專利申請(特願2019-092298號),且藉由引用將其全部內容援用於本文中。
[序列表非關鍵文字]
序列編號19:人工序列之記載:L6之CDR1胺基酸序列
序列編號20:人工序列之記載:L6之CDR2胺基酸序列
序列編號21:人工序列之記載:L6之CDR3胺基酸序列
序列編號24:人工序列之記載:R1090之HCDR1之胺基酸序列
序列編號25:人工序列之記載:R1090之HCDR2之胺基酸序列
序列編號26:人工序列之記載:R1090之HCDR3之胺基酸序列
序列編號33:人工序列之記載:Fa018之HCDR1之胺基酸序列
序列編號34:人工序列之記載:Fa018之HCDR2之胺基酸序列
序列編號35:人工序列之記載:Fa018之HCDR3之胺基酸序列
序列編號38:人工序列之記載:Fa1007之HCDR1之胺基酸序列
序列編號39:人工序列之記載:Fa1007之HCDR2之胺基酸序列
序列編號40:人工序列之記載:Fa1007之HCDR3之胺基酸序列
序列編號43:人工序列之記載:Fa1013之HCDR1之胺基酸序列
序列編號44:人工序列之記載:Fa1013之HCDR2之胺基酸序列
序列編號45:人工序列之記載:Fa1013之HCDR3之胺基酸序列
序列編號48:人工序列之記載:Fa1014之HCDR1之胺基酸序列
序列編號49:人工序列之記載:Fa1014之HCDR2之胺基酸序列
序列編號50:人工序列之記載:Fa1014之HCDR3之胺基酸序列
序列編號53:人工序列之記載:Fa1046之HCDR1之胺基酸序列
序列編號54:人工序列之記載:Fa1046之HCDR2之胺基酸序列
序列編號55:人工序列之記載:Fa1046之HCDR3之胺基酸序列
序列編號56:人工序列之記載:西羅珠單抗之VH之鹼基序列
序列編號57:人工序列之記載:西羅珠單抗之VH之胺基酸序列
序列編號58:人工序列之記載:西羅珠單抗之VL之鹼基序列
序列編號59:人工序列之記載:西羅珠單抗之VL之胺基酸序列
序列編號60:人工序列之記載:IgG4PE R409K之CH之鹼基序列
序列編號61:人工序列之記載:IgG4PE R409K之CH之胺基酸序列
序列編號64:人工序列之記載:Ct-R1090-Fa018之重鏈之鹼基序列
序列編號65:人工序列之記載:Ct-R1090-Fa018之重鏈之胺基酸序列
序列編號66:人工序列之記載:Ct-R1090-Fa1007之重鏈之鹼基序列
序列編號67:人工序列之記載:Ct-R1090-Fa1007之重鏈之胺基酸序列
序列編號68:人工序列之記載:Ct-R1090-Fa1013之重鏈之鹼基序列
序列編號69:人工序列之記載:Ct-R1090-Fa1013之重鏈之胺基酸序列
序列編號70:人工序列之記載:Ct-R1090-Fa1014之重鏈之鹼基序列
序列編號71:人工序列之記載:Ct-R1090-Fa1014之重鏈之胺基酸序列
序列編號72:人工序列之記載:Ct-R1090-Fa1046之重鏈之鹼基序列
序列編號73:人工序列之記載:Ct-R1090-Fa1046之重鏈之胺基酸序列
序列編號74:人工序列之記載:Fa1046-GTS之VH之鹼基序列
序列編號75:人工序列之記載:Fa1046-GTS之VH之胺基酸序列
序列編號76:人工序列之記載:Fa1046-FTS之VH之鹼基序列
序列編號77:人工序列之記載:Fa1046-FTS之VH之胺基酸序列
序列編號78:人工序列之記載:Fa1046-KTS之VH之鹼基序列
序列編號79:人工序列之記載:Fa1046-KTS之VH之胺基酸序列
序列編號80:人工序列之記載:Fa1046-WTS之VH之鹼基序列
序列編號81:人工序列之記載:Fa1046-WTS之VH之胺基酸序列
序列編號82:人工序列之記載:Fa1046-TTS之VH之鹼基序列
序列編號83:人工序列之記載:Fa1046-TTS之VH之胺基酸序列
序列編號84:人工序列之記載:Ct-R1090-Fa1046-GTS之重鏈之鹼基序列
序列編號85:人工序列之記載:Ct-R1090-Fa1046-GTS之重鏈之胺基酸序列
序列編號86:人工序列之記載:Ct-R1090-Fa1046-FTS之重鏈之鹼基序列
序列編號87:人工序列之記載:Ct-R1090-Fa1046-FTS之重鏈之胺基酸序列
序列編號88:人工序列之記載:Ct-R1090-Fa1046-KTS之重鏈之鹼基序列
序列編號89:人工序列之記載:Ct-R1090-Fa1046-KTS之重鏈之胺基酸序列
序列編號90:人工序列之記載:Ct-R1090-Fa1046-WTS之重鏈之鹼基序列
序列編號91:人工序列之記載:Ct-R1090-Fa1046-WTS之重鏈之胺基酸序列
序列編號92:人工序列之記載:Ct-R1090-Fa1046-TTS之重鏈之鹼基序列
序列編號93:人工序列之記載:Ct-R1090-Fa1046-TTS之重鏈之胺基酸序列
序列編號94:人工序列之記載:Ct-Fa018-R1090之重鏈之鹼基序列
序列編號95:人工序列之記載:Ct-Fa018-R1090之重鏈之胺基酸序列
序列編號96:人工序列之記載:Ct-Fa1007-R1090之重鏈之鹼基序列
序列編號97:人工序列之記載:Ct-Fa1007-R1090之重鏈之胺基酸序列
序列編號98:人工序列之記載:Ct-Fa1013-R1090之重鏈之鹼基序列
