CN102388134A - 具有β-葡糖苷酶活性的多肽和编码该多肽的多核苷酸 - Google Patents
具有β-葡糖苷酶活性的多肽和编码该多肽的多核苷酸 Download PDFInfo
- Publication number
- CN102388134A CN102388134A CN2010800153740A CN201080015374A CN102388134A CN 102388134 A CN102388134 A CN 102388134A CN 2010800153740 A CN2010800153740 A CN 2010800153740A CN 201080015374 A CN201080015374 A CN 201080015374A CN 102388134 A CN102388134 A CN 102388134A
- Authority
- CN
- China
- Prior art keywords
- polypeptide
- seq
- sequence
- polynucleotide
- cell
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images

Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P21/00—Preparation of peptides or proteins
- C12P21/02—Preparation of peptides or proteins having a known sequence of two or more amino acids, e.g. glutathione
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/24—Hydrolases (3) acting on glycosyl compounds (3.2)
- C12N9/2402—Hydrolases (3) acting on glycosyl compounds (3.2) hydrolysing O- and S- glycosyl compounds (3.2.1)
- C12N9/2405—Glucanases
- C12N9/2434—Glucanases acting on beta-1,4-glucosidic bonds
- C12N9/2445—Beta-glucosidase (3.2.1.21)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/14—Preparation of compounds containing saccharide radicals produced by the action of a carbohydrase (EC 3.2.x), e.g. by alpha-amylase, e.g. by cellulase, hemicellulase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P7/00—Preparation of oxygen-containing organic compounds
- C12P7/02—Preparation of oxygen-containing organic compounds containing a hydroxy group
- C12P7/04—Preparation of oxygen-containing organic compounds containing a hydroxy group acyclic
- C12P7/06—Ethanol, i.e. non-beverage
- C12P7/08—Ethanol, i.e. non-beverage produced as by-product or from waste or cellulosic material substrate
- C12P7/10—Ethanol, i.e. non-beverage produced as by-product or from waste or cellulosic material substrate substrate containing cellulosic material
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y302/00—Hydrolases acting on glycosyl compounds, i.e. glycosylases (3.2)
- C12Y302/01—Glycosidases, i.e. enzymes hydrolysing O- and S-glycosyl compounds (3.2.1)
- C12Y302/01021—Beta-glucosidase (3.2.1.21)
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02E—REDUCTION OF GREENHOUSE GAS [GHG] EMISSIONS, RELATED TO ENERGY GENERATION, TRANSMISSION OR DISTRIBUTION
- Y02E50/00—Technologies for the production of fuel of non-fossil origin
