CN110192541B - 表达人源化主要组织相容性复合物的小鼠 - Google Patents
表达人源化主要组织相容性复合物的小鼠 Download PDFInfo
- Publication number
- CN110192541B CN110192541B CN201910379017.5A CN201910379017A CN110192541B CN 110192541 B CN110192541 B CN 110192541B CN 201910379017 A CN201910379017 A CN 201910379017A CN 110192541 B CN110192541 B CN 110192541B
- Authority
- CN
- China
- Prior art keywords
- mhc
- human
- mouse
- polypeptide
- rodent
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
Images












Classifications
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K67/00—Rearing or breeding animals, not otherwise provided for; New or modified breeds of animals
- A01K67/027—New or modified breeds of vertebrates
- A01K67/0275—Genetically modified vertebrates, e.g. transgenic
- A01K67/0278—Knock-in vertebrates, e.g. humanised vertebrates
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70503—Immunoglobulin superfamily
- C07K14/70539—MHC-molecules, e.g. HLA-molecules
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2207/00—Modified animals
- A01K2207/15—Humanized animals
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2217/00—Genetically modified animals
- A01K2217/07—Animals genetically altered by homologous recombination
- A01K2217/072—Animals genetically altered by homologous recombination maintaining or altering function, i.e. knock in
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2267/00—Animals characterised by purpose
- A01K2267/03—Animal model, e.g. for test or diseases
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70503—Immunoglobulin superfamily
- C07K14/70514—CD4
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/705—Receptors; Cell surface antigens; Cell surface determinants
- C07K14/70503—Immunoglobulin superfamily
- C07K14/70517—CD8
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/8509—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells for producing genetically modified animals, e.g. transgenic
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Immunology (AREA)
- General Health & Medical Sciences (AREA)
- Environmental Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Molecular Biology (AREA)
- Biotechnology (AREA)
- Medicinal Chemistry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Gastroenterology & Hepatology (AREA)
- Toxicology (AREA)
- Cell Biology (AREA)
- Veterinary Medicine (AREA)
- Biodiversity & Conservation Biology (AREA)
- Animal Husbandry (AREA)
- Animal Behavior & Ethology (AREA)
- Peptides Or Proteins (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Biomedical Technology (AREA)
- General Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Physics & Mathematics (AREA)
- Plant Pathology (AREA)
- Microbiology (AREA)
- Medicinal Preparation (AREA)
Abstract
本发明提供了表达人源/非人源嵌合MHC I和MHC II多肽和/或人源或人源化β2微球蛋白多肽的基因修饰的非人动物,以及包含该多肽的胚胎、细胞和组织。本发明还提供了制备所述基因修饰的动物的构建体以及制备该构建体的方法。本发明提供了使用所述基因修饰动物对人免疫系统的不同方面进行研究的方法。
Description
本申请是申请号为201480022254.1、申请日为2014年2月20日、发明名称为“表达人源化主要组织相容性复合物的小鼠”的中国发明专利申请的分案申请,原申请为国际申请号为PCT/US2014/017387的国家阶段申请,该国际申请要求申请日为2013年2月22日,申请号为61/767,811的美国临时专利申请的优先权。
相关申请的交叉引用
本申请根据35 U.S.C.§119(e)项要求申请日为2013年2月22日的美国临时专利申请号61/767,811的权益,其全部内容通过引用并入本申请。
序列表
本申请说明书引用2014年2月20日提交的名称为“2010794-0424_ST25”的ascii.txt文件电子形式的序列表。所述.txt文件于2014年2月20日生成且大小为8kb。
发明领域
本发明涉及表达人源或人源化主要组织相容性复合物(MHC)I类和人源或人源化MHC II类分子的基因修饰的非人动物,例如,啮齿动物(例如,小鼠或大鼠)。本发明还涉及表达人源或人源化MHC I蛋白(例如,MHC I α链)和人源或人源化MHC II蛋白(例如,MHC IIα和MHC II β链),并进一步表达人源或人源化β2微球蛋白的基因修饰的非人动物,例如,小鼠或大鼠;以及表达其的胚胎、组织和细胞。本发明进一步提供了用于制备同时表达人源或人源化MHC I类和II类蛋白,和/或β2微球蛋白的基因修饰的非人动物的方法。本发明还提供了一种在体外在人源化细胞免疫系统中或在基因修饰的非人动物中鉴定和评估肽的方法,以及修饰非人动物(例如,小鼠或大鼠)的MHC基因座以使其表达人源或人源化MHC I和人源或人源化MHC II蛋白的方法。
发明背景
在适应性免疫应答中,通过B淋巴细胞和T淋巴细胞上的受体分子(例如免疫球蛋白和T细胞受体或TCR)识别外来抗原。这些外来抗原被专门的蛋白以肽片段形式提呈在细胞表面上,这些专门的蛋白通常称为主要组织相容性复合物(MHC)分子。MHC分子由多个基因座编码,已发现其为跨距约4Mb的相连的基因簇。在小鼠中,MHC基因位于17号染色体上,并且由于历史原因将其称为组织相容性2(H-2)基因。在人中,该基因位于6号染色体上,将其称为人白细胞抗原(HLA)基因。在小鼠和人中的基因座是多基因的;这些基因座中包括三种高多态性的MHC基因类型(I、II和III类),其在人和小鼠基因组中具有相似的构造(分别参见图2和图3)。
MHC基因座在基因组中显示出最高的多态性;一些基因由>300个等位基因表示(例如人HLA-DRβ和人HLA-B)。所有I类和II类MHC基因均能提呈肽片段,但是各基因表达具有不同结合特性的蛋白,这反应了多态性和等位基因变体。任何给定个体均具有在免疫应答过程中能够提呈至B和T细胞表面的独特的肽片段范围。
人与小鼠均具有I类MHC基因(参见图2和图3)。在人中,经典的I类基因被称为HLA-A、HLA-B和HLA-C,而在小鼠中将其称为H-2K、H-2D和H-2L。I类分子由两条链组成:多态性α-链(有时被称为重链)和称为β2-微球蛋白的较小链(还被称为轻链),后者通常不具有多态性(图1,左)。这两条链在细胞表面形成非共价异二聚体。所述α-链包含三个结构域(α1、α2和α3)。α链基因的外显子1编码先导序列,外显子2和3编码α1和α2结构域,外显子4编码α3结构域,外显子5编码跨膜域,而外显子6和7编码胞质尾。α-链形成含有α1和α2结构域的肽结合缝隙(类似于Ig-样结构域)接着为类似于β2-微球蛋白的α3结构域。
β2微球蛋白是一种非糖基化的12kDa的蛋白;其功能之一在于稳定MHC I类α-链。与α-链不同,β2微球蛋白不跨膜。人β2微球蛋白的基因座在15号染色体上,而小鼠基因座在2号染色体上。β2微球蛋白的基因由4个外显子和3个内含子组成。循环形式的β2微球蛋白存在于血清、尿液和其他体液中;因此,非共价MHC I结合的β2微球蛋白在生理条件下能够与循环β2微球蛋白进行交换。
I类MHC分子在所有有核细胞中表达,包括肿瘤细胞。其特异性地在T和B淋巴细胞、巨噬细胞、树突状细胞和中性粒细胞以及其他细胞上表达并且其功能为在上述细胞表面将肽片段(长度通常为8-10个氨基酸)递呈给CD8+细胞毒性T淋巴细胞(CTL)。CTL特异性地杀伤由其自身的膜结合TCR识别的携带MHC I结合肽的任意细胞。当细胞展示非正常存在的细胞蛋白来源(例如病毒、肿瘤或其他非自身来源)的肽时,这种肽被CTL所识别,CTL被激活并杀伤展示该肽的细胞。
人与小鼠还均具有II类MHC基因(参见图2和图3)。在人中,经典的MHC II基因被称为HLA-DP、HLA-DQ和HLA-DR,而在小鼠中将其称为H-2A和H-2E(通常分别缩写为I-A和I-E)。由MHC II基因座中的基因编码的其他蛋白,在人中为HLA-DM和HLA-DO,以及在小鼠中为H-2M和H-2O,其不存在于细胞表面,而是位于内吞小泡,并确保MHCII分子与肽的正确装载。II类分子由两条肽链组成:α链和β链。α链的胞外部分含有两个胞外域α1和α2;并且β链的胞外部分也含有两个胞外域β1和β2(参见图1,右)。α链和β链彼此之间非共价结合。
MHC II类分子在抗原提呈细胞(APC),例如B细胞、巨噬细胞、树突状细胞、处于炎症过程中的内皮细胞等上表达。在APC表面上表达的MHCII分子通常将在细胞内的囊泡中产生的抗原提呈给针对CD4+T细胞。为了参与与CD4+T细胞的结合,MHC II类与所关注抗原的复合物必须非常稳定的存在足够长时间以便与CD4+T细胞结合。当CD4+T辅助细胞与APC表面上的外源性肽/MHC II复合物结合时,T细胞被活化以释放辅助针对侵入物的免疫应答的细胞因子。
由于存在耐受机制,因而不是所有的抗原均将激发T细胞活化。然而,在一些疾病(例如癌症、自身免疫性疾病)中来源于自身蛋白的肽成为免疫系统细胞成分的靶标,结果导致提呈这种肽的细胞受到破坏。在识别具有临床意义的抗原方面已经取得了重要进步(例如与不同类型的癌症相关的抗原)。然而,为了改进对在人T细胞中激发适宜应答的肽的鉴定和选择,特别是对具有临床意义的抗原肽,仍需要模拟人免疫系统各方面的体内和体外系统。因此,需要能够展示人免疫系统组分的生物系统(例如,基因修饰的非人动物和细胞)。
发明概述
本申请提供了一种用于产生或鉴定与人MHC I类蛋白及其嵌合体相联并与CD8+T细胞结合的肽以及与人MHC II类蛋白及其嵌合体相联并与CD4+T细胞结合的肽的生物系统。本申请提供了非人动物,其包含表达在细胞免疫应答中具有功能的人源化分子的非人细胞。本申请还提供了编码人源化MHC I和MHC II蛋白的人源化啮齿动物基因座。本申请还提供了表达人源化MHC分子的人源化啮齿动物细胞。本申请提供了包含人源化啮齿动物细胞的体内和体外系统,其中所述啮齿动物细胞表达一种或多种人源化免疫系统分子。
在不同实施方式中,本申请提供了一种非人动物,其在内源性MHC基因座包含:编码人源/非人源嵌合MHC I多肽的第一核苷酸序列,其中所述嵌合MHC I多肽的人源部分包含人MHC I多肽的胞外域;编码人源/非人源嵌合MHC II α多肽的第二核苷酸序列,其中所述人源/非人源嵌合MHC II α多肽的人源部分包含人MHC II α多肽的胞外域;和编码人源/非人源嵌合MHC II β多肽的第三核苷酸序列,其中所述人源/非人源嵌合MHC II β多肽的人源部分包含人MHC II β多肽的胞外域,其中所述非人动物表达来自其内源性非人源MHC基因座的人源/非人源嵌合MHC I和MHC II蛋白。在一个实施方式中,所述动物不表达来自所述内源性非人源MHC基因座的功能性内源性MHC I、II α和/或II β多肽。
在一个方面,所述第一核苷酸序列位于所述内源性非人源MHC I基因座,所述第二核苷酸序列位于所述内源性非人源MHC II α基因座,并且所述第三核苷酸序列位于所述内源性非人源MHC II β基因座。在一个方面,所述第一、第二和/或第三核苷酸序列可操作地连接至内源性非人源调控元件。在一个方面,所述第一核苷酸序列可操作地连接至内源性非人源MHC I启动子和调控元件,所述第二核苷酸序列可操作地连接至内源性非人源MHCII α启动子和调控元件,并且所述第三核苷酸序列可操作地连接至内源性非人源MHC II β启动子和调控元件。
在一个实施方式中,所述嵌合MHC I多肽的人源部分包含所述人MHC I多肽的α1、α2和α3域。在一个方面,所述嵌合MHC I多肽的非人源部分包含内源性非人源MHC I多肽的跨膜和胞质域。所述人MHC I多肽可选自由以下构成的组:HLA-A、HLA-B和HLA-C。在一个实施方式中,所述人MHC I多肽为HLA-A2。在另一方面,所述人MHC I多肽为HLA-A3、HLA-B7、HLA-B27、HLA-Cw6、或在人类群体中表达的任意其它MHC I分子。在一个附加实施方式中,本发明的非人动物在其内源性非人源β2微球蛋白基因座另外包含编码人源或人源化β2微球蛋白多肽的核苷酸序列,其中所述动物表达所述人源或人源化β2微球蛋白多肽。
在一个实施方式中,所述人MHC II α胞外域包含人MHC II α1和α2域。在另一个实施方式中,所述人MHC II β胞外域包含人MHC II β1和β2域。在一个方面,所述人源/非人源嵌合MHC II α多肽的非人源部分包含内源性非人源MHC II α多肽的跨膜和胞质域。在一个方面,所述人源/非人源嵌合MHC II β多肽的非人源部分包含内源性非人源MHC II β多肽的跨膜和胞质域。在一个实施方式中,所述人/小鼠嵌合MHC II α和β多肽的人源部分来源于选自由以下构成的组的人HLA II类蛋白:HLA-DR、HLA-DQ和HLA-DP。在一个特定实施方式中,所述人源/非人源嵌合MHC II α和β多肽的人源部分来源于人HLA-DR蛋白。或者,所述人源/非人源嵌合MHC II α和β多肽的人源部分可来源于选自以下的人MHC II蛋白:HLA-DR2、HLA-DQ2.5、HLA-DQ8、或在人类群体中表达的任意其它MHC II分子。
在一些方面,所提供的动物包含两个拷贝的含有第一、第二和第三核苷酸序列的MHC基因座,而在其它方面,所提供的动物包含一个拷贝的含有第一、第二和第三核苷酸序列的MHC基因座。因此,所述动物针对含有编码人源/非人源嵌合MHC I、MHC II α和MHC IIβ多肽的MHC基因座可以是纯合的或杂合的。在本发明的一些实施方式中,包含编码本发明所述人源/非人源嵌合MHC I、MHC II α和MHC II β多肽的核苷酸序列的所述基因修饰的MHC基因座,是在非人动物的生殖细胞系中。
本申请还提供一种MHC基因座,其包含编码人源/非人源嵌合MHC I多肽的第一核苷酸序列,其中所述嵌合MHC I多肽的人源部分包含人MHC I多肽的胞外域;编码人源/非人源嵌合MHC II α多肽的第二核苷酸序列,其中所述人源/非人源嵌合MHC II α多肽的人源部分包含人MHC II α多肽的胞外域;和编码人源/非人源嵌合MHC II β多肽的第三核苷酸序列,其中所述人源/非人源嵌合MHC II β多肽的人源部分包含人MHC II β多肽的胞外域。在一些方面,所述嵌合MHC I、II α和II β多肽的非人源部分分别包含非人源MHC I、II α和II β的跨膜和胞质域。
在一个实施方式中,所述基因工程非人动物是啮齿动物。在一个实施方式中,所述啮齿动物是大鼠或小鼠。在一个实施方式中,所述啮齿动物是小鼠。因此,在一个方面,所述第一核苷酸序列编码人/小鼠嵌合MHC I多肽,并且所述嵌合MHC I多肽的小鼠部分来源于H-2K、H-2D或H-2L。在一个特定实施方式中,所述嵌合MHC I多肽的小鼠部分来源于H-2K。在一个方面,所述第二核苷酸序列编码人源/小鼠嵌合MHC II α多肽,所述第三核苷酸序列编码人源/小鼠嵌合MHC II β多肽,并且所述嵌合MHC II α和β多肽的小鼠部分来源于H-2E或H-2A。在一个特定实施方式中,所述嵌合MHC II多肽的小鼠部分来源于H-2E。
因此,本申请还提供一种基因工程小鼠,其在内源性MHC基因座包含:编码人源/小鼠嵌合MHC I多肽的第一核苷酸序列,其中所述嵌合MHC I多肽的人源部分包含人MHC I多肽的胞外域;编码人源/小鼠嵌合MHC II α多肽的第二核苷酸序列,其中所述人源/非人源嵌合MHC II α多肽的人源部分包含人MHC II α多肽的胞外域;和编码人源/小鼠嵌合MHCII β多肽的第三核苷酸序列,其中所述人源/非人源嵌合MHC II β多肽的人源部分包含人MHC II β多肽的胞外域;其中所述小鼠表达来自其内源性小鼠MHC基因座的人源/小鼠嵌合MHC I和MHC II蛋白。在一个特定实施方式中,所述第一核苷酸序列编码HLA-A2/H-2K嵌合多肽,所述第二核苷酸序列编码HLA-DR4/H-2E嵌合多肽的α链,且所述第三核苷酸序列编码HLA-DR4/H-2E嵌合多肽的β链,并且所述小鼠表达功能性HLA-A2/H-2K和HLA-DR4/H-2E蛋白。在一个附加实施方式中,所述小鼠在内源性β2微球蛋白基因座另外包含编码人源或人源化β2微球蛋白多肽的核苷酸序列。在一个实施方式中,所述小鼠不表达来自其内源性MHC基因座的功能性内源性MHC多肽。
本申请还提供的是用于生成本申请所述的基因修饰的非人动物(例如啮齿动物,例如小鼠或大鼠)的方法。因此,在一个方面,本发明提供了一种生成基因修饰的非人动物的方法,其包括:在内源性非人源MHC II基因座用编码人源/非人源嵌合MHC II复合物的核苷酸序列取代编码非人源MHC II复合物的核苷酸序列以生成第一非人动物;以及在内源性非人源MHC I基因座用编码人源/非人源嵌合MHC I多肽的核苷酸序列取代编码非人源MHCI多肽的核苷酸序列以生成第二非人动物。在一个方面,所述取代核苷酸序列的步骤包括在非人ES细胞中同源重组,并且所述第二非人动物的生成是通过在具有编码人源/非人源嵌合MHC II复合物的核苷酸序列的ES细胞中同源重组。所述嵌合MHC II复合物包含人源/非人源嵌合MHC II α和β多肽。
在一个替代实施方式中,本发明提供了一种生成基因修饰的非人动物的方法,其包括:在内源性非人源MHC I基因座用编码人源/非人源嵌合MHC I多肽的核苷酸序列取代编码非人源MHC I多肽的核苷酸序列以生成第一非人动物;以及在内源性非人源MHC II基因座用编码人源/非人源嵌合MHC II复合物的核苷酸序列取代编码非人源MHC II复合物的核苷酸序列以生成第二非人动物。在一个方面,所述取代核苷酸序列的步骤包括在非人ES细胞中同源重组,并且所述第二非人动物的生成是通过在具有编码人源/非人源嵌合MHC I多肽的核苷酸序列的ES细胞中同源重组。
本申请还提供了细胞,例如分离的抗原提呈细胞,所述细胞来源于本申请所述的非人动物(例如啮齿动物,例如小鼠或大鼠)。本申请还提供了来源于本申请所述的非人动物的组织和胚胎。
本申请所述的任何实施方式和方面均可以彼此结合使用,除非另有说明或从上下文是显而易见的。通过随后的详细描述其他实施方式对本领域技术人员而言将是显而易见的。以下的详细描述包括对本发明不同实施方式的示例性描述,其对所主张的发明不具有限制性。附图构成了本说明书的一部分,其与说明书一起仅用于说明书实施方式,而不是限制本发明。
附图简述
图1是在细胞表面表达的MHC I类(左侧图)和MHC II类(右侧图)分子的示意图。灰色的圆圈表示在肽结合裂隙中结合的肽。
图2是人HLA的相对基因组结构的示意图(未按照比例),显示了I、II和III类基因。
图3是小鼠MHC的相对基因组结构的示意图(未按照比例),显示了I、II和III类基因。