序列編號99:人工序列之記載:Ct-Fa1013-R1090之重鏈之胺基酸序列
序列編號100:人工序列之記載:Ct-Fa1014-R1090之重鏈之序列
序列編號101:人工序列之記載:Ct-Fa1014-R1090之重鏈之胺基酸序列
序列編號102:人工序列之記載:Ct-Fa1046-GTS-R1090之重鏈之鹼基序列
序列編號103:人工序列之記載:Ct-Fa1046-GTS-R1090之重鏈之胺基酸序列
序列編號104:人工序列之記載:Ct-Fa1046-FTS-R1090之重鏈之鹼基序列
序列編號105:人工序列之記載:Ct-Fa1046-FTS-R1090之重鏈之胺基酸序列
序列編號106:人工序列之記載:Ct-Fa1046-KTS-R1090之重鏈之鹼基序列
序列編號107:人工序列之記載:Ct-Fa1046-KTS-R1090之重鏈之胺基酸序列
序列編號108:人工序列之記載:Ct-Fa1046-WTS-R1090之重鏈之鹼基序列
序列編號109:人工序列之記載:Ct-Fa1046-WTS-R1090之重鏈之胺基酸序列
序列編號110:人工序列之記載:Ct-Fa1046-TTS-R1090之重鏈之鹼基序列
序列編號111:人工序列之記載:Ct-Fa1046-TTS-R1090之重鏈之胺基酸序列
序列編號112:人工序列之記載:P1AA9663之構成分子1之胺基酸序列
序列編號113:人工序列之記載:P1AA9663之構成分子2之胺基酸序列
序列編號114:人工序列之記載:P1AA9663之構成分子3之胺基酸序列
序列編號115:人工序列之記載:P1AA9663之構成分子1之鹼基序列
序列編號116:人工序列之記載:P1AA9663之構成分子2之鹼基序列
序列編號117:人工序列之記載:P1AA9663之構成分子3之鹼基序列
圖1係表示作為本發明之雙專一性抗體之結構之一例的於IgG部分之重鏈之C末端直接結合有Fab之雙專一性抗體。
圖2係表示藉由ELISA法對各種CD40-FAP雙專一性抗體與人類FAP-GST之結合進行評價所得之結果。縱軸表示吸光度,各CD40-FAP雙專一性抗體係以10、2、0.4、0.08、0.016、0.0032、或0.00064 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體。
圖3(A)係表示藉由流式細胞分析法對各種CD40-FAP雙專一性抗體對於HEK293細胞之結合活性進行評價所得之結果。圖3(B)係表示藉由流式細胞分析法對各種CD40-FAP雙專一性抗體與表現人類FAP之HEK293細胞的結合進行評價所得之結果。各圖之縱軸表示基於平均螢光強度(MFI)之結合活性。各雙專一性抗體係以10、1、0.1或0.01 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體。
圖4係表示R1090S55A對於Ramos細胞之以CD95之表現量增加作為指標的CD40訊息誘導活性。縱軸表示平均螢光強度(MFI),表示CD95之表現量。各雙專一性抗體係以10、1、或0.1 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體,使用促效性抗CD40抗體即21.4.1作為陽性對照抗體。
圖5(A)係表示各種CD40-FAP雙專一性抗體對於與HEK293共培養之Ramos細胞之以CD95之表現量增加作為指標的CD40訊息誘導活性。圖5(B)係表示各種CD40-FAP雙專一性抗體對於與表現人類FAP之HEK293共培養之Ramos細胞之以CD95之表現量增加作為指標的CD40訊息誘導活性。各圖之縱軸表示平均螢光強度(MFI),表示CD95之表現量。各雙專一性抗體係以10、1、0.1、0.01或0.001 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體,使用促效性抗CD40抗體即21.4.1作為陽性對照抗體。
圖6(A)係表示各種CD40-FAP雙專一性抗體對於與HEK293共培養之樹狀細胞之以CD83之表現量增加作為指標的CD40訊息誘導活性。圖6(B)係表示各種CD40-FAP雙專一性抗體對於與表現人類FAP之HEK293共培養之樹狀細胞之以CD83之表現量增加作為指標的CD40訊息誘導活性。各圖之縱軸表示平均螢光強度(MFI),表示CD83之表現量。各雙專一性抗體係以10、1、0.1或0.01 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體,使用促效性抗CD40抗體即21.4.1作為陽性對照抗體。
圖7(A)係表示各種CD40-FAP雙專一性抗體對於與HEK293共培養之樹狀細胞之以CD80之表現量增加作為指標的CD40訊息誘導活性。縱軸表示平均螢光強度(MFI),表示CD80之表現量。圖7(B)係表示各種CD40-FAP雙專一性抗體對於與表現人類FAP之HEK293共培養之樹狀細胞之以CD80之表現量增加作為指標的CD40訊息誘導活性。各圖之縱軸表示螢光強度,表示CD80之表現量。各雙專一性抗體係以10、1、0.1或0.01 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體,使用促效性抗CD40抗體即21.4.1作為陽性對照抗體。