- Y02E50/10—Biofuels, e.g. bio-diesel
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Health & Medical Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Genetics & Genomics (AREA)
- General Health & Medical Sciences (AREA)
- General Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Biomedical Technology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Enzymes And Modification Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
Abstract
本发明涉及具有β-葡糖苷酶活性的分离的多肽和编码所述多肽的分离的多核苷酸。本发明还涉及包含所述多核苷酸的核酸构建体、载体和宿主细胞,以及用于制备和使用所述多肽的方法。
Description
涉及序列表
本申请通过EFS电子提交的序列表,其通过提述并入本文。
涉及生物材料的保藏
本申请包含对于生物材料保藏的引用,所述保藏通过提述并入本文。
发明背景
发明领域
本发明涉及具有β-葡糖苷酶活性的分离的多肽和编码所述多肽的分离的多核苷酸。本发明还涉及包含所述多核苷酸的核酸构建体、载体和宿主细胞,以及用于产生和使用所述多肽的方法。
相关领域描述
纤维素是葡萄糖通过β-1,4-键连接的聚合物。许多微生物产生水解β-连接的葡聚糖的酶。这些酶包括内切葡聚糖酶、纤维二糖水解酶和β-葡糖苷酶。内切葡聚糖酶在随机位置消化纤维素聚合物,将其打开(opening it)以受到纤维二糖水解酶攻击(attack)。纤维二糖水解酶继而从纤维素聚合物的末端释放纤维二糖的分子。纤维二糖是水溶性的β-1,4-连接的葡萄糖二聚体。β-葡糖苷酶将纤维二糖水解成葡萄糖。
将含木素纤维素原料(lignocellulosic feedstock)转化为乙醇具有以下优势:大量原料现成可用,避免燃烧或填埋材料的合意性和乙醇燃料的清洁性。木材、农业残余物、草本作物和城市固体废物被认为是用于乙醇生产的原料。这些材料主要由纤维素、半纤维素和木质素组成。一旦将纤维素转化成葡萄糖,葡萄糖将容易地由酵母发酵成乙醇。因为葡萄糖容易地由多种酵母发酵为乙醇,而纤维二糖则否,所以任何在水解结束时残留的纤维二糖代表乙醇产量的损失。更重要的是,纤维二糖是内切葡聚糖酶和纤维二糖水解酶的强力抑制剂。在水解过程中纤维二糖的累积对于乙醇产生是不受欢迎的。
纤维二糖累积在酶水解中为主要问题,因为产生纤维素酶的微生物可能产生极少的β-葡糖苷酶。该β-葡糖苷酶的低量导致将纤维二糖水解为葡萄糖的能力的短缺。已使用了几种方法在纤维素转化中增加β-葡糖苷酶的量。
一种方法是使用产生极少纤维素酶的微生物产生β-葡糖苷酶,并将所述β-葡糖苷酶外源地添加至内切葡聚糖酶和纤维二糖水解酶以增强水解。
第二种方法是将纤维素水解与发酵同时进行。该工艺称作同时糖化和发酵(SFF)。在SFF系统中,葡萄糖的发酵将其从溶液移除。
第三种克服β-葡糖苷酶的短缺的方法是在宿主中过表达β-葡糖苷酶,由此增加β-葡糖苷酶的产量。
在本领域提供用于降解纤维素材料的新β-葡糖苷酶会是有利的。
本发明提供了具有β-葡糖苷酶活性的多肽和编码该多肽的多核苷酸。
发明概述
本发明涉及具有β-葡糖苷酶活性的分离的多肽,所述多肽选自下组:
(a)多肽,其包含氨基酸序列,所述氨基酸序列与SEQ ID NO:2的成熟多肽具有至少60%序列同一性;
(b)多肽,其由多核苷酸编码,所述多核苷酸在至少中严格条件下与以下杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链;
(c)多肽,其由包含核苷酸序列的多核苷酸编码,所述核苷酸序列与SEQID NO:1的成熟多肽编码序列具有至少60%序列同一性;和
(d)SEQ ID NO:2的包含取代、缺失和/或插入一个或多个(几个)氨基酸的成熟多肽的变体。
本发明还涉及编码具有β-葡糖苷酶活性的多肽的分离的多核苷酸,所述多核苷酸选自下组:
(a)多核苷酸,其编码包含氨基酸序列的多肽,所述氨基酸序列与SEQ IDNO:2的成熟多肽具有至少60%序列同一性;
(b)多核苷酸,其在至少中严格条件下与以下杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列或(iii)(i)或(ii)的全长互补链;
(c)多核苷酸,其包含核苷酸序列,所述核苷酸序列与SEQ ID NO:1的成熟多肽编码序列具有至少60%序列同一性;和
(d)多核苷酸,其编码SEQ ID NO:2的成熟多肽的包含取代、缺失和/或插入一个或多个(几个)氨基酸的变体。
本发明还涉及包含所述多核苷酸的核酸构建体、重组表达载体和重组宿主细胞,并涉及产生具有β-葡糖苷酶活性的多肽的方法。
本发明还涉及抑制具有β-葡糖苷酶活性的多肽在细胞中表达的方法,所述方法包括:对细胞施用或在细胞中表达双链RNA(dsRNA)分子,其中所述dsRNA包括本发明的多核苷酸的亚序列。本发明还涉及这种双链抑制性RNA(dsRNA)分子,其中任选地dsRNA是siRNA或miRNA分子。