图4描述了包含人源化MHC I和MHC II基因的人源化MHC基因座的生成策略。在所示特别实施方式中,所生成小鼠的MHC基因座包含人HLA-A2和HLA-DR4序列(H2-K+/1666MHC-II+/1681)。在人源化各阶段引入的大型靶向载体在箭头右侧示出。
图5(A-D)是生成包含人源化I-Eβ和I-Eα(即分别为H-2Eβ/HLA-DRβ1*04和H-2Eα/HLA-DRα*01嵌合体)的靶向载体的策略的示意图(未按照比例)。在图5C中,将来自图5B的最终人源化的MHC II序列连接至图5A的最终构建体的PI-SceI和I-CeuI限制性位点之间以产生包含人源化MHC II和BALB/c1-Eα的外显子1的构建体。Pg=假基因;BHR=细菌同源重组;CM=氯霉素;pec=大观霉素;hyg=潮霉素;neo=新霉素;EP=电穿孔。三角形表示外显子,实心的三角形表示来自C57BL/6小鼠的小鼠外显子(除了填充散点的三角形以外,其表示来自BALB/c小鼠I-Eα的外显子1),空心的三角形表示人外显子。
图6显示了使用潮霉素表达盒敲除小鼠基因座,随后引入包含人源化I-E β和I-Eα(即分别为H-2Eβ/HLA-DRβ1*04和H-2Eα/HLA-DRα*01嵌合体)的载体的MHC II类I-E和I-A基因的示意图,未按照比例。空心的三角形表示人外显子;实心的三角形表示小鼠外显子。圆圈内为用于基因分型的探针。
图7显示了Cre介导除去图6中的新霉素表达盒的示意图,未按照比例。空心的三角形表示人外显子;实心的三角形表示小鼠外显子。上两条链表示在人源化MHC II杂合子小鼠中的携带新霉素选择性表达盒的MHC II基因座,下两条链表示在人源化MHC II杂合子小鼠中的除去新霉素表达盒的MHC II基因座。
图8是用于制备表达人HLA-A2蛋白胞外区域的嵌合H-2K基因座的靶向策略的示意图(未按照比例)。小鼠序列以黑色表示,人序列以白色表示。L=先导序列,UTR=非翻译区,TM=跨膜域,CYT=胞质域,HYG=潮霉素。
图9是携带HLA-A2/H-2K和HLA-DR4/H-2E嵌合基因座的杂合子小鼠中HLA-A2和HLA-DR4体内表达的点图。稳态HLA-DR4表达虽低但存在;此低表达是预期的且所述表达在活化时上调。
发明实施方式详述
定义
本发明提供了同时表达人源或人源化MHC I和MHC II蛋白的基因修饰的非人动物(例如小鼠、大鼠、家兔等);包含其的胚胎、细胞和组织;制备其的方法;以及使用其的方法。除非另有定义,否则本申请使用的所有术语和短语包括所述术语和短语在本领域中所具有的含义,除非有相反的明确说明或根据使用所述术语或短语的上下文是显而易见的。
当用于描述保守的氨基酸取代时,术语“保守的”包括氨基酸残基被另一个具有带有相似化学性质(例如电荷或疏水性)的侧链R基的氨基酸残基取代。可以通过修饰核苷酸序列以引入将编码保守取代的核苷酸改变从而实现保守的氨基酸取代。在通常情况下,保守的氨基酸取代将基本上不会改变所关注蛋白的功能性性质,例如MHC I或MHC II提呈所关注肽的能力。具有带有相似化学性质的侧链的氨基酸基团的实例包括脂肪族侧链如甘氨酸、丙氨酸、缬氨酸、亮氨酸和异亮氨酸;脂肪族-羟基侧链如丝氨酸和苏氨酸;含有酰胺的侧链如天冬酰胺和谷胺酰胺;芳香侧链如苯丙氨酸、酪氨酸和色氨酸;碱性侧链如赖氨酸、精氨酸和组氨酸;酸性侧链如天冬氨酸和谷氨酸;和含硫侧链如半胱氨酸和甲硫氨酸。保守的氨基酸取代基团包括例如缬氨酸/亮氨酸/异亮氨酸、苯丙氨酸/酪氨酸、赖氨酸/精氨酸、丙氨酸/缬氨酸、谷氨酸/天冬氨酸和天冬酰胺/谷氨酰胺。在一些实施方式中,保守的氨基酸取代可以是蛋白中的任意天然残基被丙氨酸取代,如用于例如丙氨酸扫描诱变中。在一些实施方式中,如Gonnet等((1992)Exhaustive Matching of the Entire ProteinSequence Database,Science 256:1443-45)所公开的使得在PAM250 log-似然性矩阵中具有正值的保守取代,其通过引用并入本申请。在一些实施方式中,所述取代是适度保守的取代,其中所述取代在PAM250 log-似然性矩阵中具有非负值。
因此,本发明还包括一种基因修饰的非人动物,所述动物的基因组包含编码人源或人源化MHC I和II多肽的核苷酸序列,其中所述MHC I或MHC II多肽包含本申请所述的氨基酸序列的保守的氨基酸取代。
本领域技术人员将理解,除了本申请所述的编码人源或人源化MHC I或MHC II多肽的核酸残基以外,由于遗传密码的简并性,其他核酸也可以编码本发明的多肽。因此,除了在其基因组中包含编码具有保守的氨基酸取代的MHC I和MHC II多肽的核苷酸序列的基因修饰的非人动物以外,本申请还提供了由于遗传密码的简并性使得在其基因组包含不同于本申请所述的核苷酸序列的非人动物。
术语“同一性”当与序列一起使用时包括由本领域已知的能够用于测定核苷酸和/或氨基酸序列同一性的多种不同算法确定的同一性。在本申请所述的一些实施方式中,使用开放缺口罚分为10.0、延伸缺口罚分为0.1的ClustalW v.1.83(slow)算法和使用Gonnet相似性矩阵(MacVectorTM10.0.2,MacVector Inc.,2008)确定同一性。用于序列同一性比较的序列的长度将取决于特定的序列。在不同实施方式中,通过从其N末端至C末端对成熟蛋白的序列进行比较确定同一性。在不同实施方式中,当将人源/非人源嵌合序列与人序列进行比较时,将所述人源/非人源嵌合序列的人源部分(而不是非人源部分)用于比较,以确定人序列和人源/非人源嵌合序列的人源部分之间的同一性水平(例如对人/小鼠嵌合蛋白的人源胞外域与人蛋白的人胞外域进行比较)。
术语“同源性”或“同源的”涉及序列例如核苷酸或氨基酸序列时指两条序列经最佳比对和比较具有至少约75%的核苷酸或氨基酸、例如至少约80%的核苷酸或氨基酸、例如至少约90-95%的核苷酸或氨基酸,例如超过97%的核苷酸或氨基酸是相同的。本领域技术人员将理解,对于最佳基因靶向而言,靶向构建体应含有与内源性DNA序列同源的臂(即“同源臂”);因此,能够在靶向构建体与被靶向的内源性序列之间发生同源重组。
术语“可操作地连接”指并置的行为,其中被这样描述的组分处于允许其以预计方式发挥功能的关系中。因此,编码蛋白的核酸序列可以可操作地与调控序列(例如启动子、增强子、沉默子序列等)连接,以保持适宜的转录调控。此外,在细胞中本发明的嵌合或人源化蛋白的不同部分可以可操作地连接,从而保留折叠、加工、靶向、表达和所述蛋白在细胞中的其他功能性质。除非另有明示,否则本发明的嵌合或人源化蛋白的不同结构域彼此之间可操作地连接。
本申请所使用的术语“MHC I复合物”或类似表述包括MHC I α链多肽和β2-微球蛋白多肽之间的复合物。本申请所使用的术语“MHC I多肽”或类似表述包括单独的MHC I α链多肽。本申请所使用的术语“MHC II复合物”、“MHC II蛋白”或类似表述包括MHC II α多肽和MHC II β多肽之间的复合物。本申请所使用的术语“MHC II α多肽”或“MHC II β多肽”(或类似表述)分别包括单独的MHC II α多肽或单独的MHC II β多肽。类似地,术语“HLA-DR4复合物”、“HLA-DR4蛋白”、“H-2E复合物”、“H-2E蛋白”或类似表述指α和β多肽之间的复合物。通常,术语“人MHC”和“HLA”可以互换使用。
术语“取代”当涉及基因取代时,指在内源性基因座加入外源性遗传物质,以使用直系同源或同源核酸序列取代全部或部分内源性基因。如下述实例所示,内源性MHC基因座的核酸序列被包含编码人MHC I多肽的一部分(特别地,编码MHC I多肽的胞外部分)以及人MHC II α和β多肽的部分(特别地,编码MHC II α和β多肽的胞外部分)的序列的核苷酸序列取代。
本申请所使用的“功能性”,例如当涉及功能性多肽时,指保留至少一种与天然蛋白相关的正常生物活性的多肽。例如,在本发明的一些实施方式中,在内源性基因座的取代(例如在内源性非人源MHC基因座的取代)使得基因座无法表达功能性内源性多肽,例如,MHC I或MHC II多肽。同样地,本申请所使用的术语“功能性”当涉及蛋白的功能性胞外域时是指保留其功能性的胞外域,例如,就MHC I或MHC II而言,是指其结合抗原的能力、结合T细胞辅助受体的能力等。在本发明的一些实施方式中,在内源性MHC基因座的取代使得基因座无法表达内源性MHC的胞外域(例如,功能性胞外域),而表达人MHC的胞外域(例如,功能性胞外域)。
基因修饰的MHC动物
在不同实施方式中,本发明总的提供了基因修饰的非人动物,所述动物在其基因组中包含编码人源或人源化MHC I和MHC II多肽的核苷酸序列;因此,所述动物表达人源或人源化MHC I和MHC II多肽。
MHC基因分为三类:I类、II类和III类,其均在人6号染色体或小鼠17号染色体上编码。人和小鼠MHC类型相对结构的示意图分别见图2和图3。MHC基因为小鼠和人基因组中最具多态性的基因。MHC多态性被推定为在提供进化优势方面是重要的;序列的改变能够导致肽结合的差异,以便更好地将病原体提呈给细胞毒性T细胞。
MHC I类蛋白包含胞外域(其包含三个结构域:α1、α2和α3)、跨膜域和胞质尾。α1和α2结构域形成肽结合裂隙,而α3与β2-微球蛋白相互作用。
除了与β2-微球蛋白的相互作用之外,α3结构域还与TCR辅助受体CD8之间相互作用,以促进抗原特异性活化。尽管MHC I类与CD8的结合比TCR与MHC I类的结合弱约100倍,但是CD8结合增强了TCR结合的亲和性。Wooldridge等,(2010)MHC Class I Moleculeswith Superenhanced CD8 Binding Properties Bypass the Requirement for CognateTCR Recognition and Nonspecifically Activate CTLs,J.Immunol.184:3357-3366。有趣的是,增强MHC I类与CD8的结合消除了CTL活化中的抗原特异性。同上。
CD8与MHC I类分子的结合具有种属特异性;已发现CD8的小鼠同源物Lyt-2在α3结构域与H-2Dd分子结合,但其不与HLA-A分子结合。Connolly等,(1988)The Lyt-2 MoleculeRecognizes Residues in the Class I 3 Domain in Allogeneic Cytotoxic T CellResponses,J.Exp.Med.168:325-341。这种差异性结合可能是由CD8上的CDR样决定簇(CDR1-和CDR2-样)在人和小鼠之间不保守所导致的。Sanders等,(1991) Mutations inCD8 that Affect Interactions with HLA Class I and Monoclonal Anti-CD8Antibodies,J.Exp.Med.174:371-379;Vitiello等,(1991)Analysis ofthe HLA-restricted Influenza-specific Cytotoxic T Lymphocyte Response in TransgenicMice Carrying a Chimeric Human-Mouse Class I Major HistocompatibilityComplex,J.Exp.Med.173:1007-1015;以及,Gao等,(1997)Crystal structure ofthecomplex between human CD8αα and HLA-A2,Nature 387:630-634。已报道CD8在α3结构域的保守区(位点223-229处)结合HLA-A2。在HLA-A中的单一取代(V245A)导致CD8与HLA-A的结合降低,同时伴随着T细胞介导的裂解大大减少。Salter等,(1989),Polymorphism inthe α3 domain of HLA-A molecules affects binding to CD8,Nature 338:345-348。在通常情况下,HLA-A分子在α3结构域的多态性也会影响其与CD8的结合。同上。在小鼠中,在H-2Dd中残基227处的氨基酸取代影响小鼠Lyt-2与H-2Dd的结合,并且使用突变的H-2Dd转染的细胞不会被CD8+T细胞裂解。Potter等,(1989)Substitution at residue 227 of H-2class I molecules abrogates recognition by CD8-dependent,but not CD8-independent,cytotoxic T lymphocytes,Nature 337:73-75。
因此,由于MHC I类α3结构域与CD8之间的相互作用具有种属特异性,因此包含H-2K α3结构域被人HLA-A2 α3结构域取代的MHC I复合物在缺乏人CD8的小鼠中(即在体内)无功能。在HLA-A2的转基因动物中,使用人α3结构域取代小鼠α3结构域使得T细胞应答恢复。Irwin等,(1989)Species-restricted interactions between CD8 and the α3domain of class I influence the magnitude of the xenogeneic response,J.Exp.Med.170:1091-1101;Vitiello等,(1991),见前。
小鼠MHC I类蛋白的跨膜域和胞质尾也具有重要功能。MHC I跨膜域的一个功能是促进HLA-A2同型细胞粘附的调控(增强或抑制粘附),这可能是表面MHC分子交联(或连接)的结果。Wagner等,(1994)Ligation ofMHC Class I and Class II Molecules Can Leadto Heterologous Desensitization of Signal Transduction Pathways That RegulateHomotypic Adhesion in Human Lymphocytes,J.Immunol.152:5275-5287。细胞粘附能够受到与HLA-A2分子不同表位结合的mAb的影响,这表明HLA-A2上的多个位点参与调节同型细胞的粘附;依据所结合表位的不同,该影响可能会增强或抑制HLA-A2依赖性粘附。同上。
据报道由MHC I基因的外显子6和7编码的胞质尾对细胞表面上的适宜表达和LIR1介导的NK细胞毒性的抑制是必要的。Gruda等,(2007)Intracellular Cysteine Residuesin the Tail of MHC Class I Proteins Are Crucial for Extracellular Recognitionby Leukocyte Ig-Like Receptor 1,J.Immunol.179:3655-3661。胞质尾对于至少一些通过在其半胱氨酸残基上形成二硫键的MHCI分子的多聚化是必需的,因此其可能在成簇和被NK细胞识别中具有重要作用。Lynch等,(2009)Novel MHC Class I Structures onExosomes,J.Immunol.183:1884-1891。
尽管HLA-A2的胞质域含有一个组成型的磷酸化丝氨酸残基和一个可磷酸化的酪氨酸,但是在Jurkat细胞中缺乏胞质域的突变的HLA-A2分子在表达、细胞骨架结合、聚合和内吞内化方面表现正常。Gur等,(1997)Structural Analysis of Class I MHCMolecules:The Cytoplasmic Domain Is Not Required for CytoskeletalAssociation,Aggregation,and Internalization,Mol.Immunol.34(2):125-132。缺乏胞质域的截短HLA-A2分子显示出正常的表达且与β2微球蛋白的结合,同上。
然而,一些研究已表明胞质尾在细胞内转运、树突状细胞(DC)介导的抗原递呈和CTL引发中具有重要作用。已证明外显子6编码的酪氨酸残基是MHC I通过内涵体小室的转运、外源性抗原的递呈和CTL引发所必需的;而外显子7的缺失导致抗病毒CTL应答的增强。Lizee等,(2003)Control of Dendritic Cross-Presentation by the MajorHistocompatibility Complex Class ICytoplasmic Domain,Nature Immunol.4:1065-73;Basha等,(2008)MHC Class I Endosomal and Lysosomal Trafficking Coincideswith Exogenous Antigen Loading in Dendritic Cells,PLoS ONE 3:e3247;以及Rodriguez-Cruz等,(2011)Natural Splice Variant of MHC Class I Cytoplasmic TailEnhances Dendritic Cell-Induced CD8+T-Cell Responses and Boosts Anti-TumorImmunity,PLoS ONE6:e22939。
MHC II类复合物包含两个非共价结合域:α链和β链,在本申请中也称为α多肽和β多肽(图1,右)。该蛋白横跨细胞质膜;因此其含有胞外域、跨膜域和胞质域。α链的胞外部分包括α1和α2结构域,β链的胞外部分包括β1和β2结构域。α1和β1域在细胞表面形成肽结合裂隙。由于MHC II复合物的肽结合裂隙的三维构象,理论上对所结合抗原的长度没有上限,但是通常MHC II提呈肽的长度为13至17个氨基酸。
除了其与抗原肽相互作用以外,在MHC II复合物形成和肽获取过程中MHCII分子的肽结合裂隙与不变链(Ii)相互作用。α/β MHC II二聚体在内质网组装并与Ii链结合,其负责控制肽结合并且将MHC II靶向至内吞通路。在内涵体中,Ii发生蛋白水解,Ii的小片段II类相关恒定链肽(CLIP)仍位于肽结合裂隙。在内涵体中,在HLA-DM的控制下(在人中),CLIP被交换为抗原肽。
在α2和β2域交界处的疏水性缝隙,MHC II与T细胞辅助受体CD4相互作用。Wang和Reinherz(2002)Structural Basis of T Cell Recognition of Peptides Bound to MHCMolecules,Molecular Immunology,38:1039-49。当CD4和T细胞受体跟相同的与肽复合的MHCII分子结合时,T细胞对抗原的敏感性增加,其活化所需的抗原降低100倍。参见Janeway’s Immunobiology,第7版,Murphy等编辑。Garland Science,2008,其通过引用并入本申请。
已经提出了MHC II跨膜和胞质域的很多功能。就质膜域而言,已发现其对细胞内信号传导、运输至细胞质膜和最终的抗原提呈均是重要的。例如,已发现T细胞杂交瘤对转染了胞质域截短的MHC II β链的抗原提呈细胞(APC)应答较弱,并且B细胞分化的诱导受阻。参见例如Smiley等,(1996)Truncation ofthe class II β-chain cytoplasmicdomain influences the level of class II/invariant chain-derived peptidecomplexes,Proc.Natl.Acad.Sci.USA,93:241-44。II类分子的截短似乎会削弱cAMP的产生。据推测MHC II胞质尾的缺失影响细胞内运输,从而阻止复合物在内吞途径中遇到相关抗原。Smiley等(同上)证明了II类分子在胞质域的截短减少了CLIP/II类复合物的数量,推测其影响了CLIP有效调节抗原提呈的能力。
已推测由于MHC II簇对T细胞受体(TCR)的触发具有重要作用,因而如果在胞质域截短的MHCII分子无法与细胞骨架结合并且因此而聚集,则向T细胞的抗原提呈将受到影响。Ostrand-Rosenberg等,(1991)Abrogation of Tumorigenicity by MHC Class IIAntigen Expression Requires the Cytoplasmic Domain of the Class II Molecule,J.Immunol.147:2419-22。事实上,最近发现在胞质域截短的HLA-DR在寡聚化后无法与细胞骨架结合。El Fakhy等,(2004)Delineation of the HLA-DR Region and the ResiduesInvolved in the Association with the Cytoskeleton,J.Biol.Chem.279:18472-80。重要的是,肌动蛋白细胞骨架是局部信号转导活性的位点,其能够影响抗原提呈。除了与细胞骨架结合以外,最近的研究还发现在全部HLA-DR分子中有高达20%组成性地存在于APC中的脂筏,其是富含胆固醇和鞘糖脂的微结构域,并且这种定位对抗原提呈、免疫突触的形成和MHC II介导的信号传导均具有重要作用。参见例如Dolan等,(2004)Invariant Chainand the MHC IICytoplasmic Domains Regulate Localization of MHC Class IIMolecules to Lipid Rafts in Tumor Cell-Based Vaccines,J.Immunol.172:907-14。Dolan等提出MHC II胞质域的截短减少了MHC II在脂筏的组成性定位。