圖8(A)係表示各種CD40-FAP雙專一性抗體對於與HEK293共培養之樹狀細胞之以CD86之表現量增加作為指標的CD40訊息誘導活性。圖8(B)係表示各種CD40-FAP雙專一性抗體對於與表現人類FAP之HEK293共培養之樹狀細胞之以CD86之表現量增加作為指標的CD40訊息誘導活性。各圖之縱軸表示平均螢光強度(MFI),表示CD86之表現量。各雙專一性抗體係以10、1、0.1或0.01 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體,使用促效性抗CD40抗體即21.4.1作為陽性對照抗體。
圖9(A)係表示各種CD40-FAP雙專一性抗體對於與HEK293共培養之樹狀細胞之以HLA-ABC之表現量增加作為指標的CD40訊息誘導活性。圖9(B)係表示各種CD40-FAP雙專一性抗體對於與表現人類FAP之HEK293共培養之樹狀細胞之以HLA-ABC之表現量增加作為指標的CD40訊息誘導活性。各圖之縱軸表示平均螢光強度(MFI),表示HLA-ABC之表現量。各雙專一性抗體係以10、1、0.1或0.01 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體,使用促效性抗CD40抗體即21.4.1作為陽性對照抗體。
圖10(A)係表示對於與HEK293共培養之樹狀細胞之各種CD40-FAP雙專一性抗體之以IL-12之濃度增加(表現量增加)作為指標的CD40訊息誘導活性。圖10(B)係表示對於與表現人類FAP之HEK293共培養之樹狀細胞之各種CD40-FAP雙專一性抗體之以IL-12之濃度增加(表現量增加)作為指標的CD40訊息誘導活性。各圖之縱軸表示IL-12之濃度(pg/mL)。各雙專一性抗體係以10、1、0.1或0.01 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體,使用促效性抗CD40抗體即21.4.1作為陽性對照抗體。
圖11(A)係表示藉由流式細胞分析法對各種CD40-FAP雙專一性抗體對於HEK293細胞之結合活性進行評價所得之結果。圖11(B)係表示藉由流式細胞分析法對各種CD40-FAP雙專一性抗體對於表現人類FAP之HEK293細胞之結合進行評價所得之結果。各圖之縱軸表示平均螢光強度(MFI),作為結合活性之指標。各雙專一性抗體係以10、1、0.1或0.01 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗。
圖12(A)係表示各種CD40-FAP雙專一性抗體對於與HEK293共培養之Ramos細胞之以CD95之表現量增加作為指標的CD40訊息誘導活性。圖12(B)係表示各種CD40-FAP雙專一性抗體對於與表現人類FAP之HEK293共培養之Ramos細胞之以CD95之表現量增加作為指標的CD40訊息誘導活性。各圖之縱軸表示平均螢光強度(MFI),表示CD95之表現量。各雙專一性抗體係以10、1、0.1、0.01或0.001 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體,使用促效性抗CD40抗體即21.4.1作為陽性對照抗體。
圖13(A)及圖13(B)係表示各種CD40-FAP雙專一性抗體對於與FAP陽性細胞株共培養之Ramos細胞之以CD95之表現量增加作為指標的CD40訊息誘導活性。作為FAP陽性細胞株,於圖13(A)中使用纖維母細胞株IMR90,於圖13(B)中使用黑色素瘤細胞株RPMI7951。各圖之縱軸表示平均螢光強度(MFI),表示CD95之表現量。各雙專一性抗體係以10、1、0.1或0.01 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體,使用促效性抗CD40抗體即21.4.1作為陽性對照抗體。
圖14(A)及圖14(B)係表示各種CD40-FAP雙專一性抗體對於與固相化有FAP之珠粒共培養之Ramos細胞的CD40訊息誘導活性。圖14(A)及圖14(B)分別以CD83及CD95之表現量增加作為CD40訊息誘導活性之指標。各圖之縱軸表示平均螢光強度(MFI),表示CD83或CD95之表現量。各圖之橫軸表示抗FAP抗體相對於作為陰性對照抗體之抗DNP抗體的相對螢光強度(RFI),表示於經共培養之各珠粒中之FAP之表現量。各雙專一性抗體係以1 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體,使用促效性抗CD40抗體即21.4.1作為陽性對照抗體。
圖15係表示各種CD40-FAP雙專一性抗體對於與纖維母細胞株SC00A05共培養之Ramos細胞之以CD83之表現量增加作為指標的CD40訊息誘導活性。圖之縱軸表示平均螢光強度(MFI),表示CD83之表現量。各雙專一性抗體係以10、1、0.1或0.01 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體,使用促效性抗CD40抗體即21.4.1作為陽性對照抗體。