本发明还涉及用于降解或转化纤维素材料的方法,其包括:在存在具有β-葡糖苷酶活性的多肽的情况下,用酶组合物处理纤维素材料。
本发明还涉及产生发酵产物的方法,其包括:(a)在存在具有β-葡糖苷酶活性的多肽的情况下用酶组合物糖化纤维素材料;(b)用一种或多种(几种)发酵微生物发酵经糖化的纤维素材料以产生发酵产物;和(c)从发酵回收发酵产物。
本发明还涉及发酵纤维素材料的方法,其包括:用一种或多种(几种)发酵微生物发酵纤维素材料,其中在存在具有β-葡糖苷酶活性的多肽的情况下,用酶组合物糖化纤维素材料。
本发明还涉及包含分离的多核苷酸的植物,所述多核苷酸编码具有β-葡糖苷酶活性的多肽。
本发明还涉及用于产生具有β-葡糖苷酶活性的多肽的方法,其包括:(a)在有助于产生该多肽的条件下培养包含多核苷酸的转基因植物或植物细胞,所述多核苷酸编码具有β-葡糖苷酶活性的多肽;和(b)回收所述多肽。
本发明还涉及编码信号肽的分离的多核苷酸,所述信号肽包含SEQ IDNO:2的氨基酸1至19,或由SEQ ID NO:2的氨基酸1至19组成;涉及包含所述多核苷酸的核酸构建体、表达载体和重组宿主细胞;并涉及产生蛋白的方法。
附图简述
图1A和1B显示Trichophaea saccata CBS 804.70β-葡糖苷酶基因的cDNA序列和推定的氨基酸序列(分别为SEQ ID NO:1和2)。
图2显示pAHYG-33的限制图谱。
定义
β-葡糖苷酶:术语“β-葡糖苷酶”在本文中定义为β-D-葡糖苷葡糖水解酶(beta-D-glucoside glucohydrolase)(E.C.No.3.2.1.21),其催化末端非还原β-D-葡萄糖残基的水解,并释放β-D-葡萄糖。就本发明而言,根据Venturi等,2002,Extracellular beta-D-glucosidase from Chaetomium thermophilum var.coprophilum:production,purification and some biochemical properties,J.Basic Microbiol.42:55-66描述的基本方法确定β-葡糖苷酶活性。一个单位的β-葡糖苷酶活性定义为在50℃,pH 5,在50mM乙酸钠,0.01%20中从作为底物的1mM对硝基苯基-β-D-葡糖吡喃糖苷每分钟产生1.0微摩尔对硝基苯酚。
本发明的多肽具有SEQ ID NO:2的成熟多肽的β-葡糖苷酶活性的优选至少20%,更优选至少40%,更优选至少50%,更优选至少60%,更优选至少70%,更优选至少80%,甚至更优选至少90%,最优选至少95%,且甚至最优选至少100%。
家族3或家族GH3或CEL3:术语“家族3”或“家族GH3”或“CEL3”在本文中定义为根据Henrissat B.,1991,A classification of glycosyl hydrolases basedon amino-acid sequence similarities,Biochem.J.280:309-316,及Henrissat B.,和Bairoch A.,1996,Updating the sequence-based classification of glycosylhydrolases,Biochem.J.316:695-696属于糖苷水解酶家族3的多肽。
纤维素分解活性:术语“纤维素分解活性”在本文中定义为水解纤维素材料的生物学活性。测量纤维素分解活性的两种基本方法包括:(1)测量总纤维素分解活性,和(2)测量单独的纤维素分解活性(内切葡聚糖酶、纤维二糖水解酶和β-葡糖苷酶),如Zhang等,Outlook for cellulase improvement:Screening and selectionstrategies,2006,Biotechnology Advances 24:452-481所综述的。总纤维素分解活性通常是使用不溶性底物来测定的,所述底物包括Whatman No.1滤纸、微晶纤维素、细菌纤维素、藻类纤维素、棉花、经预处理的木素纤维素等。最常见的总纤维素分解活性测定法是使用Whatman No.1滤纸作为底物的滤纸测定法。该测定法是由International Union of Pure and Applied Chemistry(IUPAC)(Ghose,1987,Measurement of cellulase activities,Pure Appl.Chem.59:257-68)确立的。
就本发明而言,纤维素分解活性通过测量在下述条件下由纤维素分解酶进行的纤维素材料水解的增加来确定:1-20mg的纤维素分解蛋白/g的PCS中纤维素在50-65℃进行3-7日,与未添加纤维素分解蛋白的对照水解相比较。通常条件为:1ml反应液,经洗涤或未洗涤的PCS,5%不溶性固形物,50mM乙酸钠pH 5,1mM MnSO4,50-65℃,72小时,通过 HPX-87H柱(Bio-Rad Laboratories,Inc.,Hercules,CA,USA)进行糖分析。
HPX-87H柱(Bio-Rad Laboratories,Inc.,Hercules,CA,USA)进行糖分析。