此外,MHC II的结构域,特别是β链,含有受泛素化连接酶RING-CH I(MARCH I)泛素化的亮氨酸残基,RING-CH I(MARCH I)控制MHC II的胞内运输、内化和降解的膜结合;并且发现当树突状细胞成熟时MARCH-介导的泛素化停止,结果导致细胞质膜MHC II的水平升高。Shin等,(2006)Surface expression of MHC class II in dendritic cells iscontrolled by regulated ubiquitination,Nature 444:115-18;De Gassart等,(2008)MHC class II stabilization at the surface of human dendritic cells is theresult of maturation-dependent MARCH I down-regulation,Proc.Natl.Acad.Sci.USA105:3491-96。
MHC II的α和β链的跨膜域彼此之间相互作用并且该相互作用对II类MHC复合物的适宜组装具有重要作用。Cosson和Bonifacino (1992)Role of Transmembrane DomainInteractions in the Assembly of Class II MHC Molecules,Nature 258:659-62。事实上,α和β链的跨膜域被IL-2受体的α链取代的MHC II分子仍保留在ER中,并且其在细胞表面几乎检测不到,同上。通过诱变研究,发现在α和β跨膜域的保守Gly残基负责MHC II在细胞表面的组装,同上。因此,跨膜和胞质域对MHC II复合物的适当功能均是至关重要的。
在不同实施方式中,本申请提供的是一种基因修饰的非人动物,例如啮齿动物(例如,小鼠或大鼠),在其基因组包含编码人源或人源化MHC I多肽的核苷酸序列和编码人源或人源化MHC II蛋白的核苷酸序列。所述MHC I核苷酸序列可编码部分人源和部分非人源的MHC I多肽,例如,人源/非人源嵌合MHC I多肽,而所述MHC II核苷酸序列可编码部分人源和部分非人源的MHC II蛋白,例如,人源/非人源嵌合MHC II蛋白(例如,包含人源/非人源嵌合MHC II α和β多肽)。在一些方面,所述动物不表达内源性MHC I和II多肽,例如,功能性内源性MHC I和II多肽。
在美国专利申请号13/661,159和13/793,812中公开了基因修饰的非人动物,在其基因组中,例如在内源性基因座,包含编码人源/非人源嵌合MHC I多肽的核苷酸序列,所述申请通过引入整体并入本申请。在美国专利申请号13/661,116和13/793,935中公开了基因修饰的非人动物,在其基因组中,例如在内源性基因座,包含编码人源化(例如人源/非人源嵌合)MHC II多肽的核苷酸序列,所述申请通过引入整体并入本申请。本申请提供了一种基因修饰的非人动物,在其内源性MHC基因座包含编码人源/非人源嵌合MHC I多肽和人源/非人源嵌合MHC II多肽的核苷酸序列。由于小鼠17号染色体和人6号染色体上MHC I和MHC II基因的紧密连接,很难通过简单育种人源化MHC I和MHC II动物而生成此动物。因此,本申请还提供了用于生成包含编码人源或人源化MHC I和MHC II多肽的序列的基因修饰的非人动物(例如其中编码内源性MHC I和II多肽的序列被编码人源/非人源嵌合MHC I和II多肽的那些序列取代的动物)的新方法。
因此,在不同实施方式中本申请提供了一种基因修饰的非人动物,在其基因组中(例如在内源性MHC基因座)包含编码人源/非人源嵌合MHC I多肽的第一核苷酸序列,其中所述嵌合MHC I多肽的人源部分包含人MHC I多肽的胞外域;编码人源/非人源嵌合MHC IIα多肽的第二核苷酸序列,其中所述嵌合MHC II α多肽的人源部分包含人MHC II α多肽的胞外域;和编码人源/非人源嵌合MHC II β多肽的第三核苷酸序列,其中所述嵌合MHC II β多肽的人源部分包含人MHC II β多肽的胞外域;其中所述非人动物表达来自其内源性非人源MHC基因座的人源/非人源嵌合MHC I和MHC II蛋白。在一个实施方式中,所述第一、第二和/或第三核苷酸序列位于所述内源性非人源MHC基因座。在一个实施方式中,其中所述非人动物是小鼠,所述第一、第二和/或第三核苷酸序列位于小鼠17号染色体上的内源性小鼠MHC基因座。在一个实施方式中,所述第一核苷酸序列位于所述内源性非人源MHC I基因座。在一个实施方式中,所述第二核苷酸序列位于所述内源性非人源MHC II α基因座。在一个实施方式中,所述第三核苷酸序列位于所述内源性非人源MHC II β基因座。
在一个实施方式中,所述非人动物仅表达来自所述内源性非人源MHC基因座的所述人源/非人源嵌合MHC I、MHC II α和/或MHC II β多肽且不表达内源性非人源MHC多肽(例如,功能性内源性MHC I、II α和/或II β多肽)。在一个实施方式中,本申请所述所述动物在其细胞(例如抗原提呈细胞等)表面表达功能性嵌合MHC I和功能性嵌合MHC II。
在一个实施方式中,所述人源/非人源嵌合MHC I多肽在其人源部分中包含人MHCI多肽的肽结合域。在一个方面,所述嵌合多肽的人源部分包含人MHC I的胞外域。在此实施方式中,所述嵌合多肽的人源部分包含人MHC I α链的胞外域。在一个实施方式中,所述嵌合多肽的人源部分包含人MHC I的α1和α2域。在另一个实施方式中,所述嵌合多肽的人源部分包含人MHC I的α1、α2和α3域。
在一个方面,所述嵌合MHC II α多肽的人源部分和/或所述嵌合MHC II β多肽的人源部分分别包含人MHC II α多肽和/或人MHC II β多肽的肽结合域。在一个方面,所述嵌合MHC II α和/或β多肽的人源部分分别包含人MHC II α和/或β多肽的胞外域。在一个实施方式中,所述嵌合MHC II α多肽的人源部分包含人MHC II α多肽的α1域;在另一个实施方式中,所述嵌合MHC II α多肽的人源部分包含人MHC II α多肽的α1和α2域。在一个附加实施方式中,所述嵌合MHC II β多肽的人源部分包含人MHC II β多肽的β1域;在另一个实施方式中,所述嵌合MHC II β多肽的人源部分包含人MHC II β多肽的β1和β2域。
所述人源或人源化MHC I多肽可来源于由任一HLA-A、HLA-B、HLA-C、HLA-E、HLA-F或HLA-G基因座编码的功能性人HLA分子。所述人源或人源化MHC II多肽可来源于由任一HLA-DP、-DQ和-DR基因座编码的功能性人HLA分子。常用的HLA抗原和等位基因列表描述在Shankarkumar等.((2004)The Human Leukocyte Antigen(HLA)System,Int.J.Hum.Genet.4(2):91-103)中,其通过引用并入本申请。Shankarkumar等还提供了对本领域使用的HLA命名法的简要说明。有关HLA命名法和不同HLA等位基因的附加信息可参见Holdsworth等,(2009)The HLA dictionary 2008:a summary of HLA-A,-B,-C,-DRB1/3/4/5,and DQB1 alleles and their association with serologically defined HLA-A,-B,-C,-DR,and-DQ antigens,Tissue Antigens 73:95-170,以及由Marsh等最近更新的(2010)Nomenclature for factors of the HLA system,2010,Tissue Antigens 75:291-455,其均通过引用并入本申请。因此,所述人源或人源化MHC I和/或II多肽可来源于本申请所述的任意功能性人HLA分子。
特别感兴趣的是人HLA,已知与多种人类疾病(例如人自身免疫性疾病)相关的特定多态性HLA等位基因。事实上,已确认在HLA基因座中的特定多态性与类风湿性关节炎、I型糖尿病、桥本氏甲状腺炎、多发性硬化症、重症肌无力、格雷夫斯氏病、系统性红斑狼疮、乳糜泻、克隆氏病、溃疡性结肠炎及其他自身免疫性疾病的进展相关。参见,例如,Wong和Wen(2004)What can the HLA transgenic mouse tell us about autoimmunediabetes?,Diabetologia 47:1476-87;Taneja和David(1998)HLA Transgenic Mice asHumanized mouse Models of Disease and Immunity,J.Clin.Invest.101:921-26;Bakker等(2006),A high-resolution HLA and SNP haplotype map for diseaseassociation studies in the extended human MHC,Nature Genetics 38:1166-72及补充信息;以及International MHC and Autoimmunity Genetics Network(2009)Mappingof multiple susceptibility variants within the MHC region for 7 immune-mediated diseases,Proc.Natl.Acad.Sci.USA 106:18680-85。因此,所述人源或人源化MHC I和/或II多肽可来自于已知与特定疾病(自身免疫性疾病)相关的人HLA分子。
在一个特定方面,所述人源或人源化MHC I多肽来源于人HLA-A。在一个特定实施方式中,所述HLA-A多肽是一种HLA-A2多肽(例如,以及HLA-A2.1多肽)。在一个实施方式中,所述HLA-A多肽是一种由HLA-A*0201等位基因(例如,HLA-A*02:01:01:01等位基因)编码的多肽。所述HLA-A*0201等位基因通常用于北美人群中。尽管本实例描述了此特定的HLA序列;但是任意适宜的HLA-A序列均包含在本申请中,例如在人群中显示出的HLA-A2多态性变体,具有一个或多个保守性或非保守性氨基酸修饰的序列,由于遗传密码的简并性与本申请所述序列不同的核酸序列等。
在另一特定方面,所述嵌合MHC I多肽的人源部分来源于选自HLA-B和HLA-C的人MHC I。在一个方面,其来源于HLA-B,例如,HLA-B27。在另一个方面,其来源于HLA-A3、-B7、-Cw6等。
在一个特定方面,本申请所述的人源化MHC II α和β多肽的人源部分来源于人HLA-DR,例如HLA-DR4。典型地,HLA-DR的α链是单一形态的,例如HLA-DR复合物的α链由HLA-DRA基因(例如HLA-DRα*01基因)编码。另一方面,HLA-DR的β链是多态的。因此,HLA-DR4包含由HLA-DRA基因编码的α链和由HLA-DRB1基因(例如HLA-DRβ1*04基因)编码的β链。如本申请的下文所述,HLA-DR4已知与多种自身免疫性疾病的发生相关,例如类风湿性关节炎、I型糖尿病、多发性硬化症等。在本发明的一个实施方式中,HLA-DRA等位基因是HLA-DRα*01等位基因,例如HLA-DRα*01:01:01:01。在另一个实施方式中,HLA-DRB等位基因是HLA-DRβ1*04,例如HLA-DRβ1*04:01:01。尽管本实例描述了这些特定的HLA序列;但是任意适宜的HLA-DR序列均包含在本申请中,例如在人类群体中显示出的多态性变体、具有一个或多个保守性或非保守性氨基酸修饰的序列、由于密码子的简并性与本申请所述序列不同的核酸序列等。
所述嵌合MHC II α和/或β嵌合多肽的人源部分可以由已知与常见的人类疾病相关的HLA等位基因的核苷酸序列编码。这种HLA等位基因包括但不限于HLA-DRB1*0401、-DRB1*0301、-DQA1*0501、-DQB1*0201、-DRB1*1501、-DRB1*1502、-DQB1*0602、-DQA1*0102、-DQA1*0201、-DQB1*0202、-DQA1*0501及其组合。对于与HLA等位基因/疾病相关的概述,参见在上的Bakker等(2006),通过引用并入本申请。
在一个方面,所述人源/非人源嵌合MHC I、MHC II α和/或MHC II β多肽的非人源部分分别包含内源性非人源(例如啮齿动物,例如小鼠、大鼠等)MHC I、MHC II α和/或MHCII β多肽的跨膜和/或胞质域。因此,所述人源/非人源嵌合MHC I多肽的非人源部分可包含内源性非人源MHC I多肽的跨膜和/或胞质域。所述嵌合MHC II α多肽的非人源部分可包含内源性非人源MHC II α多肽的跨膜和/或胞质域。所述人源/非人源嵌合MHC II β多肽的非人源部分可包含内源性非人源MHC II β多肽的跨膜和/或胞质域。在一个方面,所述非人动物是小鼠,且所述嵌合MHC I多肽的非人源部分来源于小鼠H-2K蛋白。在一个方面,所述动物是小鼠,且所述嵌合MHC II α和β多肽的非人源部分来源于小鼠H-2E蛋白。因此,所述嵌合MHC I多肽的非人源部分可包含来源于小鼠H-2K的跨膜和胞质域,且所述嵌合MHC II α和β多肽的非人源部分可包含来源于小鼠H-2E蛋白的跨膜和胞质域。尽管在实例中包含了特定的H-2K和H-2E序列,但是任意适宜的序列(例如多态性变体、保守性/非保守性氨基酸取代等)均包含在本申请中。
人源/非人源嵌合多肽可以是这样的,其包含人源或非人源先导(信号)序列。在一个实施方式中,所述嵌合MHC I多肽包含内源性MHC I多肽的非人源先导序列。在一个实施方式中,所述嵌合MHC II α多肽包含内源性MHC II α多肽的非人源先导序列。在一个实施方式中,所述嵌合MHC II β多肽包含内源性MHC II β多肽的非人源先导序列。在一个替代实施方式中,所述嵌合MHC I、MHC II α和/或MHC II β多肽分别包含来自于另一种非人动物例如另一种啮齿动物或另一种小鼠品系的MHC I、MHC II α和/或MHC II β多肽的非人源先导序列。因此,编码所述嵌合MHC I、MHC II α和/或MHC II β多肽的核苷酸序列可以可操作地分别与编码非人源MHC I、MHC II α和/或MHC II β先导序列的核苷酸序列连接。在又另一个实施方式中,所述嵌合MHC I、MHC II α和/或MHC II β多肽分别包含人MHC I、人MHCII α和/或人MHC II β多肽的人先导序列(例如分别为人HLA-A2、人HLA-DRA和/或人HLA-DRβ1*04的先导序列)。
人源/或非人源嵌合MHC I、MHC II α和/或MHC II β多肽可以在其人源部分分别包含完整或基本完整的人MHC I、人MHC II α和/或人MHC II β多肽的胞外域。因此,人源部分可包含至少80%、优选至少85%、更优选至少90%例如95%或以上的编码人MHC I、人MHCII α和/或人MHC II β多肽(例如人HLA-A2、人HLA-DRA和/或人HLA-DR 1*04)胞外域的氨基酸。在一个实例中,人MHC I、人MHC II α和/或人MHC II β多肽基本上完整的胞外域缺乏人先导序列。在另一个实例中,人源/非人源嵌合MHC I、人源/非人源嵌合MHC II α和/或人源/非人源嵌合MHC II β多肽包含人先导序列。
此外,所述嵌合MHC I、MHC II α和/或MHC II β多肽可以可操作地连接至(例如,在其调节性控制下表达)内源性非人源启动子和调控元件,例如分别连接至小鼠MHC I、MHCII α和/或MHC II β调控元件。这种排列将有利于所述嵌合MHC I和/或MHC II多肽在非人动物中,例如在非人动物免疫应答期间适当表达。
基因修饰的非人动物可以选自由以下构成的组:小鼠、大鼠、家兔、猪、牛(例如奶牛、公牛、水牛)、鹿、绵羊、山羊、鸡、猫、狗、雪貂、灵长类动物(例如狨猴、猕猴)。对于适宜的基因修饰的ES细胞不易于获得的非人动物而言,可以使用其他方法制备包含基因修饰的非人动物。这类方法包括例如修饰非ES细胞基因组(例如成纤维细胞或诱导多能干细胞)以及使用核转移以便将经修饰的基因组转移至适宜细胞例如卵母细胞,并且在适宜条件下在非人动物中使经修饰的细胞(例如经修饰的卵细胞)妊娠以形成胚胎。
在一个方面,所述非人动物是哺乳动物。在一个方面,所述非人动物是小型哺乳动物,例如跳鼠科或鼠科超家族。在一个实施方式中,所述基因修饰的动物是啮齿动物。在一个实施方式中,所述啮齿动物选自小鼠、大鼠和仓鼠。在一个实施方式中,所述啮齿动物选自鼠科超家族。在一个实施方式中,所述基因修饰的动物来自选自丽仓鼠科(例如小鼠样仓鼠)、仓鼠科(例如仓鼠、新世界大鼠和小鼠、田鼠)、鼠科(真小鼠和大鼠、沙鼠、刺毛鼠、冠毛大鼠)、马岛鼠科(登山小鼠、岩小鼠、有尾大鼠、马达加斯加大鼠和小鼠)、刺山鼠科(例如多刺睡鼠)和鼹形鼠科(例如摩尔大鼠、竹大鼠和鼢鼠)家族。在一个特定实施方式中,所述基因修饰的啮齿动物选自真小鼠或大鼠(鼠科)、沙鼠、刺毛鼠和黄冠鼠。在一个实施方式中,所述基因修饰的小鼠来自鼠科家族成员。在一个实施方式中,所述动物是啮齿动物。在一个特定实施方式中,所述啮齿动物选自小鼠和大鼠。在一个实施方式中,所述非人动物是小鼠。
在一个特定实施方式中,所述非人动物是啮齿动物,其为选自C57BL/A、C57BL/An、C57BL/GrFa、C57BL/KaLwN、C57BL/6、C57BL/6J、C57BL/6ByJ、C57BL/6NJ、C57BL/10、C57BL/10ScSn、C57BL/10Cr和C57BL/Ola的C57BL品系的小鼠。在另一个实施方式中,所述小鼠是选自由以下品系构成的组的129品系:129P1、129P2、129P3、129X1、129S1(例如129S1/SV、129S1/SvIm)、129S2、129S4、129S5、129S9/SvEvH、129S6(129/SvEvTac)、129S7、129S8、129T1、129T2(参见例如Festing等(1999)Revised nomenclature for strain 129 mice,Mammalian Genome 10:836,亦参见Auerbach等(2000)Establishment and ChimeraAnalysis of 129/SvEv-and C57BL/6-Derived Mouse Embryonic Stem Cell Lines)。在一个特定实施方式中,所述基因修饰的小鼠是前述129品系和前述C57BL/6品系的混合。在另一个特定实施方式中,所述小鼠是前述129品系的混合或前述BL/6品系的混合。在一个特定实施方式中,所述混合的129品系是129S6(129/SvEvTac)品系。在另一个实施方式中,所述小鼠是BALB品系,例如BALB/c品系。在又另一个实施方式中,所述小鼠是BALB品系和另一个前述品系的混合。
在一个实施方式中,所述非人动物是大鼠。在一个实施方式中,所述大鼠选自Wistar大鼠、LEA品系、Sprague Dawley品系、Fischer品系、F344、F6和黑刺鼠。在一个实施方式中,所述大鼠品系是两种或多种选自下组的品系的混合:Wistar、LEA、SpragueDawley、Fischer、F344、F6和黑刺鼠。
因此,在一个实施方式中,本发明涉及基因修饰的小鼠,在其基因组中包含编码人/小鼠嵌合MHC I的第一核苷酸序列、编码人/小鼠嵌合MHC II α的第二核苷酸序列和编码人/小鼠嵌合MHC II β多肽的第三核苷酸序列。所述嵌合MHC I、MHC II α和MHC II β的人源部分可分别包含人MHC I、MHC II α和MHC II β的胞外域。在一个实施方式中,所述小鼠表达来自其内源性小鼠MHC基因座的功能性人/小鼠嵌合MHC I、MHC II α和MHC II β多肽。在一个实施方式中,所述小鼠不表达来自其内源性小鼠MHC基因座的功能性小鼠MHC多肽,例如,功能性小鼠MHC I、MHC II α和MHC II β多肽。
在一个实施方式中,所述人/小鼠嵌合MHC I多肽的人源部分包含人MHC I(例如人HLA-A,例如人HLA-A2,例如人HLA-A2.1)的肽结合域或胞外域。在一些实施方式中,所述小鼠不表达来自其内源性小鼠MHC I基因座的肽结合域或胞外域。所述人MHC I的肽结合域可包含α1和α2域。或者,所述人MHC I的肽结合域可包含α1、α2和α3域。在一个方面,所述人MHCI的胞外域包含人MHC I α链的胞外域。在一个实施方式中,所述内源性小鼠MHC I基因座为H-2K(例如,H-2Kb)基因座,且所述嵌合MHC I多肽的小鼠部分包含小鼠H-2K(例如,H-2Kb)多肽的跨膜和胞质域。因此,在一个实施方式中,本发明所述小鼠在其内源性小鼠MHC I基因座包含编码人/小鼠嵌合MHC I的核苷酸序列,其中所述嵌合多肽的人源部分包含人HLA-A2(例如,HLA-A2.1)多肽的胞外域且小鼠部分包含小鼠H-2K(例如,H-2Kb)多肽的跨膜和胞质域,且小鼠表达人/小鼠嵌合HLA-A2/H-2K蛋白。在其它实施方式中,所述嵌合MHC I多肽的小鼠部分可来源于其它小鼠MHC I,例如,H-2D、H-2L等;且所述嵌合MHC I多肽的人源部分可来源于其它人MHC I,例如,HLA-B、HLA-C等。在一个方面,所述小鼠不表达来自其内源性小鼠H-2K基因座的功能性内源性H-2K多肽。
在一个实施方式中,所述人/小鼠嵌合MHC II α多肽的人源部分包含人MHC II α肽结合域或胞外域且所述人/小鼠嵌合MHC II β多肽的人源部分包含人MHC II β肽结合域或胞外域。在一些实施方式中,所述小鼠不表达来自内源性小鼠基因座(例如,H-2A和/或H-2E基因座)的内源性小鼠α和/或β多肽的肽结合域或胞外域。在一些实施方式中,所述小鼠包含缺乏编码包括H-2Ab1、H-2Aa、H-2Eb1、H-2Eb2、H-2Ea及其组合的功能性MHC II类分子的基因的基因组。所述人MHC II α多肽的肽结合域可包含α1域并且所述人MHC II β多肽的肽结合域可包含β1结构域;因此,所述嵌合MHC II复合物的肽结合域可包含人α1和β1结构域。所述人MHC II α多肽的胞外域可包含α1和α2域并且所述人MHC II β多肽的胞外域可包含β1和β2结构域;因此,所述嵌合MHC II复合物的胞外域可包含人α1、α2、β1和β2结构域。在一个实施方式中,所述嵌合MHC II复合物的小鼠部分包含小鼠MHC II的跨膜和胞质域,例如小鼠H-2E(例如小鼠H-2E α和β链的跨膜和胞质域)。因此,在一个实施方式中,本发明所述小鼠在其内源性小鼠MHC II基因座包含编码人/小鼠嵌合MHC II α的核苷酸序列,其中所述嵌合MHC II α多肽的人源部分包含来源于人MHC II的α链(例如,HLA-DR4的α链)的胞外域且小鼠部分包含来源于小鼠MHC II(例如,H-2E)的α链的跨膜和胞质域;且小鼠在其内源性小鼠MHC II基因座包含编码人/小鼠嵌合MHC II β的核苷酸序列,其中所述嵌合MHCII β多肽的人源部分包含来源于人MHC II的β链(例如,HLA-DR4的β链)的胞外域且小鼠部分包含来源于小鼠MHC II(例如,H-2E)的β链的跨膜和胞质域;其中所述小鼠表达人/小鼠嵌合HLA-DR4/H-2E蛋白。在其它实施方式中,所述嵌合MHC II蛋白的小鼠部分可来源于其它小鼠MHC II,例如,H-2A等;且所述嵌合MHC II蛋白的人源部分可来源于其它人MHC II,例如,HLA-DQ等。在一个方面,所述小鼠不表达来自其内源性小鼠基因座的功能性内源性H-2A和H-2E多肽(例如,所述小鼠不表达H-2Ab1、H-2Aa、H-2Eb1、H-2Eb2和H-2Ea多肽)。