圖16(A)~(C)係表示各種CD40-FAP雙專一性抗體對於與纖維母細胞株SC00A05共培養之樹狀細胞的CD40訊息誘導活性。圖16(A)~(C)中,以CD83、CD86及HLA-ABC各自之表現量增加作為CD40訊息誘導活性之指標。各圖之縱軸表示平均螢光強度(MFI),分別表示CD83、CD86及HLA-ABC之表現量。各雙專一性抗體係以10、1、0.1或0.01 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體,使用促效性抗CD40抗體即21.4.1作為陽性對照抗體。
圖17(A)及圖17(B)係表示在添加有交聯抗體之條件下各種CD40-FAP雙專一性抗體對於Ramos細胞之FAP表現細胞非依賴性之CD40訊息誘導活性。圖17(A)及圖17(B)中,分別以CD83及CD95之表現量增加作為CD40訊息誘導活性之指標。各圖之縱軸表示平均螢光強度(MFI),表示CD83或CD95之表現量。各雙專一性抗體係以0.1、0.01或0.001 μg/mL添加。比較對象係使用抗DNP抗體作為陰性對照抗體。
















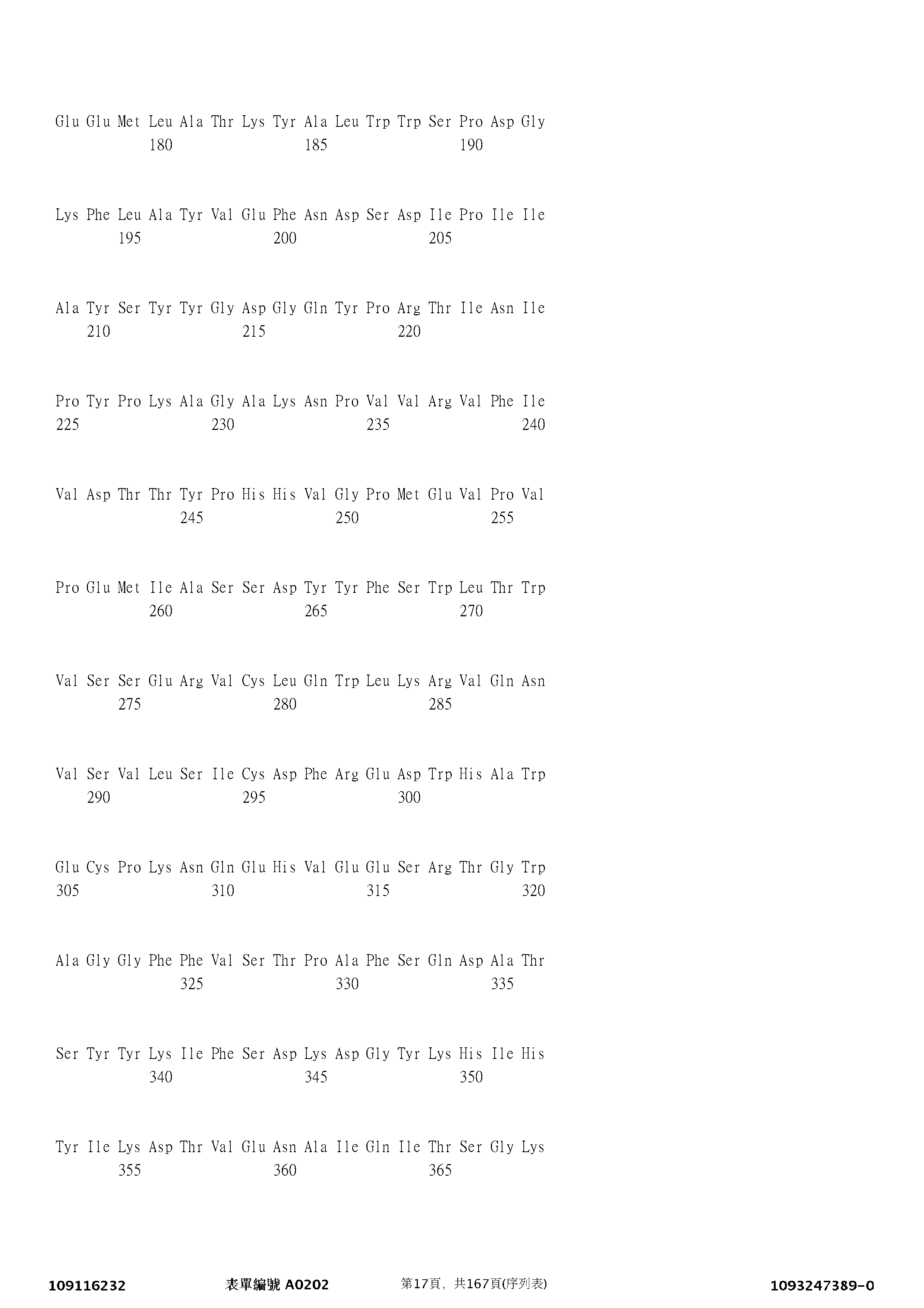




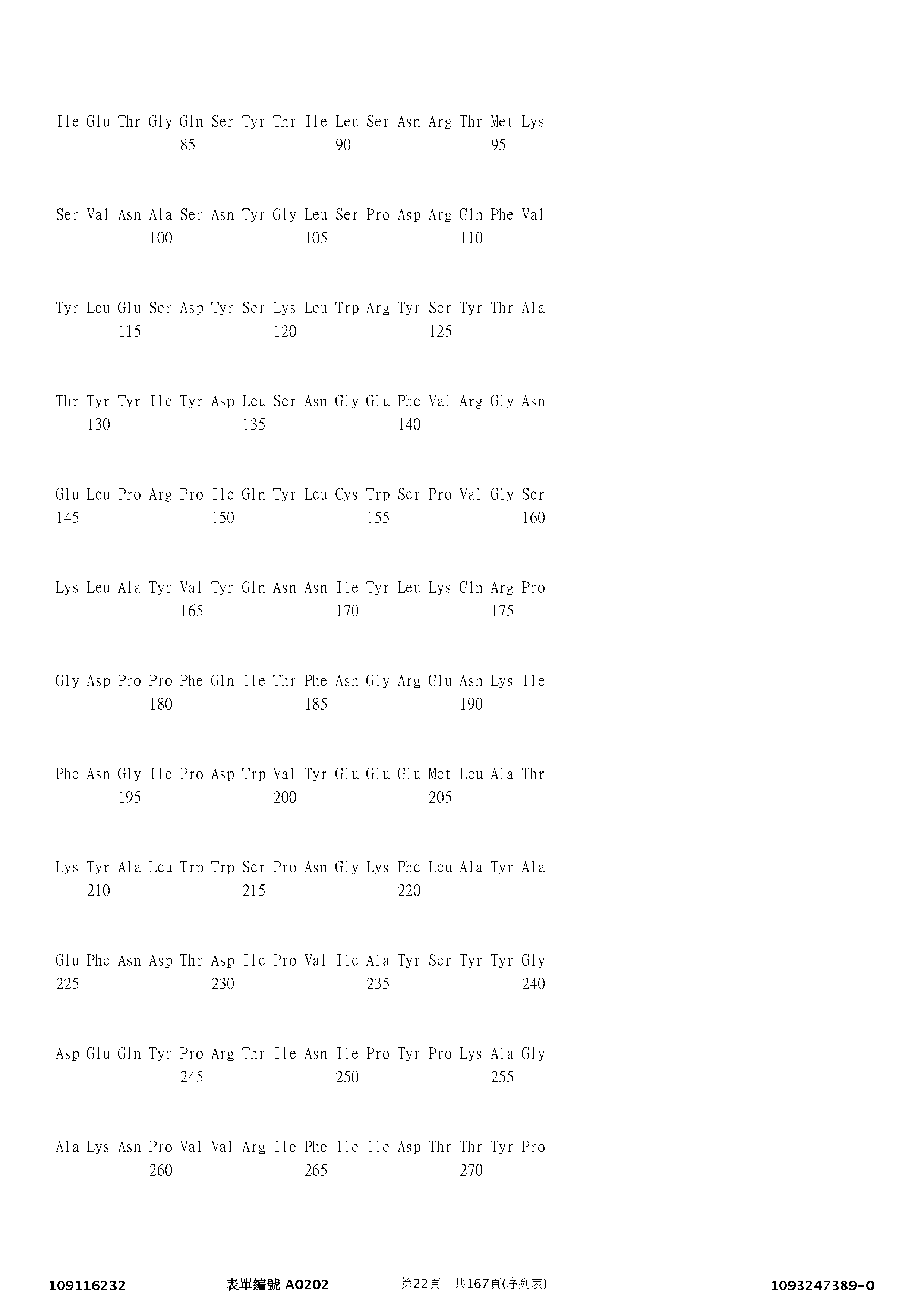



















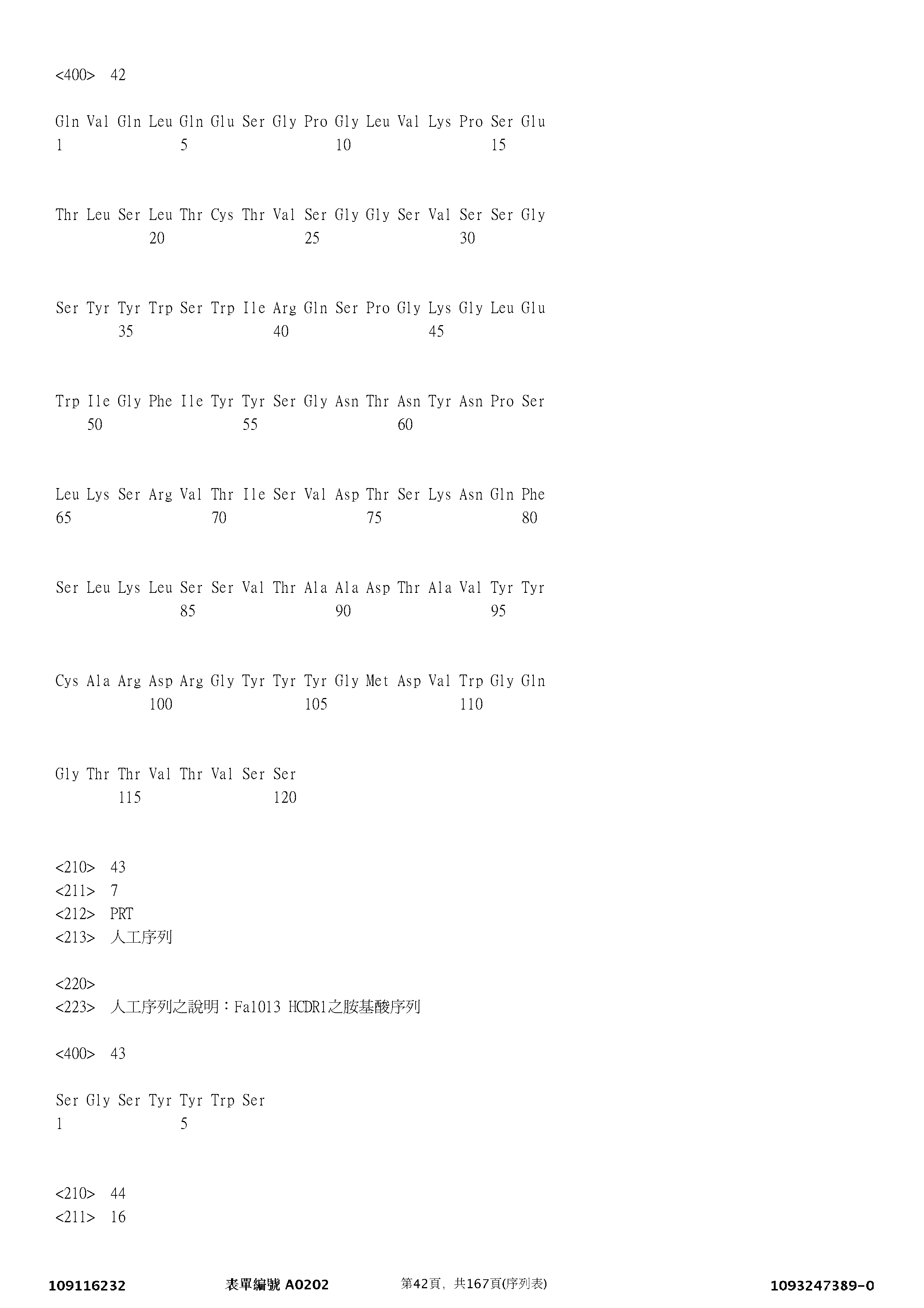




























































































































Claims (32)
- 一種雙專一性抗體,其係含有包含第一抗原結合域之IgG部分及第二抗原結合域且與人類CD40及人類纖維母細胞活化蛋白α(FAP)結合者,該IgG部分之重鏈之C末端直接或經由連接子與該第二抗原結合域結合。
- 如請求項1之雙專一性抗體,其中上述第一抗原結合域及上述第二抗原結合域之任一者源自非促效性抗CD40抗體。
- 如請求項1或2之雙專一性抗體,其以二價分別與人類CD40及人類FAP結合。
- 如請求項1至3中任一項之雙專一性抗體,其中上述IgG部分之重鏈之C末端與上述第二抗原結合域直接結合。
- 如請求項1至4中任一項之雙專一性抗體,其具有CD40促效劑活性。
- 如請求項1至5中任一項之雙專一性抗體,其於不存在FAP表現細胞下不顯示CD40促效劑活性,僅於FAP表現細胞之存在下顯示CD40促效劑活性。
- 如請求項1至6中任一項之雙專一性抗體,其中上述第一抗原結合域及上述第二抗原結合域之任一者為抗CD40 IgG抗體之Fab(抗CD40 Fab)。
- 如請求項1至7中任一項之雙專一性抗體,其中上述第一抗原結合域及上述第二抗原結合域之任一者為抗FAP抗體之Fab(抗FAP Fab)。
- 如請求項1至8中任一項之雙專一性抗體,其中上述第一抗原結合域為抗CD40 Fab,上述第二抗原結合域為抗FAP Fab。
- 如請求項1至8中任一項之雙專一性抗體,其中上述第一抗原結合域為抗FAP Fab,上述第二抗原結合域為抗CD40 Fab。
- 9或10之雙專一性抗體,其中上述抗CD40 Fab為包含如下重鏈可變區(VH)及輕鏈可變區(VL)之Fab,該重鏈可變區(VH)含有分別包含序列編號24~26所表示之胺基酸序列之互補決定區(CDR)1~3,該輕鏈可變區(VL)含有分別包含序列編號19~21所表示之胺基酸序列之CDR1~3。
- 如請求項7及9至11中任一項之雙專一性抗體,其中上述抗CD40 Fab為包含如下VH及VL之Fab,該VH包含序列編號23所表示之胺基酸序列,該VL包含序列編號18所表示之胺基酸序列。
- 如請求項8至12中任一項之雙專一性抗體,其中上述抗FAP Fab為包含如下VH及VL之Fab,該VH含有選自以下(i)~(v)之任一個重鏈CDR1~3,該VL含有分別包含序列編號19~21所表示之胺基酸序列之CDR1~3; (i)分別包含序列編號33~35所表示之胺基酸序列之重鏈CDR1~3、 (ii)分別包含序列編號38~40所表示之胺基酸序列之重鏈CDR1~3、 (iii)分別包含序列編號43~45所表示之胺基酸序列之重鏈CDR1~3、 (iv)分別包含序列編號48~50所表示之胺基酸序列之重鏈CDR1~3及 (v)分別包含序列編號53~55所表示之胺基酸序列之重鏈CDR1~3。
- 如請求項8至13中任一項之雙專一性抗體,其中上述抗FAP Fab為包含如下VH及VL之Fab,該VH包含選自序列編號32、37、42、47、52、75、77、79、81及83之任一者所表示之胺基酸序列,該VL包含序列編號18所表示之胺基酸序列。
- 如請求項1至14中任一項之雙專一性抗體,其中上述IgG部分含有包含序列編號61所表示之胺基酸序列之重鏈恆定區。
- 如請求項1至9及11至15中任一項之雙專一性抗體,其包含2條重鏈及4條輕鏈,該重鏈包含選自序列編號65、67、69、71、73、85、87、89、91及93之任一者所表示之胺基酸序列,該輕鏈含有包含序列編號18所表示之胺基酸序列之VL。
- 如請求項1至8及10至15中任一項之雙專一性抗體,其包含2條重鏈及4條輕鏈,該重鏈包含選自序列編號95、97、99、101、103、105、107、109及111之任一者所表示之胺基酸序列,該輕鏈含有包含序列編號18所表示之胺基酸序列之VL。