内切葡聚糖酶:术语“内切葡聚糖酶”在本文中定义为内切-1,4-(1,3;1,4)-β-D-葡聚糖4-葡聚糖水解酶(endo-1,4-β-D-glucan 4-glucanohydrolase)(E.C.3.2.1.4),其催化纤维素、纤维素衍生物(例如羧甲基纤维素和羟乙基纤维素)、地衣淀粉(lichenin)中的1,4-β-D-糖苷键、混合的β-1,3葡聚糖例如谷类β-D-葡聚糖或木葡聚糖和含有纤维素组分的其它植物材料中的β-1,4键的内水解(endohydrolysis)。内切葡聚糖酶活性可基于底物粘度的减少或由还原糖测定法(Zhang等,2006,Biotechnology Advances 24:452-481)确定的还原端增加来确定。就本发明而言,根据Ghose,1987,Pure and Appl.Chem.59:257-268的方法,使用羧甲基纤维素(CMC)水解来确定内切葡聚糖酶活性。
纤维二糖水解酶:术语“纤维二糖水解酶”在本文中定义为1,4-β-D-葡聚糖纤维二糖水解酶(1,4-beta-D-glucan cellobiohydrolase)(E.C.No.3.2.1.91),其催化纤维素、纤维素寡糖,或任何包含β-1,4-连接的葡萄糖的聚合物中的1,4-β-D-糖苷键的水解,从链的还原或非还原末端释放纤维二糖(Teeri,1997,Crystallinecellulose degradation:New insight into the function of cellobiohydrolases,Trends inBiotechnology 15:160-167;Teeri等,1998,Trichoderma reesei cellobiohydrolases:why so efficient on crystalline cellulose?,Biochem.Soc.Trans.26:173-178)。就本发明而言,根据van Tilbeurgh等,1982,FEBSLetters 149:152-156和van Tilbeurgh和Claeyssens,1985,FEBS Letters 187:283-288描述的方法使用荧光性二糖衍生物4-甲基伞形基-β-D-乳糖苷来测定纤维二糖水解酶活性。
纤维素分解增强活性:术语“纤维素分解增强活性”在本文中定义为增强具有纤维素分解活性的多肽水解纤维素材料的生物学活性。就本发明而言,通过测量由纤维素分解蛋白在下述条件下水解纤维素材料所导致的还原糖增加或纤维二糖与葡萄糖的总量增加来确定纤维素分解增强活性:1-50mg总蛋白/gPCS中的纤维素,其中总蛋白包含50-99.5%w/w的纤维素分解蛋白,及0.5-50%w/w的具有纤维素分解增强活性的蛋白质,在50-65℃历时1-7天,与用等量的总蛋白加载量而无纤维素分解增强活性(1-50mg纤维素分解蛋白/g PCS中的纤维素)所进行的对照水解相比。在一个优选的方面,使用在总蛋白重量的3%的米曲霉β-葡糖苷酶(根据WO 02/095014在米曲霉中重组产生)或者总蛋白质量的3%的烟曲霉β-葡糖苷酶(如WO 2002/095014所述在米曲霉中重组产生)的纤维素酶蛋白加载量存在下的 1.5L(Novozymes A/S,Bagsvaerd,Denmark)的混合物作为纤维素分解活性的来源。
1.5L(Novozymes A/S,Bagsvaerd,Denmark)的混合物作为纤维素分解活性的来源。
本发明的具有纤维素分解增强活性的多肽具有GH61多肽的成熟多肽的纤维素分解增强活性的优选至少20%,更优选至少40%,更优选至少50%,更优选至少60%,更优选至少70%,更优选至少80%,甚至更优选至少90%,最优选至少95%,并且甚至最优选至少100%。
具有纤维素分解增强活性的多肽通过降低达到相同水解水平所需的纤维素分解酶的量而增强由具有纤维素分解活性的蛋白质催化的纤维素材料的水解,优选降低至少1.01倍,更优选至少1.05倍,更优选至少1.10倍,更优选至少1.25倍,更优选至少1.5倍,更优选至少2倍,更优选至少3倍,更优选至少4倍,更优选至少5倍,甚至更优选至少10倍,并且最优选至少20倍。
家族61糖苷水解酶:术语“家族61糖苷水解酶”或“家族GH61”在本文中定义为根据Henrissat B.,1991,A classification of glycosyl hydrolases based onamino-acid sequence similarities,Biochem.J.280:309-316,及Henrissat B.,和Bairoch A.,1996,Updating the sequence-based classification of glycosylhydrolases,Biochem.J.316:695-696属于糖苷水解酶家族61的多肽。目前,Henrissat将GH61家族列为未分类的,表明尚未知属于该家族的多肽的性质如机理,催化亲核体/碱和催化质子供体。
木聚糖降解活性:术语“木聚糖降解活性”或“木聚糖分解活性”在本文中定义为水解含木聚糖材料的生物学活性。两种测定木聚糖分解活性的基础方法包括:(1)测定总木聚糖分解活性,和(2)测定单独的木聚糖分解活性(内切木聚糖酶、β-木糖苷酶、阿拉伯呋喃糖苷酶、α-葡糖醛酸糖苷酶、乙酰木聚糖酯酶、阿魏酸酯酶和α-葡糖醛酸酯酶(α-glucuronyl esterase))。最近在木聚糖分解酶测定法的进展总结于几个公开文献中,包括Biely和Puchard,Recentprogress in the assays of xylanolytic enzymes,2006,Journal of the Science ofFood and Agriculture 86(11):1636-1647;Spanikova和Biely,2006,Glucuronoylesterase-Novel carbohydrate esterase produced by Schizophyllum commune,FEBS Letters 580(19):4597-4601;Herrmann,Vrsanska,Jurickova,Hirsch,Biely和Kubicek,1997,The beta-D-xylosidase of Trichoderma reesei is amultifunctional beta-D-xylan xylohydrolase,Biochemical Journal 321:375-381。