在一另外的实施方式中,本发明的非人动物,例如啮齿动物,例如小鼠,在其内源性β2微球蛋白基因座包含编码人源或人源化β2微球蛋白的核苷酸序列。β2微球蛋白或所述MHC I类复合物的轻链(还缩写为“β2M”)是一种小的(12kDa)非糖基化蛋白,其主要功能为稳定化MHC I α链。人源或人源化微球蛋白动物的生成在美国专利申请号13/661,159中详细描述,其通过引入并入本申请。包含如本发明公开内容中所述的人源化MHC基因座以及如美国专利申请号13/661,159中所述的人源或人源化β2微球蛋白基因座的小鼠,可通过本领域中已知的任何方法生成(例如繁殖)。
基因修饰的非人动物(例如啮齿动物,例如大鼠或小鼠)的多种其它实施方式将通过本发明的公开内容以及通过美国专利申请号13/661,159的公开内容显示,后者通过引用并入本申请。
在本发明的不同方面,编码人源/非人源嵌合MHC I和MHC II多肽的序列位于内源性非人源MHC基因座(例如,小鼠H-2K和/或H-2E基因座)。在一个实施方式中,这导致用编码人源或人源化MHC I多肽的核苷酸序列取代内源性MHC基因或其一部分。由于所述编码MHCI、MHC II α和MHC II β多肽的核苷酸序列在染色体上的定位彼此接近,为在一种动物中取得对MHC I和MHC II二者的人源化的最大成功,所述MHC I和MHC II基因座应被连续靶向。因此,本申请还提供了生成基因修饰的非人动物的方法,所述非人动物包含编码如本申请所述的人源/非人源嵌合MHC I、MHC II α和MHC II β多肽的核苷酸序列。
在一些实施方式中,如实施例中所描述的,所述方法利用使用 技术制备的靶向构建体,将所述构建体引入ES细胞并使用
技术制备的靶向构建体,将所述构建体引入ES细胞并使用 技术将被靶向的ES细胞克隆引入小鼠胚胎。
技术将被靶向的ES细胞克隆引入小鼠胚胎。
本申请还提供了用于产生本申请所述非人动物的核苷酸构建体。在一个方面,所述核苷酸构建体包含:5’和3’非人同源臂、包含人MHC基因序列(例如,人HLA-A2或人HLA-DR4基因序列)的人DNA片段以及被重组位点翼侧包围的选择性表达盒(选择性表达盒)。在一个实施方式中,所述人DNA片段是同时包含人MHC基因(例如,人HLA-A2或HLA-DR4基因)的内含子和外显子的基因组片段。在一个实施方式中,所述非人同源臂与非人MHC基因座(例如,MHC I或MHC II基因座)具有同源性。特定构建体描述在以下实施例(例如,图6针对MHCII构建体,MAID 1680;图8针对MHC I构建体,MAID 1665)以及美国申请号13/661,159和13/661,116中,其通过引用并入本申请。
选择性表达盒是插入靶向构建体的核苷酸序列,其有助于选择已整合了所关注构建体的细胞(例如ES细胞)。多种适宜的选择性表达盒是本领域已知的。通常,在存在特定抗生素(例如Neo、Hyg、Pur、CM、Spec等)的条件下选择性表达盒能够得到阳性选择。此外,选择性表达盒可以被重组位点翼侧包围,这使得在使用重组酶处理后可以将选择性表达盒去除。常用的重组位点是loxP和Frt,其分别由Cre和Flp酶识别,但其它的也是本领域已知的。在一个实施方式中,所述选择性表达盒位于人DNA片段的5’末端。在另一个实施方式中,所述选择性表达盒位于人DNA片段的3’末端。在另一个实施方式中,所述选择性表达盒位于人DNA片段内。在另一个实施方式中,所述选择性表达盒位于人DNA片段的内含子内。
在一个实施方式中,所述5’和3’非人同源臂在内源性非人源(例如,鼠类)MHC I类或II类基因座(例如,所述小鼠MHC I基因的第一先导序列的5’和α3外显子的3’,或小鼠H-2Ab1基因的上游和小鼠H-2Ea基因的下游)的5’和3’位分别包含基因组序列。在一个实施方式中,所述内源性MHC I类基因座选自小鼠H-2K、H-2D和H-2L。在一个特定实施方式中,所述内源性MHC I类基因座为小鼠H-2K。在一个实施方式中,所述内源性MHC II基因座选自小鼠H-2E和H-2A。在一个实施方式中,所述基因工程MHC II构建体允许小鼠H-2E和H-2A基因都被取代。
因此,在一个实施方式中,本申请提供的是一种生成可表达人源化MHC I和II蛋白的基因工程非人动物(例如啮齿动物,例如大鼠或小鼠)的方法,其包括在内源性非人源MHCII基因座用编码人源/非人源嵌合MHC II复合物的核苷酸序列取代编码非人源MHC II复合物的核苷酸序列以生成第一非人动物;以及在内源性非人源MHC I基因座用编码人源/非人源嵌合MHC I多肽的核苷酸序列取代编码非人源MHC I多肽的核苷酸序列以生成第二非人动物。在一个实施方式中,所述取代核苷酸序列的步骤包括在ES细胞中同源重组。在一个实施方式中,所述第二非人动物的生成是通过在具有编码人源/非人源嵌合MHC II复合物的核苷酸序列的ES细胞中同源重组。或者,本申请还提供的是一种生成可表达人源化MHC I和II蛋白的基因工程非人动物(例如啮齿动物,例如大鼠或小鼠)的方法,其包括在内源性非人源MHC I基因座用编码人源/非人源嵌合MHC I多肽的核苷酸序列取代编码非人源MHC I多肽的核苷酸序列以生成第一非人动物;以及在内源性非人源MHC II基因座用编码人源/非人源嵌合MHC II复合物的核苷酸序列取代编码非人源MHC II复合物的核苷酸序列以生成第二非人动物。在此实施方式中,所述第二非人动物的生成是通过在具有编码人源/非人源嵌合MHC I多肽的核苷酸序列的ES细胞中同源重组。
在基因靶向完成后,对ES细胞或基因修饰的非人动物进行筛选以确证成功并入了所关注的外源性核苷酸序列或表达了外源性多肽。多种技术是本领域技术人员已知的,并包括(但不限于)Southern印迹、长PCR、定量PCT(例如使用 的实时PCR)、荧光原位杂交、Northern印迹、流式细胞术、Western分析、免疫细胞化学、免疫组化等。在一个实例中,可以通过筛选小鼠等位基因的缺失情况和/或人等位基因的获得情况对携带所关注基因修饰的非人动物(例如小鼠)进行鉴定,其采用Valenzuela等,(2003)High-throughputengineering of the mouse genome coupled with high-resolution expressionanalysis,Nature Biotech.21(6):652-659中所述的经修订的等位基因检测。鉴定基因修饰动物中的特定核苷酸或氨基酸序列的其他检测是本领域技术人员所已知的。
的实时PCR)、荧光原位杂交、Northern印迹、流式细胞术、Western分析、免疫细胞化学、免疫组化等。在一个实例中,可以通过筛选小鼠等位基因的缺失情况和/或人等位基因的获得情况对携带所关注基因修饰的非人动物(例如小鼠)进行鉴定,其采用Valenzuela等,(2003)High-throughputengineering of the mouse genome coupled with high-resolution expressionanalysis,Nature Biotech.21(6):652-659中所述的经修订的等位基因检测。鉴定基因修饰动物中的特定核苷酸或氨基酸序列的其他检测是本领域技术人员所已知的。
在一个方面,提供了表达人源/非人源嵌合MHC I和MHC II蛋白(例如,HLA-A2/H-2K和HLA-DR4/H-2E蛋白)的细胞。在一个实施方式中,所述细胞包含含有如本申请所述的嵌合MHC I类序列和嵌合MHC II类序列的表达载体。在一个实施方式中,所述细胞选自CHO、COS、293、HeLa和表达病毒核酸序列的视网膜细胞(例如PERC.6TM细胞)。
包含本申请所述HLA-DR4胞外域的嵌合MHC II复合物可被抗-HLA-DR抗体检测。因此,可使用抗-HLA-DR抗体检测和/或选择展示人源/非人源嵌合MHC II多肽的细胞。包含本申请所述HLA-A2胞外域的嵌合MHC I复合物可使用抗-HLA-A,例如抗-HLA-A2抗体进行检测。因此,可使用抗-HLA-A抗体检测和/或选择展示人源/非人源嵌合MHC I多肽的细胞。识别其它HLA等位基因的抗体是市售的或可生成的,并可用于检测/选择。
虽然以下实施例描述了基因组包含分别用编码人/小鼠嵌合HLA-A2/H-2K和HLA-DR4/H-2E蛋白的核苷酸序列取代编码小鼠H-2K、和H-2A和H-2E蛋白的核苷酸序列的基因工程动物,但本领域技术人员应当理解类似策略可用于引入包含其它人MHC I和II基因(其它HLA-A、HLA-B和HLA-C;以及其它HLA-DR、HLA-DP和HLA-DQ基因)的嵌合体。还提供了在内源性MHC基因座包含人源/非人源(例如人/啮齿动物,例如人/小鼠)多重嵌合MHC I和MHC II基因的此类动物。
在不同实施方式中,本申请所述的基因修饰的非人动物生成在细胞表面具有人源或人源化MHC I和II的APC,并且因为复合物基本上所有的组分均是人源或人源化的,导致以人样方式针对T细胞提呈作为表位的肽。本发明的基因修饰的非人动物可以用于在人源化动物中研究人免疫系统的功能;用于鉴定激发免疫应答(例如T细胞表位,例如独特的人癌表位)的抗原和抗原表位,例如用于开发疫苗;用于评估疫苗候选物和其他疫苗策略;用于研究人自身免疫;用于研究人感染性疾病;以及设计基于人MHC表达的其他更好的治疗策略。
实施例
将通过下述的非限制性实施例对本发明进行进一步的说明。列出这些实施例以辅助阐述本发明,但并非旨在并且不应将其解释为以任何方式限制其范围。所述实施例不包括对本领域的普通技术人员公知的常规方法(分子克隆技术等)的具体描述。除非另有明示,否则份为重量份,分子量是平均分子量,温度以摄氏度表示和压力为处于或接近大气压。
实施例1:人/小鼠嵌合MHC II基因座的构建和嵌合MHC II小鼠的生成
实施例1.1:内源性MHC II类H-2A和H-2E基因座的删除
使用 基因工程技术制备用于引入内源性MHC II类H-2Ab1、H-2Aa、H-2Eb1、H-2Eb2和H-2Ea基因删除的靶向载体(参见例如美国专利号6,586,251和Valenzuela等,如上所述)。对细菌人工染色体(BAC)RP23-458i22(Invitrogen)DNA进行修饰以删除内源性MHC II类基因H-2Ab1、H-2Aa、H-2Eb1、H-2Eb2和H-2Ea。
基因工程技术制备用于引入内源性MHC II类H-2Ab1、H-2Aa、H-2Eb1、H-2Eb2和H-2Ea基因删除的靶向载体(参见例如美国专利号6,586,251和Valenzuela等,如上所述)。对细菌人工染色体(BAC)RP23-458i22(Invitrogen)DNA进行修饰以删除内源性MHC II类基因H-2Ab1、H-2Aa、H-2Eb1、H-2Eb2和H-2Ea。
简而言之,分别通过对小鼠BAC DNA H-2Ab1基因的5’端和H-2Ea基因的3’端进行PCR获得上游和下游同源臂。如图5所示,这些同源臂(BHR)用于制备表达盒,该表达盒通过细菌同源重组(BHR)删除包含MHC II类基因座的基因H-2Ab1、H-2Aa、H-2Eb1、H-2Eb2和H-2Ea的~79kb的RP23-458i22。该区域被由lox66和lox71位点翼侧包围的潮霉素表达盒取代。最终的靶向载体从5’至3’包括含有小鼠基因组5’至内源性MHC II类基因座的H-2Ab1基因的序列的34kb同源臂、5’lox66位点、潮霉素表达盒、3’lox71位点和含有小鼠基因组3’至内源性MHC II类基因座的H-2Ea基因的序列的63kb同源臂(MAID5111,见图6)。
使用BAC DNA靶向载体(如上所述)电穿孔小鼠ES细胞以制备包含内源性MHC II基因座删除的经修饰的ES细胞。通过使用TAQMANTM探针的定量PCR检测鉴定含有内源性MHC II类基因座删除的阳性ES细胞(Lie和Petropoulos(1998)Curr.Opin.Biotechnology9:43-48)。通过PCR使用引物5111U F (CAGAACGCCAGGCTGTAAC;SEQ ID NO:1)和5111U R(GGAGAGCAGGGTCAGTCAAC;SEQ ID NO:2)以及探针5111U P(CACCGCCACTCACAGCTCCTTACA;SEQ ID NO:3)确证被删除基因座的上游区,而使用引物5111D F(GTGGGCACCATCTTCATCATTC;SEQ ID NO:4)和5111D R(CTTCCTTTCCAGGGTGTGACTC;SEQ IDNO:5)以及探针5111D P(AGGCCTGCGATCAGGTGGCACCT;SEQ ID NO:6)确证被删除基因座的下游区。使用引物HYGF (TGCGGCCGATCTTAGCC;SEQ ID NO:7)和HYGR (TTGACCGATTCCTTGCGG;SEQ ID NO:8)以及探针HYGP(ACGAGCGGGTTCGGCCCATTC;SEQ ID NO:9)确证靶向载体中存在的潮霉素表达盒。跨上游删除点的核苷酸序列(SEQ ID NO:10)包括下述,其表明了删除点上游的内源性小鼠序列(在下文中的括号内)连续地连接至删除点处存在的表达盒序列:(TTTGTAAACA AAGTCTACCC AGAGACAGAT GACAGACTTC AGCTCCAATGCTGATTGGTT CCTCACTTGGGACCAACCCT)CTCGAGTACC GTTCGTATAA TGTATGCTAT ACGAAGTTAT ATGCATCCGG GTAGGGGAGG。跨下游删除点的核苷酸序列(SEQ ID NO:11)包括下述,其表明了删除点下游的内源性小鼠序列(在下文中的括号内)连续地连接至表达盒序列:CCTCGACCTG CAGCCCTAGG ATAACTTCGTATAATGTATG CTATACGAAC GGTAGAGCTC(CACAGGCATT TGGGTGGGCA GGGATGGACG GTGACTGGGACAATCGGGAT GGAAGAGCAT AGAATGGGAG TTAGGGAAGA)。然后使用 法(如下所述)将阳性ES细胞克隆植入雌性小鼠中以生成含有内源性MHC II类基因座删除的一窝胎仔。
法(如下所述)将阳性ES细胞克隆植入雌性小鼠中以生成含有内源性MHC II类基因座删除的一窝胎仔。
使用上文所述的被靶向的ES细胞作为供体ES细胞并通过 法(参见例如美国专利号7,294,754和Poueymirou等,(2007)F0 generation mice that areessentially fully derived from the donor gene-targeted ES cells allowingimmediate phenotypic analyses,Nature Biotech.25(1):91-99)将其引入至8-细胞阶段的小鼠胚胎中。使用检测潮霉素表达盒的存在情况并且确证不存在内源性MHC II类序列的改良的等位基因检测(Valenzuela等,同上)进行基因型鉴定得到在内源性MHC II类基因座携带H-2Ab1、H-2Aa、H-2Eb1、H-2Eb2和H-2Ea基因删除的小鼠。
法(参见例如美国专利号7,294,754和Poueymirou等,(2007)F0 generation mice that areessentially fully derived from the donor gene-targeted ES cells allowingimmediate phenotypic analyses,Nature Biotech.25(1):91-99)将其引入至8-细胞阶段的小鼠胚胎中。使用检测潮霉素表达盒的存在情况并且确证不存在内源性MHC II类序列的改良的等位基因检测(Valenzuela等,同上)进行基因型鉴定得到在内源性MHC II类基因座携带H-2Ab1、H-2Aa、H-2Eb1、H-2Eb2和H-2Ea基因删除的小鼠。
可以将在内源性MHC II类基因座H-2Ab1、H-2Aa、H-2Eb1、H-2Eb2和H-2Ea基因删除的小鼠与Cre基因清除小鼠品系交配繁殖(参见例如国际专利申请公开号WO2009/114400)以除去由靶向载体引入的、在例如ES细胞阶段或在胚胎中未除去的任何被lox位点翼侧包围的潮霉素表达盒。任选地,在小鼠中保留潮霉素表达盒。
实施例1.2:含有人源化H-2Eb1和H-2Ea基因的大型靶向载体(LTVEC)的生成
如图5所示设计引入人源化MHC II序列的靶向载体。使用 基因工程技术,通过多个步骤修饰细菌人工染色体(BAC)RP23-458i22 DNA:(1)制备包含来自BALB/c H-2Ea基因的功能性I-Eα外显子1的载体(图5A);(2)制备包含用人DRβ1*04的外显子2和3取代小鼠I-E β基因的外显子2和3以及用人DRα1*01的外显子2和3取代小鼠I-Eα的外显子2和3的载体(图5B);(3)制备载体,其携带人DRβ1*04的外显子2和3以及其余的小鼠I-Eβ外显子,并携带人DRα1*01的外显子2和3以及其余的小鼠I-Eα外显子,包括来自BALB/c小鼠的功能性I-Eα外显子1(步骤(1),图5C);以及(4)在步骤(3)得到的载体中除去隐蔽剪接位点(图5D)。
基因工程技术,通过多个步骤修饰细菌人工染色体(BAC)RP23-458i22 DNA:(1)制备包含来自BALB/c H-2Ea基因的功能性I-Eα外显子1的载体(图5A);(2)制备包含用人DRβ1*04的外显子2和3取代小鼠I-E β基因的外显子2和3以及用人DRα1*01的外显子2和3取代小鼠I-Eα的外显子2和3的载体(图5B);(3)制备载体,其携带人DRβ1*04的外显子2和3以及其余的小鼠I-Eβ外显子,并携带人DRα1*01的外显子2和3以及其余的小鼠I-Eα外显子,包括来自BALB/c小鼠的功能性I-Eα外显子1(步骤(1),图5C);以及(4)在步骤(3)得到的载体中除去隐蔽剪接位点(图5D)。
特别地,因为在C57B1/6小鼠中,非功能性外显子1的存在使得I-Eα基因是假基因,所以首先制备包含来自BALB/c H-2Ea基因的功能性I-Eα外显子1的载体(图5A)。利用细菌同源重组(1.BHR)对RP23-458i22BAC进行修饰以便使用大观霉素取代氯霉素抗性基因。利用BHR对所得到的载体进行进一步修饰以便用被重组位点(2.BHR)翼侧包围的新霉素表达盒取代整个I-A和I-E编码区。使用含有编码BALB/c I-Eα先导序列的外显子(外显子1)及被PI-SceI和I-CeuI限制性位点翼侧包围的氯霉素基因的构建体进行另一轮BHR(3.BHR)使得载体包含功能性BALB/c H-2Ea外显子1。
独立地,为了产生包含用人DRβ1*04的外显子2和3取代小鼠I-Eβ基因的外显子2和3以及用人DRα1*01的外显子2和3取代小鼠I-Eα的外显子2和3的载体,通过若干同源重组步骤对RP23-458i22BAC进行修饰,4.BHR-8.BHR(图5B)。将所得到的核酸序列用PI-SceI/I-CeuI限制性位点翼侧包围,以使得其能够连接至携带上文所述的包含BALB/c I-Eα外显子1的构建体中(图5C)。
图5C中所述的最终构建体的序列在BALB/c内含子的3’末端含有隐蔽剪接位点。几个BHR步骤(11.BHR-12.BHR)后进行了删除步骤以获得被用于电穿孔至ES细胞的最终靶向载体(MAID1680)(图5D)。
具体的,最终的靶向载体(MAID1680),从5’至3’包含5’小鼠同源臂,其由末端紧邻内源性MHC II类基因座的H-2Ab1基因上游的~26kb小鼠基因组序列组成;含有人源化MHCII β链基因(人源化H-2Eb1基因)和人源化MHC II α链基因(人源化H-2Ea基因)的~59kb插入以及被lox位点翼侧包围的新霉素表达盒;和3’小鼠同源臂,其由起始端紧邻内源性MHCII类基因座的H-2Ea基因下游的~57kb小鼠基因组序列组成。跨5’臂和所述插入之间接头的核苷酸序列(SEQ ID NO:12)包括以下:


在所述~59kb插入中,对H-2Eb1基因进行以下修饰:使H-2Eb15136bp的区域(包括内含子1的最后153bp、外显子2、内含子2、外显子3以及内含子3的起始122bp)被311ibp的人HLA-DRB1*04同源区取代,后者包括内含子1的最后148bp、外显子2、内含子2、外显子3和内含子3的起始132bp。在人和小鼠内含子3序列之间的接头处,插入由5’lox2372位点、UbC启动子、新霉素抗性基因和3’lox2372位点组成的表达盒。所得到的编码嵌合HLA-DRB1*04/H-2Eb1蛋白的基因包括小鼠H-2Eb1先导序列、来自DRB1*04的人β1和β2域以及小鼠跨膜域和胞质尾。内含子1中的跨小鼠/人接头的核苷酸序列(SEQ ID NO:14)包括以下:

 其中用斜体表示的序列是在克隆步骤中引入的多克隆位点,括号中为小鼠内含子1的序列。跨人内含子3和新霉素表达盒接头的核苷酸序列(SEQ ID NO:15)包括以下:
其中用斜体表示的序列是在克隆步骤中引入的多克隆位点,括号中为小鼠内含子1的序列。跨人内含子3和新霉素表达盒接头的核苷酸序列(SEQ ID NO:15)包括以下:

 其中5’lox2372位点用斜体表示,括号中为人内含子3序列。跨新霉素表达盒和小鼠内含子3接头的核苷酸序列(SEQ ID NO:16)包括以下:
其中5’lox2372位点用斜体表示,括号中为人内含子3序列。跨新霉素表达盒和小鼠内含子3接头的核苷酸序列(SEQ ID NO:16)包括以下:
 其中3’lox2372位点用斜体表示,括号中为小鼠内含子3序列。
其中3’lox2372位点用斜体表示,括号中为小鼠内含子3序列。
亦在所述~59kb插入中,对H-2Ea基因进行以下修饰:使H-2Ea 1185bp的区域(包括内含子1的最后101bp、外显子2、内含子2、外显子3和内含子3的起始66bp)被1189bp的人HLA-DRA1*01同源区取代,后者包含内含子1的最后104bp、外显子2、内含子2、外显子3和内含子3的起始66bp。如上所述,由于H-2Ea C57BL/6等位基因的外显子1含有使该基因不具有功能的缺失,因而用来自具有功能的H-2Ea BALB/c等位基因的等量2616bp区域取代H-2Ea外显子1和剩余的内含子1。所得到的编码嵌合H-2Ea/HLA-DRA1*01蛋白的基因含有来自BALB/c的小鼠H-2Ea先导序列、来自DRA1*01的人α1和α2域以及小鼠跨膜域和胞质尾。内含子1中的跨小鼠/人接头的核苷酸序列(SEQ ID NO:17)包括以下:
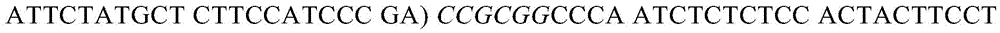
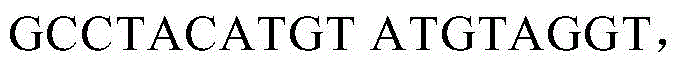 其中用斜体表示的序列是在克隆步骤中引入的限制性酶位点,括号中为BALB/c内含子1的序列。内含子3中的跨人/小鼠接头的核苷酸序列(SEQ IDNO:18)包括以下:
其中用斜体表示的序列是在克隆步骤中引入的限制性酶位点,括号中为BALB/c内含子1的序列。内含子3中的跨人/小鼠接头的核苷酸序列(SEQ IDNO:18)包括以下:
 其中用斜体表示的序列是在克隆步骤中引入的限制性酶位点,括号中为小鼠内含子3序列。外显子1中的跨C57BL/6-BALB/c5’接头的核苷酸序列(SEQ ID NO:19)包括以下:(GAAAGCAGTCTTCCCAGCCT TCACACTCAG AGGTACAAAT)CCCCATTTTC ATATTAGCGA TTTTAATTTA TTCTAGCCTC,其中括号中为C57BL/6-特异性序列。外显子1中的跨BALB/c-C57BL/63’接头的核苷酸序列(SEQ ID NO:20)包括以下:
其中用斜体表示的序列是在克隆步骤中引入的限制性酶位点,括号中为小鼠内含子3序列。外显子1中的跨C57BL/6-BALB/c5’接头的核苷酸序列(SEQ ID NO:19)包括以下:(GAAAGCAGTCTTCCCAGCCT TCACACTCAG AGGTACAAAT)CCCCATTTTC ATATTAGCGA TTTTAATTTA TTCTAGCCTC,其中括号中为C57BL/6-特异性序列。外显子1中的跨BALB/c-C57BL/63’接头的核苷酸序列(SEQ ID NO:20)包括以下: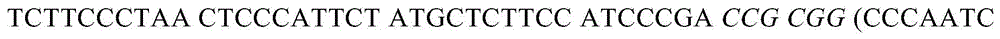
 其中SacII限制性位点用斜体表示,括号中为C57BL/6的序列。
其中SacII限制性位点用斜体表示,括号中为C57BL/6的序列。
实施例1.3:人源化MHC II小鼠的生成
使用实施例1.2载体生成人源化MHC II小鼠的策略简图呈现于图6。
特别地,使用MAID1680BAC DNA(如上所述)电穿孔MAID5111ES细胞以制备经修饰的ES细胞,所述细胞包含内源性小鼠I-A和I-E基因座被包含人源DR4/小鼠I-E嵌合基因座的基因组片段取代。通过使用TAQMANTM探针的定量PCR检测(Lie和Petropoulos,同上)鉴定阳性ES细胞,所述细胞含有删除的内源性I-A和I-E基因座被包含人DR4/小鼠I-E嵌合基因座的基因组片段取代。通过使用引物hDRA1F(CTGGCGGCTTGAAGAATTTGG;SEQ ID NO:21)、hDRA1R(CATGATTTCCAGGTTGGCTTTGTC;SEQ ID NO:22)和探针hDRA1P(CGATTTGCCAGCTTTGAGGCTCAAGG;SEQ ID NO:23)的PCR确证人DR序列的插入。通过使用引物hDRB1F(AGGCTTGGGTGCTCCACTTG;SEQ ID NO:24)、hDRB1R(GACCCTGGTGATGCTGGAAAC;SEQ IDNO:25)和探针hDRB1P(CAGGTGTAAACCTCTCCACTCCGAGGA;SEQ ID NO:26)的PCR确证人DR序列的插入。使用引物HYGF(TGCGGCCGATCTTAGCC;SEQ ID NO:7)和HYGR(TTGACCGATTCCTTGCGG;SEQ ID NO:8)和探针HYGP(ACGAGCGGGTTCGGCCCATTC;SEQ ID NO:9)确证从靶向载体上除去了潮霉素表达盒。
然后使用 法(同上)将阳性ES细胞克隆植入雌性小鼠以产生含有内源性I-A和I-E基因座被人DR4/小鼠I-E嵌合基因座取代的一窝胎仔。使用上述被靶向的ES细胞作为供体细胞并通过
法(同上)将阳性ES细胞克隆植入雌性小鼠以产生含有内源性I-A和I-E基因座被人DR4/小鼠I-E嵌合基因座取代的一窝胎仔。使用上述被靶向的ES细胞作为供体细胞并通过 法引入8细胞阶段的小鼠胚胎。使用检测人DR4/小鼠I-E嵌合基因座的存在情况的改良的等位基因检测(Valenzuela等,同上)通过基因分型鉴定携带人DR4/小鼠I-E嵌合基因座的小鼠。
法引入8细胞阶段的小鼠胚胎。使用检测人DR4/小鼠I-E嵌合基因座的存在情况的改良的等位基因检测(Valenzuela等,同上)通过基因分型鉴定携带人DR4/小鼠I-E嵌合基因座的小鼠。
可以将携带人DR4/小鼠I-E嵌合基因座的小鼠与Cre基因清除小鼠品系交配(参见,例如国际专利申请公开号WO2009/114400)以除去由靶向载体引入的在例如ES细胞阶段或在胚胎中未除去的所有被lox位点翼侧包围的潮霉素表达盒(参见图7)。
实施例2:人/小鼠嵌合MHC I基因座的构建和嵌合MHC I小鼠的生成
构建来自人和小鼠细菌人工染色体(BAC)DNA的独特靶向载体,使用 技术(参见,例如美国专利号6,586,251和Valenzuela等,(2003)High-throughput engineering of the mouse genome coupled with high-resolutionexpression analysis.Nat.Biotech.21(6):652-659)将各小鼠H-2K基因一步人源化。通过同源重组用编码人HLA-A基因α1、α2和α3亚基的人基因组DNA取代编码小鼠H-2K基因α1、α2和α3域的基因组DNA对来自小鼠BAC克隆RP23-173k21(Invitrogen)的DNA进行修饰(图8)。
技术(参见,例如美国专利号6,586,251和Valenzuela等,(2003)High-throughput engineering of the mouse genome coupled with high-resolutionexpression analysis.Nat.Biotech.21(6):652-659)将各小鼠H-2K基因一步人源化。通过同源重组用编码人HLA-A基因α1、α2和α3亚基的人基因组DNA取代编码小鼠H-2K基因α1、α2和α3域的基因组DNA对来自小鼠BAC克隆RP23-173k21(Invitrogen)的DNA进行修饰(图8)。
简而言之,在单一靶向事件中,通过靶向载体用编码人HLA-A*0201基因α1、α2和α3域的人基因组DNA取代编码小鼠H-2K基因α1、α2和α3亚基的基因组序列,所述靶向载体包含由loxP位点翼侧包围的潮霉素表达盒,具有5’小鼠同源臂,其含有包括5’非翻译区的小鼠H-2K基因座的5’序列(UTR;5’同源臂如SEQ ID NO:27所示),以及3’小鼠同源臂,其含有小鼠H-2K α3编码序列的3’基因组序列(3’同源臂如SEQ ID NO:28所示)。
用于靶向内源性H-2K基因座的最终构建体从5’至3’包括:(1)5’同源臂,其包含~200bp包括5’UTR的内源性H-2K基因的小鼠5’基因组序列,(2)~1339bp人基因组序列,其包括HLA-A*0201先导序列、HLA-A*0201先导序列/α1内含子、HLA-A*0201 α1外显子、HLA-A*0201 α1-α2内含子、HLA-A*0201 α2外显子、~316bp α2-α3内含子的5’端,(3)5’loxP位点,(4)潮霉素表达盒,(5)3’loxP位点,(6)~580bp人基因组序列,其包括~304bp α2-α3内含子的3’端、HLA-A*0201 α3外显子和(7)3’同源臂,其包含~200bp包括小鼠H-2Kα3与跨膜编码序列之间的内含子的小鼠基因组序列(参见图8针对H-2K靶向载体的示意图)。在靶向载体5’端小鼠/人序列接头处的149个核苷酸的序列如SEQ ID NO:29所示,在靶向载体3’端人/小鼠序列接头处的159个核苷酸的序列如SEQ ID NO:30所示。使用该靶向载体进行同源重组得到了经修饰的小鼠H-2K基因座,其含有与内源性小鼠H-2K跨膜和胞质域的编码序列可操作地连接的编码HLA-A*0201基因α1、α2和α3域的人基因组DNA,其在翻译后将形成人/小鼠嵌合MHC I类蛋白。
使用靶向BAC DNA电穿孔小鼠F1H4 ES细胞以形成用于生成在有核细胞(例如T和B淋巴细胞、巨噬细胞、中性粒细胞)表面表达嵌合MHC I类蛋白的小鼠的经修饰的ES细胞。通过定量TAQMANTM检测鉴定含有人HLA序列插入的ES细胞。设计特定的引物对和探针以检测人HLA序列和相关选择性表达盒(获得等位基因,GOA)的插入以及内源性小鼠序列的缺失(缺失等位基因,LOA)。表2中显示了在定量PCR检测中使用的各探针的名称以及检测部位。
表2:用于检测基因型的探针

可以采用本领域技术人员已知的方法除去选择性表达盒。例如,可以使用表达Cre的构建体转染携带人/小鼠嵌合MHC I类基因座的ES细胞以除去插入含有人HLA-A*0201基因序列的靶向构建体引入的被lox位点翼侧包围的潮霉素表达盒(参见图8)。可以任选地通过与表达Cre重组酶的小鼠交配除去潮霉素表达盒。任选地,所述潮霉素表达盒保留在小鼠中。
使用上述被靶向的ES细胞作为供体ES细胞并通过 法(参见,例如美国专利号7,294,754和Poueymirou等,(2007)F0 generation mice that areessentially fully derived from the donor gene-targeted ES cells allowingimmediate phenotypic analyses Nature Biotech.25(1):91-99)将其引入8-细胞阶段的小鼠胚胎中。通过使用检测独特的人HLA-A*0201基因序列的存在情况的改良的等位基因检测(Valenzuela等,同上)进行基因型鉴定得到独立地携带嵌合MHC I类基因的
法(参见,例如美国专利号7,294,754和Poueymirou等,(2007)F0 generation mice that areessentially fully derived from the donor gene-targeted ES cells allowingimmediate phenotypic analyses Nature Biotech.25(1):91-99)将其引入8-细胞阶段的小鼠胚胎中。通过使用检测独特的人HLA-A*0201基因序列的存在情况的改良的等位基因检测(Valenzuela等,同上)进行基因型鉴定得到独立地携带嵌合MHC I类基因的 (F0小鼠完全来源于供体ES细胞)。
(F0小鼠完全来源于供体ES细胞)。
实施例3:包含嵌合MHC I和MHC II基因的小鼠的生成和表征
实施例3.1:包含嵌合MHC I和II基因的小鼠的生成
包含嵌合MHC I和MHC II基因的小鼠的生成策略在图4示出。特别地,使用MAID1680BAC DNA(以上实施例2中所述的HLA-A2/H-2K BAC)电穿孔MAID1680ES细胞(携带实施例1中所述的人源化MHC II基因的ES细胞)以制备经修饰的ES细胞,所述细胞包含人/小鼠嵌合MHC I和MHC II基因。通过使用TAQMANTM探针的定量PCR检测(Lie和Petropoulos,同上)采用以上实施例1和2中所述的引物和探针鉴定同时含有嵌合MHC I和II基因的阳性ES细胞。
可以采用本领域技术人员已知的方法除去所述HYG和NEO选择性表达盒。例如,可以使用表达Cre的构建体转染携带人/小鼠嵌合MHC I类和II类基因座的ES细胞以除去通过插入靶向构建体被引入的被lox位点翼侧包围的潮霉素和新霉素表达盒(参见图4、7和8)。可选地可通过与表达Cre重组酶的小鼠进行繁殖以除去所述选择性表达盒。可选地,所述选择性表达盒保留在小鼠中。
使用上述同时包含人源化MHC I和II的被靶向的ES细胞作为供体ES细胞并通过 法(参见,例如美国专利号7,294,754和Poueymirou等,(2007)F0generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech.25(1):91-99)将其引入至8-细胞阶段的小鼠胚胎中。使用改良的等位基因检测(Valenzuela等,同上)对携带嵌合MHC I类和II类基因的
法(参见,例如美国专利号7,294,754和Poueymirou等,(2007)F0generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech.25(1):91-99)将其引入至8-细胞阶段的小鼠胚胎中。使用改良的等位基因检测(Valenzuela等,同上)对携带嵌合MHC I类和II类基因的 (F0小鼠完全来源于供体ES细胞)进行基因型鉴定。
(F0小鼠完全来源于供体ES细胞)进行基因型鉴定。
实施例3.2:包含嵌合MHC I和II基因的小鼠的表征
使用胶原酶D(Roche Bioscience)对来自野生型(WT)或双杂合人源化HLA-A2/HLA-DR4小鼠广1666HET/1681 HET”或“H-2K+/1666 MHC-II+/1681”)的脾脏进行灌流并使用ACK裂解缓冲液裂解红细胞。使用荧光团偶联的抗-CD3(17A2)、抗-CD19(1D3)、抗-HLA-A2(BB7.2)和抗-HLA-DR(L243)通过FACS检测人HLA-A2和HLA-DR4在细胞表面的表达情况。使用BD-Fortessa进行流式细胞术。在CD19+B细胞表面上能够明确检测到人HLA-A2和HLA-DR4二者的表达(图9)。
等同物
本领域技术人员将认识到,或者能够使用不超过常规的实验确定本申请所述的发明的特定实施方式的多种等同物。这类等同物旨在包含在下述权利要求中。
本申请通篇引用的非专利文件、专利申请和专利的全部内容通过引用整体并入本申请。
序列表
<110> 瑞泽恩制药公司
<120> 表达人源化主要组织相容性复合物的小鼠
<130> 2010794-0424
<150> 61/767,811
<151> 2013-02-22
<160> 33
<170> PatentIn 3.5版
<210> 1
<211> 19
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 1
cagaacgcca ggctgtaac 19
<210> 2
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 2
ggagagcagg gtcagtcaac 20
<210> 3
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 3
caccgccact cacagctcct taca 24
<210> 4
<211> 22
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 4
gtgggcacca tcttcatcat tc 22
<210> 5
<211> 22
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 5
cttcctttcc agggtgtgac tc 22
<210> 6
<211> 23
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 6
aggcctgcga tcaggtggca cct 23
<210> 7
<211> 17
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 7
tgcggccgat cttagcc 17
<210> 8
<211> 18
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 8
ttgaccgatt ccttgcgg 18
<210> 9
<211> 21
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 9
acgagcgggt tcggcccatt c 21
<210> 10
<211> 140
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 10
tttgtaaaca aagtctaccc agagacagat gacagacttc agctccaatg ctgattggtt 60
cctcacttgg gaccaaccct ctcgagtacc gttcgtataa tgtatgctat acgaagttat 120
atgcatccgg gtaggggagg 140
<210> 11
<211> 140
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 11
cctcgacctg cagccctagg ataacttcgt ataatgtatg ctatacgaac ggtagagctc 60
cacaggcatt tgggtgggca gggatggacg gtgactggga caatcgggat ggaagagcat 120
agaatgggag ttagggaaga 140
<210> 12
<211> 110
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 12
tgctgattgg ttcctcactt gggaccaacc ctaagcttta tctatgtcgg gtgcggagaa 60
agaggtaatg aaatggcaca aggagatcac acacccaaac caaactcgcc 110
<210> 13
<211> 96
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 13
cacatcagtg aggctagaat aaattaaaat cgctaatatg aaaatgggga tttgtacctc 60
tgagtgtgaa ggctgggaag actgctttca agggac 96
<210> 14
<211> 150
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 14
tccatcactt cactgggtag cacagctgta actgtccagc ctgggtaccg agctcggatc 60
cactagtaac ggccgccagt gtgctggaat tcgcccttga tcgagctccc tgggctgcag 120
gtggtgggcg ttgcgggtgg ggccggttaa 150
<210> 15
<211> 80
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 15
atctccatca gaagggcacc ggtataactt cgtataaggt atcctatacg aagttatatg 60
catggcctcc gcgccgggtt 80
<210> 16
<211> 90
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 16
ataacttcgt ataaggtatc ctatacgaag ttatctcgag tggcttacag gtaggtgcgt 60
gaagcttcta caagcacagt tgccccctgg 90
<210> 17
<211> 90
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 17
ctgtttcttc cctaactccc attctatgct cttccatccc gaccgcggcc caatctctct 60
ccactacttc ctgcctacat gtatgtaggt 90
<210> 18
<211> 80
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 18
caaggtttcc tcctatgatg cttgtgtgaa actcggggcc ggccagcatt taacagtaca 60
gggatgggag cacagctcac 80
<210> 19
<211> 80
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 19
gaaagcagtc ttcccagcct tcacactcag aggtacaaat ccccattttc atattagcga 60
ttttaattta ttctagcctc 80
<210> 20
<211> 80
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 20