- 一種雙專一性抗體片段,其係如請求項1至17中任一項之雙專一性抗體之雙專一性抗體片段。
- 一種DNA,其編碼如請求項1至17中任一項之雙專一性抗體或如請求項18之雙專一性抗體片段。
- 一種重組體載體,其含有如請求項19之DNA。
- 一種轉形株,其係將如請求項20之重組體載體導入至宿主細胞而獲得。
- 一種如請求項1至17中任一項之雙專一性抗體或如請求項18之雙專一性抗體片段之製造方法,其特徵在於:將如請求項21之轉形株於培養基中進行培養,於培養物中生產蓄積如請求項1至17中任一項之雙專一性抗體或如請求項18之雙專一性抗體片段,自該培養物採集雙專一性抗體或如請求項18之雙專一性抗體片段。
- 一種與CD40及FAP之至少一者相關之疾病之治療藥及/或診斷藥,其含有如請求項1至17中任一項之雙專一性抗體或如請求項18之雙專一性抗體片段作為有效成分。
- 如請求項23之治療藥及/或診斷藥,其中與CD40及FAP之至少一者相關之疾病為癌症。
- 一種與CD40及FAP之至少一者相關之疾病之治療方法及/或診斷方法,其使用如請求項1至17中任一項之雙專一性抗體或如請求項18之雙專一性抗體片段。
- 如請求項25之方法,其中與CD40及FAP之至少一者相關之疾病為癌症。
- 一種如請求項1至17中任一項之雙專一性抗體或如請求項18之雙專一性抗體片段,其用以供使用於治療及/或診斷與CD40及FAP之至少一者相關之疾病。
- 如請求項27之雙專一性抗體或該雙專一性抗體片段,其中與CD40及FAP之至少一者相關之疾病為癌症。
- 一種如請求項1至17中任一項之雙專一性抗體或如請求項18之雙專一性抗體片段之用途,其係用於製造與CD40及FAP之至少一者相關之疾病之治療劑及/或診斷劑。
- 如請求項28之用途,其中與CD40及FAP之至少一者相關之疾病為癌症。
- 一種用以檢測或測定CD40及FAP之至少一者之試劑,其包含如請求項1至17中任一項之雙專一性抗體或如請求項18之雙專一性抗體片段。
- 一種雙專一性抗體或該雙專一性抗體片段之衍生物,其係使放射性同位素、低分子藥劑、高分子藥劑、蛋白質或抗體醫藥以化學或基因工程方式與如請求項1至17中任一項之雙專一性抗體或如請求項18之雙專一性抗體片段結合而成。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2019092298 | 2019-05-15 | ||
| JP2019-092298 | 2019-05-15 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| TW202108626A true TW202108626A (zh) | 2021-03-01 |
Family
ID=73289814
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| TW109116232A TW202108626A (zh) | 2019-05-15 | 2020-05-15 | 與cd40及fap結合之雙專一性抗體 |
Country Status (9)
| Country | Link |
|---|---|
| US (1) | US20220259328A1 (zh) |
| EP (1) | EP3970743A4 (zh) |
| JP (1) | JPWO2020230899A1 (zh) |
| KR (1) | KR20220008820A (zh) |
| CN (1) | CN113891728A (zh) |
| AU (1) | AU2020276931A1 (zh) |
| CA (1) | CA3140385A1 (zh) |
| TW (1) | TW202108626A (zh) |
| WO (1) | WO2020230899A1 (zh) |
Families Citing this family (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| UY38995A (es) | 2019-12-20 | 2021-06-30 | Amgen Inc | Constructos de anticuerpo multiespecíficos agonistas de cd40 dirigidos a mesotelina para el tratamiento de tumores sólidos |
| AU2021249888A1 (en) * | 2020-04-01 | 2022-10-27 | Kyowa Kirin Co., Ltd. | Antibody composition |
| CN117616049A (zh) * | 2021-08-24 | 2024-02-27 | 江苏恒瑞医药股份有限公司 | Fap/cd40结合分子及其医药用途 |
| CR20240246A (es) * | 2021-12-20 | 2024-07-19 | Hoffmann La Roche | Anticuerpos agonistas anti-ltbr y anticuerpos biespecificos que los comprenden |