总木聚糖降解活性可通过确定从多种类型的木聚糖形成的还原糖来测量,所述木聚糖包括燕麦小麦(oat spelt)、山毛榉木(beechwood)和落叶松木(larchwood)木聚糖,或者可通过光度法确定从多种共价染色的木聚糖释放出的染色的木聚糖片段来测量。最常见的总木聚糖分解活性测定法基于从多聚的4-O-甲基葡糖醛酸木聚糖产生还原糖,如Bailey,Biely,Poutanen,1992,Interlaboratory testing of methods for assay of xylanase activity,Journal ofBiotechnology 23(3):257-270中所述。
就本发明而言,木聚糖降解活性是通过测量由木聚糖降解酶在下述通常条件下造成的桦木木聚糖(Sigma Chemical Co.,Inc.,St.Louis,MO,USA)水解的增加来确定的:1ml反应液,5mg/ml底物(总固形物),5mg木聚糖分解蛋白质/g底物,50mM乙酸钠,pH 5,50℃,24小时,如Lever,1972,A new reactionfor colorimetric determination of carbohydrates,Anal.Biochem 47:273-279所述使用对羟基苯甲酸酰肼(PHBAH)测定法进行糖分析。
木聚糖酶活性:术语“木聚糖酶活性”在本文中定义为1,4-β-D-木聚糖-木糖水解酶(1,4-β-D-xylan-xylohydrolase)活性(E.C.3.2.1.8),其催化木聚糖中1,4-β-D-木糖苷键的内水解。就本发明而言,木聚糖酶活性是使用桦木木聚糖作为底物确定的。一个单位的木聚糖酶活性定义为在50℃,pH 5从每升2g桦木木聚糖作为底物在含有0.01% 20的50mM乙酸钠中水解的起始时间过程中每分钟产生1.0μmol还原糖(以葡萄糖当量(glucose equivalents)测定,如Lever,1972,A new reaction for colorimetric determination ofcarbohydrates,Anal.Biochem 47:273-279所述)。
20的50mM乙酸钠中水解的起始时间过程中每分钟产生1.0μmol还原糖(以葡萄糖当量(glucose equivalents)测定,如Lever,1972,A new reaction for colorimetric determination ofcarbohydrates,Anal.Biochem 47:273-279所述)。
β-木糖苷酶活性:术语“β-木糖苷酶活性”在本文中定义为β-D木糖苷木糖水解酶(β-D-xyloside xylohydrolase)(E.C.3.2.1.37),其催化短β(1→4)木寡糖(xylooligosaccharide)的外水解以从非还原端去除连续的D-木糖残基。就本发明而言,一个单位的β-木糖苷酶活性定义为在40℃,pH 5从1mM对硝基苯基-β-D-木糖苷作为底物在含有0.01% 20的100mM柠檬酸钠中每分钟产生1.0μmol对硝基苯酚。
20的100mM柠檬酸钠中每分钟产生1.0μmol对硝基苯酚。
乙酰木聚糖酯酶活性:术语“乙酰木聚糖酯酶活性”在本文中定义为羧基酯酶活性(EC 3.1.1.72),其催化乙酰基从聚合木聚糖、乙酰化木糖、乙酰化葡萄糖、乙酸α-萘酯(alpha-napthyl acetate)和乙酸对硝基苯酯(p-nitrophenylacetate)的水解。就本发明而言,乙酰木聚糖酯酶活性是使用0.5mM乙酸对硝基苯酯作为底物,在含有0.01%TWEENTM 20的50mM乙酸钠pH 5.0中确定的。一个单位的乙酰木聚糖酯酶活性定义为能够在pH 5,25℃每分钟释放1微摩尔对硝基苯酚阴离子(p-nitrophenolate anion)的酶量。
阿魏酸酯酶活性:术语“阿魏酸酯酶(feruloyl esterase)活性”在本文中定义为4-羟基-3-甲氧基肉桂酰-糖水解酶酶活性(EC 3.1.1.73),其催化4-羟基-3-甲氧基肉桂酰(阿魏酰)基团从酯化的糖(其在“天然”底物中通常为阿拉伯糖)的水解,以产生阿魏酸(4-羟基-3-甲氧基肉桂酸)。阿魏酸酯酶也称作阿魏酸酯酶(ferulic acid esterase)、FAE-III、肉桂酸酯水解酶、FAEA、cinnAE、FAE-I或FAE-II。就本发明而言,阿魏酸酯酶活性是使用50mM乙酸钠pH 5.0中的0.5mM阿魏酸对硝基苯酯作为底物确定的。一个单位的阿魏酸酯酶活性等于能够在pH 5,25℃每分钟释放出1μmol对硝基苯酚阴离子的酶量。
α-葡糖醛酸糖苷酶活性:术语“α-葡糖醛酸糖苷酶活性”在本文中定义为α-D-葡糖苷酸葡糖醛酸水解酶活性(alpha-D-glucosiduronateglucuronohydrolase activity)(EC 3.2.1.139),其催化α-D-葡糖醛酸糖苷水解为D-葡糖醛酸和醇。就本发明而言,α-葡糖醛酸糖苷酶活性是根据de Vries,1998,J.Bacteriol.180:243-249确定的。一个单位的α-葡糖醛酸糖苷酶活性等于能够在pH 5,40℃每分钟释放出1μmol葡糖醛酸或4-O-甲基葡糖醛酸的酶量。