tcttccctaa ctcccattct atgctcttcc atcccgaccg cggcccaatc tctctccact 60
acttcctgcc tacatgtatg 80
<210> 21
<211> 21
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 21
ctggcggctt gaagaatttg g 21
<210> 22
<211> 24
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 22
catgatttcc aggttggctt tgtc 24
<210> 23
<211> 26
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 23
cgatttgcca gctttgaggc tcaagg 26
<210> 24
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 24
aggcttgggt gctccacttg 20
<210> 25
<211> 21
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 25
gaccctggtg atgctggaaa c 21
<210> 26
<211> 27
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 26
caggtgtaaa cctctccact ccgagga 27
<210> 27
<211> 200
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 27
ggattcccca tctccacagt ttcacttctg cacctaacct gggtcaggtc cttctgtccg 60
gacactgttg acgcgcagtc agctcttacc cccattgggt ggcgcgatca cccaagaacc 120
aatcagtgtc gccgcggacg ctggatataa agtccacgca gcccgcagaa ctcagaagtc 180
gcgaatcgcc gacaggtgcg 200
<210> 28
<211> 200
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 28
gtaaggagag tgtgggtgca gagctggggt cagggaaagc tggagctttc tgcagaccct 60
gagctgctca gggctgagag ctggggtcat gaccctcacc ttcatttctt gtacctgtcc 120
ttcccagagc ctcctccatc cactgtctcc aacatggcga ccgttgctgt tctggttgtc 180
cttggagctg caatagtcac 200
<210> 29
<211> 149
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 29
agtgtcgccg cggacgctgg atataaagtc cacgcagccc gcagaactca gaagtcgcga 60
atcgccgaca ggtgcgatgg ccgtcatggc gccccgaacc ctcgtcctgc tactctcggg 120
ggctctggcc ctgacccaga cctgggcgg 149
<210> 30
<211> 159
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 30
ggtggtgcct tctggacagg agcagagata cacctgccat gtgcagcatg agggtttgcc 60
caagcccctc accctgagat ggggtaagga gagtgtgggt gcagagctgg ggtcagggaa 120
agctggagct ttctgcagac cctgagctgc tcagggctg 159
<210> 31
<211> 27
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 31
agtccttcag cctccactca ggtcagg 27
<210> 32
<211> 25
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 32
taccaccagt acgcctacga cggca 25
<210> 33
<211> 18
<212> DNA
<213> 人工序列
<220>
<223> 合成的
<400> 33
atcctgtacc agagagtg 18
Claims (36)
1.一种基因修饰的啮齿动物细胞,所述细胞在其基因组中包含:
(i)编码嵌合人源和啮齿动物MHC I多肽的第一核苷酸序列,其中所述嵌合人源和啮齿动物MHC I多肽的人源部分包含人MHC I多肽的α1、α2和α3域,并且其中所述嵌合人源和啮齿动物MHC I多肽的所述啮齿动物部分包含内源性啮齿动物MHC I多肽的跨膜和胞质域,
(ii)编码嵌合人源和啮齿动物MHC IIα多肽的第二核苷酸序列,其中所述嵌合人源和啮齿动物MHC IIα多肽的人源部分包含人MHC IIα多肽的α1和α2域,并且其中所述嵌合人源和啮齿动物MHC IIα多肽的啮齿动物部分包含内源性啮齿动物MHC IIα多肽的跨膜和胞质域,和
(iii)编码嵌合人源和啮齿动物MHC IIβ多肽的第三核苷酸序列,其中所述嵌合人源和啮齿动物MHC IIβ多肽的人源部分包含人MHC IIβ多肽的β1和β2域,并且其中所述嵌合人源和啮齿动物MHC IIβ多肽的啮齿动物部分包含内源性啮齿动物MHC IIβ多肽的跨膜和胞质域,
其中所述啮齿动物细胞是大鼠细胞或小鼠细胞。
2.根据权利要求1所述的基因修饰的啮齿动物细胞,其中所述第一核苷酸序列位于内源性啮齿动物MHC I基因座,所述第二核苷酸序列位于内源性啮齿动物MHC IIα基因座,并且所述第三核苷酸序列位于内源性啮齿动物MHC IIβ基因座。
3.根据权利要求1所述的基因修饰的啮齿动物细胞,其中所述第一核苷酸序列可操作地连接至内源性啮齿动物MHC Iα调控元件,所述第二核苷酸序列可操作地连接至内源性啮齿动物MHC IIα调控元件,和/或所述第三核苷酸序列可操作地连接至内源性啮齿动物MHCIIβ调控元件。
4.根据权利要求1所述的基因修饰的啮齿动物细胞,其中所述人MHC I多肽是HLA-A、HLA-B或HLA-C。
5.根据权利要求1所述的基因修饰的啮齿动物细胞,在内源性啮齿动物β2微球蛋白基因座另外包含编码人源或人源化β2微球蛋白多肽的核苷酸序列。
6.根据权利要求1所述的基因修饰的啮齿动物细胞,其中所述第一核苷酸序列在内源性啮齿动物MHC I基因座处可操作地连接至内源性啮齿动物MHC I启动子和调控元件,所述第二核苷酸序列在内源性啮齿动物MHC IIα基因座处可操作地连接至内源性啮齿动物MHCIIα启动子和调控元件,并且所述第三核苷酸序列在内源性啮齿动物MHC IIβ基因座处可操作地连接至内源性啮齿动物MHC IIβ启动子和调控元件。
7.根据权利要求1所述的基因修饰的啮齿动物细胞,其中所述嵌合人源和啮齿动物MHCIIα多肽的人源部分由选自下组的人HLA IIα链基因编码:HLA-DR、HLA-DQ和HLA-DP的任意α链基因,并且其中所述嵌合人源和啮齿动物MHC IIβ多肽的人源部分由选自下组的人HLAIIβ链基因编码:HLA-DR、HLA-DQ和HLA-DP的任意β链基因。
8.根据权利要求1所述的基因修饰的啮齿动物细胞,其中所述嵌合人源和啮齿动物MHCIIα和β多肽的人源部分分别由(a)人HLA-DRα和β链基因编码或(b)由人HLA-DQα和β链基因编码。
9.根据权利要求1所述的基因修饰的啮齿动物细胞,
其中所述啮齿动物细胞是小鼠细胞,
其中所述第一核苷酸序列编码嵌合人源和小鼠MHC I多肽,并且
其中所述嵌合人源和小鼠MHC I多肽的小鼠部分由选自下组的小鼠MHC I基因编码:
H-2K基因、H-2D基因和H-2L基因。
10.根据权利要求9所述的基因修饰的啮齿动物细胞,其中所述嵌合人源和小鼠MHC I多肽的小鼠部分由小鼠H-2K基因编码。
11.根据权利要求1所述的基因修饰的啮齿动物细胞,
其中所述啮齿动物细胞是小鼠细胞,且
其中所述第二核苷酸序列编码嵌合人源和小鼠MHC IIα多肽,所述第三核苷酸序列编码嵌合人源和小鼠MHC IIβ多肽,所述嵌合人源和小鼠MHC IIα多肽的小鼠部分由小鼠MHCII H-2Eα链基因或由小鼠MHC II H-2Aα链基因编码和所述嵌合人源和小鼠MHC IIβ多肽的小鼠部分由小鼠MHC II H-2Eβ链基因或由小鼠MHC II H-2Aβ链基因编码。
12.根据权利要求11所述的基因修饰的啮齿动物细胞,其中所述嵌合人源和小鼠MHCIIα和β多肽的小鼠部分分别由H-2Eα和β链基因编码。
13.根据权利要求1-12中任一项所述的基因修饰的啮齿动物细胞,其中所述细胞是抗原递呈细胞(APC),并且其中所述APC在其表面上表达所述嵌合人源和啮齿动物MHC I多肽和嵌合人源和啮齿动物MHC II复合物,所述MHC II复合物包含来自所述内源性啮齿动物MHC基因座的所述嵌合人源和啮齿动物MHC IIα多肽和嵌合人源和啮齿动物MHC IIβ多肽。
14.根据权利要求13所述的基因修饰的啮齿动物细胞,其中所述APC进一步不表达来自所述内源性啮齿动物MHC基因座的功能性内源性啮齿动物MHC多肽。
15.根据权利要求1-12中任一项所述的基因修饰的啮齿动物细胞,其中所述细胞不是胚胎干细胞。
16.一种基因修饰的小鼠细胞,所述细胞在其基因组中:
(i)在内源性小鼠MHC I基因座处包含编码嵌合人源和小鼠MHC I多肽的第一核苷酸序列,其中所述嵌合人源和小鼠MHC I多肽的人源部分包含人MHC I多肽的α1、α2和α3域,并且其中所述嵌合人源和小鼠MHC I多肽的小鼠部分包含内源性小鼠MHC I多肽的跨膜和胞质域,
(ii)在内源性小鼠MHC IIα基因座处包含编码嵌合人源和小鼠MHC IIα多肽的第二核苷酸序列,其中所述嵌合人源和小鼠MHC IIα多肽的人源部分包含人MHC IIα多肽的α1和α2域,并且其中所述嵌合人源和小鼠MHC IIα多肽的非人源部分包含内源性小鼠MHC IIα多肽的跨膜和胞质域,和
(iii)在内源性小鼠MHC IIβ基因座处包含编码嵌合人源和小鼠MHC IIβ多肽的第三核苷酸序列,其中所述嵌合人源和小鼠MHC IIβ多肽的人源部分包含人MHC IIβ多肽的β1和β2域,并且其中所述嵌合人源和小鼠MHC IIβ多肽的小鼠部分包含内源性小鼠MHC IIβ多肽的跨膜和胞质域。
17.根据权利要求16所述的基因修饰的小鼠细胞,其中所述第一核苷酸序列编码嵌合人源和小鼠HLA-A/H-2K多肽,所述第二核苷酸序列编码嵌合人源和小鼠HLA-DR/H-2E多肽的α链,且所述第三核苷酸序列编码嵌合人源和小鼠HLA-DR/H-2E多肽的β链。
18.根据权利要求17所述的基因修饰的小鼠细胞,其中所述小鼠细胞在内源性小鼠β2微球蛋白基因座还包含编码人源或人源化β2微球蛋白多肽的核苷酸序列。
19.根据权利要求16-18中任一项所述的基因修饰的小鼠细胞,其中所述小鼠细胞是抗原递呈细胞(APC),并且其中所述APC在其表面上表达来自所述内源性小鼠MHC基因座的所述嵌合人源和小鼠MHC I多肽和嵌合人源和小鼠MHC II复合物,所述MHC II复合物包含所述嵌合人源和小鼠MHC IIα多肽和嵌合人源和小鼠MHC IIβ多肽。
20.根据权利要求19所述的基因修饰的小鼠细胞,其中所述小鼠细胞表达人源或人源化β2微球蛋白多肽。
21.根据权利要求16-18中任一项所述的基因修饰的小鼠细胞,其中所述小鼠细胞不是胚胎干细胞。
22.一种基因修饰的啮齿动物基因组,所述基因组包含:
(i)编码嵌合人源和啮齿动物MHC I多肽的第一核苷酸序列,其中所述嵌合人源和啮齿动物MHC I多肽的人源部分包含人MHC I多肽的α1、α2和α3域,并且其中所述嵌合人源和啮齿动物MHC I多肽的啮齿动物部分包含内源性啮齿动物MHC I多肽的跨膜和胞质域,
(ii)编码嵌合人源和啮齿动物MHC IIα多肽的第二核苷酸序列,其中所述嵌合人源和啮齿动物MHC IIα多肽的人源部分包含人MHC IIα多肽的α1和α2域,并且其中所述嵌合人源和啮齿动物MHC IIα多肽的啮齿动物部分包含内源性啮齿动物MHC IIα多肽的跨膜和胞质域,和
(iii)编码嵌合人源和啮齿动物MHC IIβ多肽的第三核苷酸序列,其中所述嵌合人源和啮齿动物MHC IIβ多肽的人源部分包含人MHC IIβ多肽的β1和β2域,并且其中所述嵌合人源和啮齿动物MHC IIβ多肽的啮齿动物部分包含内源性啮齿动物MHC IIβ多肽的跨膜和胞质域,
其中所述啮齿动物基因组 是大鼠基因组或小鼠基因组。
23.根据权利要求22所述的基因修饰的啮齿动物基因组,其中所述第一核苷酸序列位于内源性啮齿动物MHC I基因座,所述第二核苷酸序列位于内源性啮齿动物MHC IIα基因座,并且所述第三核苷酸序列位于内源性啮齿动物MHC IIβ基因座。
24.根据权利要求23所述的基因修饰的啮齿动物基因组,其中所述第一核苷酸序列可操作地连接至内源性啮齿动物MHC Iα调控元件、所述第二核苷酸序列可操作地连接至内源性啮齿动物MHC IIα调控元件,和/或所述第三核苷酸序列可操作地连接至内源性啮齿动物MHC IIβ调控元件。
25.根据权利要求22所述的基因修饰的啮齿动物基因组,其中所述人MHC I多肽是HLA-A、HLA-B或HLA-C。
26.根据权利要求22所述的基因修饰的啮齿动物基因组,在内源性啮齿动物β2微球蛋白基因座另外包含编码人源或人源化β2微球蛋白多肽的核苷酸序列。
27.根据权利要求22所述的基因修饰的啮齿动物基因组,其中所述第一核苷酸序列在内源性啮齿动物MHC I基因座处可操作地连接至内源性啮齿动物MHC I启动子和调控元件,所述第二核苷酸序列在内源性啮齿动物MHC IIα基因座处可操作地连接至内源性啮齿动物MHC IIα启动子和调控元件,并且所述第三核苷酸序列在内源性啮齿动物MHC IIβ基因座处可操作地连接至内源性啮齿动物MHC IIβ启动子和调控元件。
28.根据权利要求22所述的基因修饰的啮齿动物基因组,其中所述嵌合人源和啮齿动物MHC IIα多肽的人源部分由选自下组的人HLA IIα链基因编码:HLA-DR、HLA-DQ和HLA-DP的任意α链基因,并且其中所述嵌合人源和啮齿动物MHC IIβ多肽的人源部分由选自下组的人HLA IIβ链基因编码:HLA-DR、HLA-DQ和HLA-DP的任意β链基因。
29.根据权利要求22所述的基因修饰的啮齿动物基因组,其中所述嵌合人源和啮齿动物MHC IIα和β多肽的人源部分分别(a)由人HLA-DRα和β链基因编码或(b)由人HLA-DQα和β链基因编码。
30.根据权利要求22所述的基因修饰的啮齿动物基因组,
其中所述啮齿动物基因组是小鼠基因组,
其中所述第一核苷酸序列编码嵌合人源和小鼠MHC I多肽,并且
其中所述嵌合人源和小鼠MHC I多肽的小鼠部分由选自下组的小鼠MHC I基因编码:
H-2K基因、H-2D基因或H-2L基因。
31.根据权利要求30所述的基因修饰的啮齿动物基因组,其中所述嵌合人源和小鼠MHCI多肽的小鼠部分由小鼠H-2K基因编码。
32.根据权利要求22所述的基因修饰的啮齿动物基因组,
其中所述啮齿动物基因组是小鼠基因组,
其中所述第二核苷酸序列编码嵌合人源和小鼠MHC IIα多肽,所述第三核苷酸序列编码嵌合人源和小鼠MHC IIβ多肽,所述嵌合人源和小鼠MHC IIα多肽的小鼠部分由小鼠MHCII H-2Eα链基因或由小鼠MHC II H-2Aα链基因编码和所述嵌合人源和小鼠MHC IIβ多肽的小鼠部分由小鼠MHC II H-2Eβ链基因或由小鼠MHC II H-2Aβ链基因编码。
33.根据权利要求32所述的基因修饰的啮齿动物基因组,其中所述嵌合人源和小鼠MHCIIα和β多肽的小鼠部分分别由H-2Eα和β链基因编码。
34.一种基因修饰的小鼠基因组,所述基因组包含:
(i)在内源性MHC I基因座的编码嵌合人源和小鼠MHC I多肽的第一核苷酸序列,其中所述嵌合人源和小鼠MHC I多肽的人源部分包含人MHC I多肽的α1、α2和α3域,并且其中所述嵌合人源和小鼠MHC I多肽的小鼠部分包含内源性小鼠MHC I多肽的跨膜和胞质域,
(ii)在内源性小鼠MHC IIα基因座的编码嵌合人源和小鼠MHC IIα多肽的第二核苷酸序列,其中所述嵌合人源和小鼠MHC IIα多肽的人源部分包含人MHC IIα多肽的α1和α2域,并且其中所述嵌合人源和小鼠MHC IIα多肽的小鼠部分包含内源性小鼠MHC IIα多肽的跨膜和胞质域,和
(iii)在内源性小鼠MHC IIβ基因座的编码嵌合人源和小鼠MHC IIβ多肽的第三核苷酸序列,其中所述嵌合人源和小鼠MHC IIβ多肽的人源部分包含人MHC IIβ多肽的β1和β2域,并且其中所述嵌合人源和小鼠MHC IIβ多肽的小鼠部分包含内源性小鼠MHC IIβ多肽的跨膜和胞质域。
35.根据权利要求34所述的基因修饰的小鼠基因组,其中所述第一核苷酸序列编码嵌合人源和小鼠HLA-A/H-2K多肽,所述第二核苷酸序列编码嵌合人源和小鼠HLA-DR/H-2E多肽的α链,且所述第三核苷酸序列编码嵌合人源和小鼠HLA-DR/H-2E多肽的β链。
36.根据权利要求35所述的基因修饰的小鼠基因组,其中所述小鼠基因组在内源性小鼠β2微球蛋白基因座还包含编码人源或人源化β2微球蛋白多肽的核苷酸序列。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201361767811P | 2013-02-22 | 2013-02-22 | |
| US61/767,811 | 2013-02-22 | ||
| CN201480022254.1A CN105164154B (zh) | 2013-02-22 | 2014-02-20 | 表达人源化主要组织相容性复合物的小鼠 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201480022254.1A Division CN105164154B (zh) | 2013-02-22 | 2014-02-20 | 表达人源化主要组织相容性复合物的小鼠 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| CN110192541A CN110192541A (zh) | 2019-09-03 |
| CN110192541B true CN110192541B (zh) | 2022-02-18 |
Family
ID=50236326
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201910379017.5A Active CN110192541B (zh) | 2013-02-22 | 2014-02-20 | 表达人源化主要组织相容性复合物的小鼠 |
| CN201480022254.1A Active CN105164154B (zh) | 2013-02-22 | 2014-02-20 | 表达人源化主要组织相容性复合物的小鼠 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201480022254.1A Active CN105164154B (zh) | 2013-02-22 | 2014-02-20 | 表达人源化主要组织相容性复合物的小鼠 |
Country Status (19)
| Country | Link |
|---|---|
| US (3) | US10154658B2 (zh) |
| EP (3) | EP2958937B1 (zh) |
| JP (2) | JP6444321B2 (zh) |
| KR (1) | KR102211267B1 (zh) |
| CN (2) | CN110192541B (zh) |
| AU (2) | AU2014218893C1 (zh) |
| CA (1) | CA2900824A1 (zh) |
| CY (1) | CY1121223T1 (zh) |
| DK (1) | DK2958937T3 (zh) |
| ES (3) | ES2949029T3 (zh) |
| HR (1) | HRP20181866T1 (zh) |
| HU (1) | HUE041878T2 (zh) |
| LT (1) | LT2958937T (zh) |
| PL (1) | PL2958937T3 (zh) |
| PT (1) | PT2958937T (zh) |
| RS (1) | RS57963B1 (zh) |
| SI (1) | SI2958937T1 (zh) |
| SM (1) | SMT201800614T1 (zh) |
| WO (1) | WO2014130667A1 (zh) |
Families Citing this family (46)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA2852962C (en) | 2011-10-28 | 2022-05-03 | Regeneron Pharmaceuticals, Inc. | Genetically modified mice expressing chimeric major histocompatibility complex (mhc) ii molecules |
| US9591835B2 (en) | 2011-10-28 | 2017-03-14 | Regeneron Pharmaceuticals, Inc. | Genetically modified major histocompatibility complex animals |