| EP4671277A1 (en) * | 2023-02-24 | 2025-12-31 | Shanghai Mabgen Biotech Ltd. | Pharmaceutical Composition of the FAP/CD40 Binding Molecule and its Pharmaceutical Use |
| WO2025005966A1 (en) * | 2023-06-29 | 2025-01-02 | Edge Animal Health, Inc. | Urine pcr assay and use thereof to diagnose and stage feline chronic kidney disease |
| WO2025096843A1 (en) * | 2023-11-03 | 2025-05-08 | Amgen Inc. | Bispecific molecules |
| WO2025113643A1 (en) | 2023-12-01 | 2025-06-05 | Gilead Sciences Inc. | Anti-fap-light fusion protein and use thereof |
| WO2025255405A1 (en) * | 2024-06-06 | 2025-12-11 | Bristol-Myers Squibb Company | Anti-fap antibodies and uses thereof |
Family Cites Families (39)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPS5623587A (en) | 1979-08-03 | 1981-03-05 | Mitsuwa Seiki Co Ltd | Vane type compressor |
| JPS58110600A (ja) | 1981-12-25 | 1983-07-01 | Kyowa Hakko Kogyo Co Ltd | ヒトβ型インタ−フエロン遺伝子を含む組みかえ体プラスミド |
| WO1985003934A1 (fr) | 1984-03-06 | 1985-09-12 | Takeda Chemical Industries, Ltd. | Proteine modifiee chimiquement et son procede de preparation |
| JPS60221091A (ja) | 1983-12-21 | 1985-11-05 | Kyowa Hakko Kogyo Co Ltd | 新規プロモ−タ− |
| US5168062A (en) | 1985-01-30 | 1992-12-01 | University Of Iowa Research Foundation | Transfer vectors and microorganisms containing human cytomegalovirus immediate-early promoter-regulatory DNA sequence |
| JP2564268B2 (ja) | 1985-08-28 | 1996-12-18 | 協和醗酵工業株式会社 | 融合抗原ポリペプチド |
| ZA872705B (en) | 1986-04-22 | 1987-10-05 | Immunex Corporation | Human g-csf protein expression |
| JP2958019B2 (ja) | 1988-05-06 | 1999-10-06 | 住友製薬株式会社 | ポリエチレングリコール誘導体、修飾ペプチドおよびその製造方法 |
| JP2928287B2 (ja) | 1988-09-29 | 1999-08-03 | 協和醗酵工業株式会社 | 新規ポリペプチド |
| JPH02257891A (ja) | 1989-03-31 | 1990-10-18 | Kyowa Hakko Kogyo Co Ltd | 組換え動物細胞による蛋白質の製造 |
| JPH0322979A (ja) | 1989-06-19 | 1991-01-31 | Kyowa Hakko Kogyo Co Ltd | 新規プラスミノーゲン活性化因子 |
| JP3131322B2 (ja) | 1991-12-17 | 2001-01-31 | 協和醗酵工業株式会社 | 新規α2→3シアリルトランスフェラーゼ |
| WO1994023021A1 (fr) | 1993-03-29 | 1994-10-13 | Kyowa Hakko Kogyo Co., Ltd. | α-1,3-FUCOSYLTRANSFERASE |
| WO1997010354A1 (fr) | 1995-09-11 | 1997-03-20 | Kyowa Hakko Kogyo Co., Ltd. | Anticorps de la chaine alpha du recepteur de l'interleukine 5 humaine |