α-L-阿拉伯呋喃糖苷酶活性:术语“α-L-阿拉伯呋喃糖苷酶活性”在本文中定义为α-L-阿拉伯呋喃糖苷阿拉伯呋喃水解酶活性(EC 3.2.1.55),其催化对α-L-阿拉伯糖苷中的末端非还原性α-L-阿拉伯呋喃糖苷残基的水解。该酶活性对α-L-阿拉伯呋喃糖苷、含有(1,3)-和/或(1,5)-键的α-L-阿拉伯聚糖、阿拉伯木聚糖和阿拉伯半乳聚糖起作用。α-L-阿拉伯呋喃糖苷酶也称为阿拉伯糖苷酶、α-阿拉伯糖苷酶、α-L-阿拉伯糖苷酶、α-阿拉伯呋喃糖苷酶、多糖α-L-阿拉伯呋喃糖苷酶、α-L-阿拉伯呋喃糖苷水解酶、L-阿拉伯糖苷酶或α-L-阿拉伯聚糖酶。就本发明而言,α-L-阿拉伯呋喃糖苷酶活性是使用总体积200μl中的每ml的100mM乙酸钠pH 5中5mg的中等粘性小麦阿拉伯木聚糖(Megazyme International Ireland,Ltd.,Bray,Co.Wicklow,Ireland)在40℃进行30分钟,接着通过 HPX-87H柱层析(Bio-Rad Laboratories,Inc.,Hercules,CA,USA)的阿拉伯糖分析来确定的。
HPX-87H柱层析(Bio-Rad Laboratories,Inc.,Hercules,CA,USA)的阿拉伯糖分析来确定的。
纤维素材料:纤维素材料可以是包含纤维素的任何材料。生物质的初生细胞壁(primary cell wall)中的主要多糖是纤维素,其次最丰富的是半纤维素,而第三是果胶。次生细胞壁(secondary cell wall)在细胞停止生长后产生,其同样含有多糖并通过共价交联至半纤维素的聚合木质素而加强。纤维素是脱水纤维二糖的均聚物,并且因此是直链β-(1-4)-D-葡聚糖,而半纤维素包括多种化合物,例如木聚糖、木葡聚糖(xyloglucan)、阿拉伯木聚糖和甘露聚糖,具有系列取代基的复杂分支结构。尽管通常是多形的,存在于植物组织中的纤维素主要是平行葡聚糖链的不溶晶体基质。半纤维素通常与纤维素以及其它半纤维素以氢键相连,其帮助稳定细胞壁基质。
纤维素通常见于例如植物的茎、叶、壳、皮和穗轴,或树的叶、枝和木材。纤维素材料可以是,但不限于,草本材料、农业残余物、林业残余物、城市固体废物、废纸和纸浆与造纸厂残余物(参见,例如,Wiselogel等,1995,于Handbook on Bioethanol(Charles E.Wyman编),pp.105-118,Taylor & Francis,Washington D.C.;Wyman,1994,Bioresource Technology 50:3-16;Lynd,1990,Applied Biochemistry and Biotechnology 24/25:695-719;Mosier等,,1999,Recent Progress in Bioconversion of Lignocellulosics,于Advances inBiochemical Engineering/Biotechnology,T.Scheper主编,Volume 65,pp.23-40,Springer-Verlag,New York)。在本文中应理解的是,纤维素可以是任何形式的木素纤维素,在混合基质中包含木质素、纤维素和半纤维素的植物细胞壁材料。在一个优选的方面,纤维素材料是木素纤维素。
在一个方面,纤维素材料是草本材料。在另一个方面,纤维素材料是农业残余物。在另一个方面,纤维素材料是林业残余物。在另一个方面,纤维素材料是城市固体废物。在另一个方面,纤维素材料是废纸。在另一个方面,纤维素材料是纸浆和造纸厂残余物。
在另一个方面,纤维素材料是玉米秸秆。在另一个方面,纤维素材料是玉米纤维。在另一个方面,纤维素材料是玉米穗轴。在另一个方面,纤维素材料是橙皮。在另一个方面,纤维素材料是稻杆。在另一个方面,纤维素材料是麦杆。在另一个方面,纤维素材料是柳枝稷(switch grass)。在另一个方面,纤维素材料是芒草属。在另一个方面,纤维素材料是甘蔗渣。
在另一个方面,纤维素材料是微晶纤维素。在另一个方面,纤维素材料是细菌纤维素。在另一个方面,纤维素材料是藻类纤维素。在另一个方面,纤维素材料是棉线头(cotton linter)。在另一个方面,纤维素材料是无定形的磷酸处理的纤维素。在另一个方面,纤维素材料是滤纸。
纤维素材料可以直接使用或进行预处理,使用本领域已知的常规方法,如本文所述。在一个优选的方面,预处理纤维素材料。
预处理的玉米秸秆:术语“PCS”或“预处理的玉米秸秆“在本文中定义为通过用热和稀硫酸处理的源自玉米秸秆的纤维素材料。
分离的多肽:术语“分离的多肽”用于本文中指从来源分离的多肽。在一个优选的方面,多肽如通过SDS-PAGE测定的,为至少1%纯,优选至少5%纯,更优选至少10%纯,更优选至少20%纯,更优选至少40%纯,更优选至少60%纯,甚至更优选至少80%纯,并且最优选至少90%纯。