| US9043996B2 (en) | 2011-10-28 | 2015-06-02 | Regeneron Pharmaceuticals, Inc. | Genetically modified major histocompatibility complex animals |
| RS56656B1 (sr) | 2011-10-28 | 2018-03-30 | Regeneron Pharma | Miševi sa genetski modifikovanim glavnim kompleksom gena tkivne podudarnosti |
| PT2958938T (pt) | 2013-02-20 | 2019-07-23 | Regeneron Pharma | Ratinhos que expressam correcetores de células t humanizados |
| US20150342163A1 (en) | 2013-02-22 | 2015-12-03 | Regeneron Pharmaceuticals, Inc. | Genetically modified major histocompatibility complex mice |
| CN110192541B (zh) | 2013-02-22 | 2022-02-18 | 瑞泽恩制药公司 | 表达人源化主要组织相容性复合物的小鼠 |
| CA2903025C (en) * | 2013-03-11 | 2025-10-07 | Regeneron Pharmaceuticals, Inc. | TRANSGENIC MICE EXPRESSING CHIMERIC MAJOR HISTOCOMPATIBILITY COMPLEX (MHC) CLASS II MOLECULES |
| SMT202300287T1 (it) | 2013-09-23 | 2023-11-13 | Regeneron Pharma | Animali non umani aventi un gene umanizzato della proteina regolatrice del segnale |
| RU2674910C2 (ru) | 2013-10-15 | 2018-12-13 | Регенерон Фармасьютикалс, Инк. | Животные с гуманизированным геном il-15 |
| SG11201603360QA (en) | 2013-11-19 | 2016-05-30 | Regeneron Pharma | Non-human animals having a humanized b-cell activating factor gene |
| KR102173297B1 (ko) | 2013-11-19 | 2020-11-03 | 리제너론 파마슈티칼스 인코포레이티드 | 인간화된 증식-유도 리간드 유전자를 가지고 있는 비-인간 동물 |
| HUE051088T2 (hu) | 2014-04-08 | 2021-03-01 | Regeneron Pharma | Humanizált FC-gamma receptorokkal rendelkezõ nem humán élõlények |
| CA3225091A1 (en) | 2014-05-05 | 2015-11-12 | Regeneron Pharmaceuticals, Inc. | Humanized c5 and c3 animals |
| NO2785538T3 (zh) | 2014-05-07 | 2018-08-04 | ||
| SG10201811116UA (en) | 2014-06-19 | 2019-01-30 | Regeneron Pharma | Non-human animals having a humanized programmed cell death 1 gene |
| KR20170083534A (ko) | 2014-09-19 | 2017-07-18 | 리제너론 파마슈티칼스 인코포레이티드 | 키메라 항원 수용체 |
| SI3689140T1 (sl) | 2014-11-24 | 2022-10-28 | Regeneron Pharmaceuticals, Inc. | Nehumane živali, ki izražajo humaniziran CD3 kompleks |
| PL3086637T3 (pl) | 2014-12-05 | 2019-09-30 | Regeneron Pharmaceuticals, Inc. | Zwierzęta nie będące człowiekiem mające humanizowany gen kompleksu różnicowania 47 |
| LT3230320T (lt) | 2014-12-09 | 2021-01-11 | Regeneron Pharmaceuticals, Inc. | Gyvūnai, išskyrus žmogų, turintys humanizuotą diferenciacijos klasterio 274 geną |
| WO2016164492A2 (en) | 2015-04-06 | 2016-10-13 | Regeneron Pharmaceuticals, Inc. | Humanized t cell mediated immune responses in non-human animals |
| WO2017087780A1 (en) | 2015-11-20 | 2017-05-26 | Regeneron Pharmaceuticals, Inc. | Non-human animals having a humanized lymphocyte-activation gene 3 |
| WO2017151453A1 (en) | 2016-02-29 | 2017-09-08 | Regeneron Pharmaceuticals, Inc. | Rodents having a humanized tmprss gene |
| AU2017272337C1 (en) | 2016-06-03 | 2024-02-29 | Regeneron Pharmaceuticals, Inc. | Non-human animals expressing exogenous terminal deoxynucleotidyltransferase |
| EP3585162B1 (en) | 2017-09-29 | 2023-08-30 | Regeneron Pharmaceuticals, Inc. | Rodents comprising a humanized ttr locus and methods of use |
| SG11202002456WA (en) | 2017-11-30 | 2020-04-29 | Regeneron Pharma | Non-human animals comprising a humanized trkb locus |
| DK3772927T3 (da) | 2018-03-24 | 2025-02-17 | Regeneron Pharma | Genetisk modificerede mus eller rotter til generering af terapeutiske antistoffer mod peptid¿MHC-komplekser, fremgangsmåde til fremstilling og anvendelser deraf |
| KR20250121150A (ko) | 2018-03-26 | 2025-08-11 | 리제너론 파마슈티칼스 인코포레이티드 | 치료제를 시험하기 위한 인간화된 설치류 |
| AU2019308205B2 (en) | 2018-07-16 | 2025-11-06 | Regeneron Pharmaceuticals, Inc. | Non-human animal models of DITRA disease and uses thereof |
| ES2966625T3 (es) | 2019-04-04 | 2024-04-23 | Regeneron Pharma | Roedores que comprenden un locus del factor de coagulación 12 humanizado |
| JP7610339B2 (ja) | 2019-06-04 | 2025-01-08 | リジェネロン・ファーマシューティカルズ・インコーポレイテッド | ベータスリップ変異を有するヒト化ttr遺伝子座を含む非ヒト動物と使用方法 |
| CA3137764A1 (en) | 2019-06-07 | 2020-12-10 | Regeneron Pharmaceuticals, Inc. | Non-human animals comprising a humanized albumin locus |
| CN113105555B (zh) * | 2020-01-10 | 2023-05-30 | 百奥赛图(北京)医药科技股份有限公司 | Mhc分子人源化的非人动物的构建方法及应用 |
| CN116981357A (zh) | 2021-03-31 | 2023-10-31 | 瑞泽恩制药公司 | 包含具有改善的tcrb组库多样性的人源化细胞免疫系统组分的基因修饰的小鼠 |
| WO2023240124A1 (en) | 2022-06-07 | 2023-12-14 | Regeneron Pharmaceuticals, Inc. | Pseudotyped viral particles for targeting tcr-expressing cells |
| WO2023240109A1 (en) | 2022-06-07 | 2023-12-14 | Regeneron Pharmaceuticals, Inc. | Multispecific molecules for modulating t-cell activity, and uses thereof |
| CN119947583A (zh) | 2022-09-22 | 2025-05-06 | 瑞泽恩制药公司 | 表达人细胞免疫系统组分的经基因修饰的小鼠 |
| EP4654982A1 (en) | 2023-01-27 | 2025-12-03 | Regeneron Pharmaceuticals, Inc. | Modified rhabdovirus glycoproteins and uses thereof |
| TW202520975A (zh) | 2023-08-09 | 2025-06-01 | 英商劍橋企業有限公司 | 基因改造之囓齒動物及囓齒動物細胞以及其等用途 |
| WO2025064470A1 (en) | 2023-09-19 | 2025-03-27 | Regeneron Pharmaceuticals, Inc. | Viral peptides and uses thereof |
| WO2025064761A1 (en) | 2023-09-22 | 2025-03-27 | Regeneron Pharmaceuticals, Inc. | Kras10-18 g12d off-target peptides and uses thereof |
| WO2025064738A1 (en) | 2023-09-22 | 2025-03-27 | Regeneron Pharmaceuticals, Inc. | Dntt 250-258 off-target peptides and uses thereof |
| WO2025136809A2 (en) | 2023-12-19 | 2025-06-26 | Regeneron Pharmaceuticals, Inc. | Foxp3 peptides and uses thereof |
| WO2025178991A1 (en) | 2024-02-21 | 2025-08-28 | Regeneron Pharmaceuticals, Inc. | Splicing factor gene mutation-associated neoantigenic peptides and uses thereof |
| WO2026006734A1 (en) | 2024-06-28 | 2026-01-02 | Regeneron Pharmaceuticals, Inc. | Off-target peptide-mhc complex conformation modeling systems and methods for antigen-recognition molecule development |
| WO2026006724A1 (en) | 2024-06-28 | 2026-01-02 | Regeneron Pharmaceuticals, Inc. | Prame off-target peptides and uses thereof |
Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5523226A (en) * | 1993-05-14 | 1996-06-04 | Biotechnology Research And Development Corp. | Transgenic swine compositions and methods |
| CN101060776A (zh) * | 2004-11-23 | 2007-10-24 | 安迪拓普有限公司 | Mhc-ⅱ转基因动物耐受性的研究方法 |
| WO2012039779A1 (en) * | 2010-09-24 | 2012-03-29 | The United States Of America As Represented By The Secretary Of The Navy | Humanized transgenic mouse model |
| CN102711449A (zh) * | 2009-12-10 | 2012-10-03 | 瑞泽恩制药公司 | 生产重链抗体的小鼠 |
Family Cites Families (66)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP0950707B1 (en) | 1989-07-25 | 2009-02-18 | Cell Genesys, Inc. | Homologous recombination for universal donor cells and chimeric mammalian hosts |
| US5413923A (en) | 1989-07-25 | 1995-05-09 | Cell Genesys, Inc. | Homologous recombination for universal donor cells and chimeric mammalian hosts |
| US5574205A (en) | 1989-07-25 | 1996-11-12 | Cell Genesys | Homologous recombination for universal donor cells and chimeric mammalian hosts |
| US6150584A (en) | 1990-01-12 | 2000-11-21 | Abgenix, Inc. | Human antibodies derived from immunized xenomice |
| US7041871B1 (en) | 1995-10-10 | 2006-05-09 | Genpharm International, Inc. | Transgenic non-human animals capable of producing heterologous antibodies |
| US5489742A (en) * | 1990-10-23 | 1996-02-06 | Board Of Regents, The University Of Texas System | Transgenic rats and animal models of inflammatory disease |
| GB9100481D0 (en) | 1991-01-10 | 1991-02-20 | Inst Nat Sante Rech Med | Genetically engineered mice |
| AU2661692A (en) | 1991-09-19 | 1993-04-27 | President And Fellows Of Harvard College | Transgenic mhc class i and class ii antigen-deficient mammals |
| WO1994006908A1 (en) | 1992-09-11 | 1994-03-31 | The Regents Of The University Of California | Transgenic non-human animals having targeted lymphocyte transduction genes |
| GB9315303D0 (en) | 1993-07-23 | 1993-09-08 | Zeneca Ltd | Polypeptide |
| US5965787A (en) | 1995-08-31 | 1999-10-12 | Mayo Foundation For Medical Education And Research | HLA-DRBI peptides with specific binding affinity for HLA-DQ molecules: prevention and treatment of rheumatoid arthritis |
| US6002066A (en) | 1996-01-16 | 1999-12-14 | Ortho Pharmaceutical Corp. | H2-M modified transgenic mice |
| JP2000512127A (ja) | 1996-03-05 | 2000-09-19 | ザ スクリップス リサーチ インスティトゥート | ヒトhla拘束性腫瘍抗原に特異的なt細胞受容体をコードする組み換え体 |
| CA2273194C (en) | 1996-12-03 | 2011-02-01 | Abgenix, Inc. | Transgenic mammals having human ig loci including plural vh and vk regions and antibodies produced therefrom |
| ATE423789T1 (de) | 1997-09-16 | 2009-03-15 | Univ Oregon Health & Science | Rekombinante mhc-moleküle welche nützlich sind für die manipulation von antigen-spezifischen t- zellen |
| US6372955B1 (en) | 1998-02-17 | 2002-04-16 | Ortho Mcneil Pharmaceutical, Inc. | Methods for Producing B cells and antibodies from H2-O modified transgenic mice |
| MXPA01010625A (es) * | 1999-04-23 | 2003-09-04 | Pharmexa As | Metodo para regulacion por disminucion de la actividad de interleucina 5. |
| AU1023401A (en) | 1999-10-12 | 2001-04-23 | Institut Pasteur | Design of a polyepitopic construct for the induction of hla-a2.1 restricted hiv 1 specific ctl responses using hhd mice |
| WO2002000933A2 (en) | 2000-06-23 | 2002-01-03 | Interleukin Genetics, Inc. | Screening assays for identifying modulators of the inflammatory or immune responses |
| US6596541B2 (en) | 2000-10-31 | 2003-07-22 | Regeneron Pharmaceuticals, Inc. | Methods of modifying eukaryotic cells |
| US6586251B2 (en) | 2000-10-31 | 2003-07-01 | Regeneron Pharmaceuticals, Inc. | Methods of modifying eukaryotic cells |