| US6001358A (en) | 1995-11-07 | 1999-12-14 | Idec Pharmaceuticals Corporation | Humanized antibodies to human gp39, compositions containing thereof |
| IL132560A0 (en) | 1997-05-02 | 2001-03-19 | Genentech Inc | A method for making multispecific antibodies having heteromultimeric and common components |
| US6528624B1 (en) | 1998-04-02 | 2003-03-04 | Genentech, Inc. | Polypeptide variants |
| WO1999061057A2 (en) | 1998-05-23 | 1999-12-02 | Tanox, Inc. | Molecules targeting cd40 and tumor cells |
| EP1141024B1 (en) | 1999-01-15 | 2018-08-08 | Genentech, Inc. | POLYPEPTIDE COMPRISING A VARIANT HUMAN IgG1 Fc REGION |
| US6737056B1 (en) | 1999-01-15 | 2004-05-18 | Genentech, Inc. | Polypeptide variants with altered effector function |
| ES2601882T5 (es) | 1999-04-09 | 2021-06-07 | Kyowa Kirin Co Ltd | Procedimiento para controlar la actividad de una molécula inmunofuncional |
| DE19934785C2 (de) | 1999-07-27 | 2001-05-23 | Dorma Gmbh & Co Kg | Türterminal mit Montageplatte |
| KR20080074231A (ko) | 2000-06-29 | 2008-08-12 | 아보트 러보러터리즈 | 이중 특이성 항체 및 이의 제조 및 사용 방법 |
| HU231090B1 (hu) | 2000-10-06 | 2020-07-28 | Kyowa Kirin Co., Ltd. | Antitest-kompozíciót termelő sejt |
| EP1369431A4 (en) | 2001-10-15 | 2005-01-12 | Kirin Brewery | ANTI-HLA-DR ANTIBODIES |
| AR039067A1 (es) | 2001-11-09 | 2005-02-09 | Pfizer Prod Inc | Anticuerpos para cd40 |
| US7317091B2 (en) | 2002-03-01 | 2008-01-08 | Xencor, Inc. | Optimized Fc variants |
| WO2005035586A1 (ja) | 2003-10-08 | 2005-04-21 | Kyowa Hakko Kogyo Co., Ltd. | 融合蛋白質組成物 |
| EP1810979B1 (en) | 2004-09-22 | 2012-06-20 | Kyowa Hakko Kirin Co., Ltd. | STABILIZED HUMAN IgG4 ANTIBODIES |
| US7923538B2 (en) | 2005-07-22 | 2011-04-12 | Kyowa Hakko Kirin Co., Ltd | Recombinant antibody composition |
| ES2828627T3 (es) | 2008-04-25 | 2021-05-27 | Kyowa Kirin Co Ltd | Anticuerpo multivalente estable |
| RU2563514C2 (ru) | 2009-06-11 | 2015-09-20 | Интер-Юниверсити Рисерч Инститьют Корпорейшн Рисерч Организейшн Оф Информейшн Энд Системз | Способ получения белка |
| MY194892A (en) | 2014-04-07 | 2022-12-22 | Chugai Pharmaceutical Co Ltd | Immunoactivating antigen-binding molecule |
| BR112018074453A2 (pt) | 2016-05-27 | 2019-03-19 | Abbvie Biotherapeutics Inc. | proteínas de ligação biespecíficas ligando uma protéina imunomoduladora e um antígeno tumoral |