基本上纯的多肽:术语“基本上纯的多肽”在本文表示多肽制备物,所述多肽制备物含有按重量计至多10%,优选至多8%,更优选至多6%,更优选至多5%,更优选至多4%,更优选至多3%,甚至更优选至多2%,最优选至多1%,并且甚至最优选至多0.5%的与其天然或重组结合的(associated)的其它多肽材料。因此,优选所述基本上纯的多肽是按存在于制备物中的全部多肽材料的重量计至少92%纯,优选至少94%纯,更优选至少95%纯,更优选至少96%纯,更优选至少97%纯,更优选至少98%纯,甚至更优选至少99%纯,最优选至少99.5%纯,并且甚至最优选100%纯。本发明的多肽优选是基本上纯的形式。具体而言,优选所述多肽是“基本上(essentially)纯的形式”,即,所述多肽制备物基本上(essentially)不含与其天然或重组结合的其它多肽材料。例如,这能够通过以下实现:通过公知的重组方法或由经典纯化方法制备多肽。
成熟多肽:术语“成熟多肽”在本文中定义为以其在翻译和任何翻译后修饰之后的最终形式存在的多肽,所述修饰例如N-末端加工、C-末端截短、糖基化、磷酸化等。在一个方面,根据预测SEQ ID NO:2的氨基酸1至19是信号肽的SignalP软件(Nielsen等,1997,Protein Engineering 10:1-6),成熟多肽是SEQ ID NO:2的氨基酸20至856。
成熟多肽编码序列:术语“成熟多肽编码序列”在本文中定义为编码具有β-葡糖苷酶活性的成熟多肽的核苷酸序列。在一个方面,根据预测SEQ ID NO:1的核苷酸1至57编码信号肽的SignalP软件(Nielsen等,1997,见上),成熟多肽编码序列是SEQ ID NO:1的核苷酸58至2568。
序列同一性:参数“序列同一性”描述两个氨基酸序列之间或两个核苷酸序列之间的相关性。
就本发明而言,两个氨基酸序列之间的序列同一性程度使用如EMBOSS软件包(EMBOSS:The European Molecular Biology Open Software Suite,Rice等,2000,Trends in Genetics 16:276-277),优选3.0.0版或更高版本的Needle程序中所执行的Needleman-Wunsch算法(Needleman和Wunsch,1970,J.Mol.Biol.48:443-453)来测定。使用的可选参数为缺口罚分(gap penalty)10,缺口延伸罚分(gap extension penalty)0.5和EBLOSUM62(BLOSUM62的EMBOSS版)取代矩阵。使用Needle标记为“最高同一性(longest identity)”的输出结果(使用-nobrief选项获得)作为同一性百分比,并计算如下:
(同样的残基×100)/(比对长度-比对中缺口的总数)
就本发明而言,两个核苷酸序列之间的序列同一性程度使用如EMBOSS软件包(EMBOS S:The European Molecular Biology Open Software Suite,Rice等,2000,见上文),优选3.0.0版或更高版本的Needle程序中所执行的Needleman-Wunsch算法(Needleman和Wunsch,1970,见上文)来测定。使用的可选参数为缺口罚分10,缺口延伸罚分0.5和EDNAFULL (NCBI NUC4.4的EMBOSS版)取代矩阵。使用Needle标记为“最高同一性”的输出结果(使用-nobrief选项获得)作为同一性百分比,并计算如下:
(同样的脱氧核糖核苷酸×100)/(比对长度-比对中缺口的总数)
同源序列:术语“同源序列”在本文中定义为在用SEQ ID NO:2的Trichophaea saccata β-葡糖苷酶进行的tfasty搜索(Pearson,W.R.,1999,于Bioinformatics Methodsand Protocols中,S.Misener和S.A.Krawetz编,pp.185-219)中E值(或期望值)小于0.001的预测蛋白质。
多肽片段:术语“多肽片段”在本文中定义为从SEQ ID NO:2的成熟多肽或其同源序列的氨基和/或羧基末端缺失一个或多个(几个)氨基酸的多肽;其中所述片段具有β-葡糖苷酶活性。在一个方面,所述片段含有SEQ ID NO:2,SEQ IDNO:2的成熟多肽或其同源序列的至少740个氨基酸残基,更优选至少770个氨基酸残基,并且最优选至少800个氨基酸残基。
亚序列:术语“亚序列(subsequence)”在本文中定义为从SEQ ID NO:1或其同源序列的5’和/或3’端缺失一个或多个(几个)核苷酸的核苷酸序列;其中所述亚序列编码具有β-葡糖苷酶活性的多肽片段。在一个方面,所述亚序列含有SEQ ID NO:1的成熟多肽编码序列或其同源序列的至少2220个核苷酸,更优选至少2310个核苷酸,并且最优选至少2400个核苷酸。
等位变体(allelic variant):术语“等位变体”在本文中表示占据相同染色体基因座的基因的任何两种或更多种(几种)可选形式。等位变异通过突变天然地发生,并且可导致种群内的多态性。基因突变可以是沉默的(在编码的多肽中无变化)或可以编码具有改变的氨基酸序列的多肽。多肽的等位变体是由基因的等位变体编码的多肽。
分离的多核苷酸:术语“分离的多核苷酸”用于本文中指从来源分离的多核苷酸。在一个优选的方面,多核苷酸如通过琼脂糖电泳测定的,为优选至少1%纯,更优选至少5%纯,更优选至少10%纯,更优选至少20%纯,更优选至少40%纯,更优选至少60%纯,甚至更优选至少80%纯,并且最优选至少90%纯。