| US7105348B2 (en) | 2000-10-31 | 2006-09-12 | Regeneron Pharmaceuticals, Inc. | Methods of modifying eukaryotic cells |
| JPWO2002047474A1 (ja) | 2000-12-13 | 2004-04-15 | 住友製薬株式会社 | Hla−a24発現トランスジェニック動物及びその利用 |
| US20030093818A1 (en) | 2000-12-19 | 2003-05-15 | Belmont Heather J. | Transgenic animals comprising a humanized immune system |
| FR2827302B1 (fr) * | 2001-07-13 | 2003-10-10 | Genoway | Cellule et animal transgenique modelisant la presentation antigenique humaine et leurs utilisations |
| US8895020B2 (en) | 2002-04-19 | 2014-11-25 | Washington University | Single chain trimers and uses therefor |
| US20040072262A1 (en) | 2002-10-11 | 2004-04-15 | Montero-Julian Felix A. | Methods and systems for detecting MHC class I binding peptides |
| CN1791679A (zh) | 2003-03-19 | 2006-06-21 | 伊索格尼斯股份有限公司 | 基于CD8α链的具有降低的免疫原性的基因治疗载体 |
| SE0301109D0 (sv) * | 2003-04-14 | 2003-04-14 | Mallen Huang | Nucleotide vaccine composition |
| US7663017B2 (en) | 2003-07-30 | 2010-02-16 | Institut Pasteur | Transgenic mice having a human major histocompatability complex (MHC) phenotype, experimental uses and applications |
| DE10347710B4 (de) | 2003-10-14 | 2006-03-30 | Johannes-Gutenberg-Universität Mainz | Rekombinante Impfstoffe und deren Verwendung |
| WO2005085438A1 (ja) | 2004-03-09 | 2005-09-15 | Nagoya Industrial Science Research Institute | ヒト関節リウマチの病態を再現するトランスジェニック非ヒト哺乳動物 |
| DK1802193T3 (da) | 2004-10-19 | 2014-06-10 | Regeneron Pharma | Fremgangsmåde til frembringelse af en homozygot mus til genetisk modifikation |
| CA2651174A1 (en) | 2006-05-03 | 2007-11-15 | Government Of The United States Of America, Represented By The Secretary , Department Of Health And Human Services | Chimeric t cell receptors and related materials and methods of use |
| EP1878342A1 (en) | 2006-07-13 | 2008-01-16 | Institut Pasteur | Immunodeficient mice transgenic for HLA class I and HLA class II molecules and their uses |
| EP1878798A1 (en) | 2006-07-13 | 2008-01-16 | Institut Pasteur | Method of producing a multichimeric mouse and applications to study the immunopathogenesis of human tissue-specific pathologies |
| HU0700534D0 (en) * | 2006-11-24 | 2007-10-29 | Mezoegazdasagi Biotechnologiai | Transgenic animal with enhanced immune response and method for the preparation thereof |
| RU2448979C2 (ru) | 2006-12-14 | 2012-04-27 | Ридженерон Фармасьютикалз, Инк. | Антитела человека к дельта-подобному лиганду-4 человека |
| JP2008159109A (ja) | 2006-12-21 | 2008-07-10 | Matsushita Electric Ind Co Ltd | データ転送装置 |
| US20090271884A1 (en) | 2008-03-07 | 2009-10-29 | Regeneron Pharmaceuticals, Inc. | ES Cell-Derived Mice From Diploid Host Embryo Injection |
| UA121453C2 (uk) | 2008-04-11 | 2020-06-10 | Чугей Сейяку Кабусікі Кайся | Спосіб одержання фармацевтичної композиції, яка містить антитіло |
| US20090328240A1 (en) | 2008-06-24 | 2009-12-31 | Sing George L | Genetically modified mice as predictors of immune response |
| PL2346994T3 (pl) | 2008-09-30 | 2022-04-19 | Ablexis, Llc | Myszy knock-in do wytwarzania chimerycznych przeciwciał |
| DK2564695T3 (en) | 2009-07-08 | 2015-05-26 | Kymab Ltd | Animal models and therapeutic molecules |
| GB2472108B (en) | 2009-08-27 | 2011-07-13 | Budha Singh Dhinjan | Wall bead |
| PL2505654T5 (pl) | 2010-02-08 | 2020-11-30 | Regeneron Pharmaceuticals, Inc. | Mysz ze wspólnym łańcuchem lekkim |
| KR20150002894A (ko) | 2010-03-11 | 2015-01-07 | 리나트 뉴로사이언스 코프. | pH 의존성 항원 결합을 갖는 항체 |
| PL2553100T3 (pl) | 2010-03-31 | 2018-03-30 | Ablexis, Llc | Inżynieria genetyczna zwierząt nie będących ludźmi w celu wytworzenia przeciwciał chimerycznych |
| JP2013538555A (ja) | 2010-07-15 | 2013-10-17 | テクニオン リサーチ アンド ディベロップメント ファウンデーション リミテッド | Mhcクラスiiとグルタミン酸デカルボキシラーゼ(gad)自己抗原性ペプチドとの天然型複合体に対するt細胞受容体様特異性を有する単離された高親和性実体 |
| US10881084B2 (en) | 2010-07-26 | 2021-01-05 | Trianni, Inc | Transgenic animals and methods of use |
| RS56656B1 (sr) * | 2011-10-28 | 2018-03-30 | Regeneron Pharma | Miševi sa genetski modifikovanim glavnim kompleksom gena tkivne podudarnosti |
| CA2852962C (en) | 2011-10-28 | 2022-05-03 | Regeneron Pharmaceuticals, Inc. | Genetically modified mice expressing chimeric major histocompatibility complex (mhc) ii molecules |
| US9043996B2 (en) | 2011-10-28 | 2015-06-02 | Regeneron Pharmaceuticals, Inc. | Genetically modified major histocompatibility complex animals |
| RU2634417C2 (ru) | 2011-10-28 | 2017-10-26 | Регенерон Фармасьютикалс, Инк. | Гуманизированные il-6 и рецептор il-6 |
| US9591835B2 (en) | 2011-10-28 | 2017-03-14 | Regeneron Pharmaceuticals, Inc. | Genetically modified major histocompatibility complex animals |
| EP3939423B1 (en) | 2012-11-05 | 2024-03-06 | Regeneron Pharmaceuticals, Inc. | Genetically modified non-human animals and methods of use thereof |
| PT2958938T (pt) | 2013-02-20 | 2019-07-23 | Regeneron Pharma | Ratinhos que expressam correcetores de células t humanizados |
| CN110192541B (zh) * | 2013-02-22 | 2022-02-18 | 瑞泽恩制药公司 | 表达人源化主要组织相容性复合物的小鼠 |
| US20150342163A1 (en) | 2013-02-22 | 2015-12-03 | Regeneron Pharmaceuticals, Inc. | Genetically modified major histocompatibility complex mice |
| CA2903025C (en) | 2013-03-11 | 2025-10-07 | Regeneron Pharmaceuticals, Inc. | TRANSGENIC MICE EXPRESSING CHIMERIC MAJOR HISTOCOMPATIBILITY COMPLEX (MHC) CLASS II MOLECULES |
| ES2764829T3 (es) | 2013-03-11 | 2020-06-04 | Regeneron Pharma | Ratones transgénicos que expresan moléculas quiméricas del complejo mayor de histocompatibilidad (mhc) de clase I |
| BR112015030211A2 (pt) | 2013-06-03 | 2017-08-22 | Theravectys | Vetores lentivirais contendo uma sequência promotora a montante da mhc de classe i, mhc de classe ii ou microglobulina beta-2 |
| CN105578876B (zh) | 2013-10-18 | 2018-07-13 | 大鹏药品工业株式会社 | 制备表达hlai类的非人动物的方法 |
| WO2016164492A2 (en) | 2015-04-06 | 2016-10-13 | Regeneron Pharmaceuticals, Inc. | Humanized t cell mediated immune responses in non-human animals |
| AU2017272337C1 (en) | 2016-06-03 | 2024-02-29 | Regeneron Pharmaceuticals, Inc. | Non-human animals expressing exogenous terminal deoxynucleotidyltransferase |
| PT3720279T (pt) | 2017-12-05 | 2022-10-06 | Regeneron Pharma | Ratinhos tendo uma cadeia leve lambda de imunoglobulina manipulada e seus usos |
-
2014
- 2014-02-20 CN CN201910379017.5A patent/CN110192541B/zh active Active
- 2014-02-20 CN CN201480022254.1A patent/CN105164154B/zh active Active
- 2014-02-20 LT LTEP14708429.7T patent/LT2958937T/lt unknown
- 2014-02-20 JP JP2015558950A patent/JP6444321B2/ja active Active
- 2014-02-20 EP EP14708429.7A patent/EP2958937B1/en active Active
- 2014-02-20 PL PL14708429T patent/PL2958937T3/pl unknown
- 2014-02-20 WO PCT/US2014/017387 patent/WO2014130667A1/en not_active Ceased
- 2014-02-20 KR KR1020157026032A patent/KR102211267B1/ko active Active
- 2014-02-20 SI SI201430965T patent/SI2958937T1/sl unknown
- 2014-02-20 PT PT14708429T patent/PT2958937T/pt unknown
- 2014-02-20 RS RS20181333A patent/RS57963B1/sr unknown
- 2014-02-20 ES ES20163735T patent/ES2949029T3/es active Active
- 2014-02-20 ES ES14708429T patent/ES2697248T3/es active Active
- 2014-02-20 EP EP20163735.2A patent/EP3699190B9/en active Active
- 2014-02-20 AU AU2014218893A patent/AU2014218893C1/en active Active
- 2014-02-20 SM SM20180614T patent/SMT201800614T1/it unknown
- 2014-02-20 DK DK14708429.7T patent/DK2958937T3/en active
- 2014-02-20 CA CA2900824A patent/CA2900824A1/en active Pending
- 2014-02-20 HR HRP20181866TT patent/HRP20181866T1/hr unknown
- 2014-02-20 ES ES18182319T patent/ES2798751T3/es active Active
- 2014-02-20 HU HUE14708429A patent/HUE041878T2/hu unknown
- 2014-02-20 US US14/185,316 patent/US10154658B2/en active Active
- 2014-02-20 EP EP18182319.6A patent/EP3421490B1/en active Active
-
2018
- 2018-05-28 JP JP2018101501A patent/JP6681939B2/ja active Active
- 2018-11-08 US US16/184,473 patent/US11224208B2/en active Active
- 2018-11-13 CY CY20181101195T patent/CY1121223T1/el unknown
- 2018-11-29 AU AU2018271326A patent/AU2018271326C1/en active Active
-
2021
- 2021-12-14 US US17/550,945 patent/US12414551B2/en active Active
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5523226A (en) * | 1993-05-14 | 1996-06-04 | Biotechnology Research And Development Corp. | Transgenic swine compositions and methods |
| CN101060776A (zh) * | 2004-11-23 | 2007-10-24 | 安迪拓普有限公司 | Mhc-ⅱ转基因动物耐受性的研究方法 |
| CN102711449A (zh) * | 2009-12-10 | 2012-10-03 | 瑞泽恩制药公司 | 生产重链抗体的小鼠 |
| WO2012039779A1 (en) * | 2010-09-24 | 2012-03-29 | The United States Of America As Represented By The Secretary Of The Navy | Humanized transgenic mouse model |
Non-Patent Citations (1)
| Title |
|---|
| MHC Class-1 transgenic mice;B Arnold等;《Annual Review of Immunology》;19910101;第297-322页 * |
Also Published As
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US12414551B2 (en) | Genetically modified major histocompatibility complex mice | |
| US10314296B2 (en) | Genetically modified major histocompatibility complex mice | |
| CN108401986B (zh) | 表达嵌合主要组织相容性复合物(mhc)ii类分子的基因修饰小鼠 | |
| CN105208855B (zh) | 表达嵌合的主要组织相容性复合物(mhc)ii类分子的转基因小鼠 | |
| WO2014130671A1 (en) | Mice expressing humanized t-cell co-receptors | |
| HK40036775A (zh) | 表达人源化主要组织相容性复合物的小鼠细胞 | |
| HK40036775B (zh) | 表达人源化主要组织相容性复合物的小鼠细胞 | |
| RU2797549C2 (ru) | Генетически модифицированные мыши, экспрессирующие химерные молекулы главного комплекса гистосовместимости | |
| HK40007414B (zh) | 产生表达人类主要组织相容性复合物的非人类动物的方法 | |
| HK40007414A (zh) | 产生表达人类主要组织相容性复合物的非人类动物的方法 | |
| HK1214613B (zh) | 表達人源化主要組織相容性複合物的小鼠 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| GR01 | Patent grant | ||
| GR01 | Patent grant |