| US20190336615A1 (en) | 2017-01-27 | 2019-11-07 | Silverback Therapeutics, Inc. | Tumor targeting conjugates and methods of use thereof |
| EP3606956B1 (en) * | 2017-04-04 | 2024-07-31 | F. Hoffmann-La Roche AG | Novel bispecific antigen binding molecules capable of specific binding to cd40 and to fap |
| MA50505A (fr) * | 2017-11-01 | 2020-09-09 | Hoffmann La Roche | Anticorps 2 + 1 bispécifiques (contorsbodies) |
| CN111344307B (zh) * | 2017-11-08 | 2025-01-14 | 协和麒麟株式会社 | 与CD40和EpCAM结合的双特异性抗体 |
| JP6633598B2 (ja) | 2017-11-15 | 2020-01-22 | ファナック株式会社 | ステータ、ステータのブロックおよび回転電機 |
-
2020
- 2020-05-15 TW TW109116232A patent/TW202108626A/zh unknown
- 2020-05-15 AU AU2020276931A patent/AU2020276931A1/en not_active Abandoned
- 2020-05-15 US US17/611,361 patent/US20220259328A1/en not_active Abandoned
- 2020-05-15 KR KR1020217037019A patent/KR20220008820A/ko not_active Withdrawn
- 2020-05-15 CN CN202080035990.6A patent/CN113891728A/zh active Pending
- 2020-05-15 CA CA3140385A patent/CA3140385A1/en active Pending
- 2020-05-15 WO PCT/JP2020/019545 patent/WO2020230899A1/ja not_active Ceased
- 2020-05-15 JP JP2021519509A patent/JPWO2020230899A1/ja active Pending
- 2020-05-15 EP EP20805264.7A patent/EP3970743A4/en not_active Withdrawn
Also Published As
| Publication number | Publication date |
|---|---|
| CA3140385A1 (en) | 2020-11-19 |
| KR20220008820A (ko) | 2022-01-21 |
| EP3970743A1 (en) | 2022-03-23 |
| WO2020230899A1 (ja) | 2020-11-19 |
| CN113891728A (zh) | 2022-01-04 |
| JPWO2020230899A1 (zh) | 2020-11-19 |
| EP3970743A4 (en) | 2023-02-15 |
| AU2020276931A1 (en) | 2021-12-16 |
| US20220259328A1 (en) | 2022-08-18 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR101846590B1 (ko) | 항 tim-3 항체 | |
| TWI811260B (zh) | 與CD40及EpCAM結合之雙專一性抗體 | |
| TW202108626A (zh) | 與cd40及fap結合之雙專一性抗體 | |
| US20250250352A1 (en) | BISPECIFIC ANTIBODY BINDING TO TfR | |
| EP3243837A1 (en) | Bispecific antibody binding to trailr2 and psma | |
| TWI707868B (zh) | 抗人類Gas6單株抗體 | |
| TWI875760B (zh) | 與cd40及gpc3結合之雙專一性抗體 | |
| TW202140561A (zh) | 與cd3結合之雙特異性抗體 | |
| TW202317637A (zh) | 與cd116及cd131結合之雙特異性抗體 | |
| WO2021261597A1 (ja) | GPC3とTfRに結合するバイスペシフィック抗体 | |
| HK40062464A (zh) | 与tfr结合的双特异性抗体 | |
| HK40038050A (zh) | 与cd40和epcam结合的双特异性抗体 |