基本上纯的多核苷酸:术语“基本上纯的多核苷酸”用于本文指不含其它外来的或不期望的核苷酸的多核苷酸制备物,并且所述多核苷酸制备物处于适合于在遗传工程的蛋白质生产体系中使用的形式。因此,基本上纯的多核苷酸含有按重量计优选至多10%,更优选至多8%,更优选至多6%,更优选至多5%,更优选至多4%,更优选至多3%,甚至更优选至多2%,最优选至多1%,并且甚至最优选至多0.5%的与其天然或重组结合的其它多核苷酸材料。然而,基本上纯的多核苷酸可以包括天然存在的5’和3’非翻译区,例如启动子和终止子。优选基本上纯的多核苷酸是按重量计优选至少90%纯,更优选至少92%纯,更优选至少94%纯,更优选至少95%纯,更优选至少96%纯,更优选至少97%纯,甚至更优选至少98%纯,最优选至少99%,并且甚至最优选至少99.5%纯的。本发明所述多核苷酸优选为基本上纯的形式。具体而言,优选在本文公开的多核苷酸是“基本上(essentially)纯的形式”,即,所述多核苷酸制备物基本上不含与其天然或重组结合的其它多核苷酸材料。所述多核苷酸可以是基因组、cDNA、RNA、半合成、合成来源的,或它们的任何组合。
编码序列:当用于本文时术语“编码序列”的意思是直接指定其蛋白产物的氨基酸序列的核苷酸序列。编码序列的边界通常由开读框决定,所述开读框通常以ATG起始密码子或可供选择的起始密码子例如GTG和TTG开始,并且以终止密码子例如TAA、TAG和TGA结束。编码序列可以是DNA、cDNA、合成的或重组的核苷酸序列。
cDNA:术语“cDNA”在本文中定义为能够通过反转录从得自真核细胞的成熟的、已剪接的mRNA分子制备的DNA分子。cDNA缺少通常存在于相应基因组DNA中的内含子序列。起始的(initial)、初级的RNA转录物是mRNA的前体,其通过一系列的步骤加工然后作为成熟的已剪接的mRNA出现。这些步骤包括通过称为剪接的过程去除内含子序列。因而源自mRNA的cDNA没有任何内含子序列。
核酸构建体:术语“核酸构建体”用于本文指分离自天然存在的基因的单链或双链的核酸分子,所述核酸分子受修饰以本来不存在于(not otherwise exist)自然界中的方式含有核酸的区段。所述核酸构建体亦可为“合成的”。当所述核酸构建体含有表达本发明的编码序列所需的调控序列时,术语核酸构建体与术语“表达盒”同义。
调控序列(control sequence):术语“调控序列”在本文定义为包括对编码本发明多肽的多核苷酸表达是必需的或有利的所有成分。各个调控序列对于编码所述多肽的核苷酸序列可以是天然的或外源的,或各个调控序列对于彼此可以是天然的或外源的。这些调控序列包括但不限于前导序列、聚腺苷酸化序列、前肽序列、启动子、信号肽序列和转录终止子。最少的情况,调控序列包括启动子和转录和翻译的终止信号。调控序列可以和用于引入特异性限制位点的接头一起提供,所述特异性限制位点促进调控序列与编码多肽的核苷酸序列编码区的连接。
可操作地连接:术语“可操作地连接”在本文表示这样的构型,其中将调控序列置于相对于多核苷酸序列的编码序列的适当位置,使得调控序列指导多肽编码序列的表达。
表达:术语“表达”包括涉及多肽产生的任何步骤,其包括但不限于转录、转录后修饰、翻译、翻译后修饰和分泌。
表达载体:术语“表达载体”在本文定义为线性的或环状的DNA分子,其包含编码本发明多肽的多核苷酸,并且所述多核苷酸与提供用于其表达的额外核苷酸可操作地连接。
宿主细胞:如本文中所使用的术语“宿主细胞”包括任何细胞类型,所述细胞类型对于使用包含本发明多核苷酸的核酸构建体或表达载体的转化、转染、转导等是易感的(susceptible)。
修饰:术语“修饰”在本文的意思是,对包含SEQ ID NO:2的成熟多肽的多肽,或由SEQ ID NO:2的成熟多肽组成的多肽或其同源序列的任何化学修饰,以及对编码所述多肽的DNA的遗传操作。所述修饰可以是一个或多个(几个)氨基酸的取代、缺失和/或插入,以及一个或多个(几个)氨基酸侧链的置换。
人工变体:当用在本文时,术语“人工变体”的意思是具有β-葡糖苷酶活性的多肽,所述多肽由表达SEQ ID NO:1的成熟多肽编码序列或其同源序列的修饰的核苷酸序列的生物体产生。所述修饰的核苷酸序列通过人为干预(human intervention),通过修饰公开于SEQ ID NO:1或它们的同源序列的核苷酸序列来获得。
发明详述
具有β-葡糖苷酶活性的多肽
在第一个方面,本发明涉及包含氨基酸序列的分离的多肽,所述氨基酸序列与SEQ ID NO:2的成熟多肽具有优选至少60%,更优选至少65%,更优选至少70%,更优选至少75%,更优选至少80%,更优选至少85%,甚至更优选至少90%,最优选至少95%,并且甚至最优选至少96%、至少97%、至少98%或至少99%的序列同一性程度,所述多肽具有β-葡糖苷酶活性(下文中的“同源多肽”)。在一个优选的方面,所述同源多肽包含氨基酸序列,其与SEQID NO:2的成熟多肽相差十个氨基酸,优选相差五个氨基酸,更优选相差四个氨基酸,甚至更优选相差三个氨基酸,最优选相差两个氨基酸,并且甚至最优选相差一个氨基酸。
本发明的多肽优选包含SEQ ID NO:2的氨基酸序列或其等位变体;或它们的具有β-葡糖苷酶活性的片段。在一个优选的方面,多肽包含SEQ ID NO:2的氨基酸序列。在另一个优选方面,多肽包含SEQ ID NO:2的成熟多肽。在另一个优选方面,多肽包含SEQ ID NO:2的氨基酸20至856,或其等位变体;或它们的具有β-葡糖苷酶活性的片段。在另一个优选方面,多肽包含SEQID NO:2的氨基酸20至856。在另一个优选方面,多肽由SEQ ID NO:2的氨基酸序列或其等位变体;或它们的具有β-葡糖苷酶活性的片段组成。在另一个优选方面,多肽由SEQ ID NO:2的氨基酸序列组成。在另一个优选方面,多肽由SEQ ID NO:2的成熟多肽组成。在另一个优选方面,多肽由SEQ ID NO:2的氨基酸20至856或其等位变体;或它们的具有β-葡糖苷酶活性的片段组成。在另一个优选方面,多肽由SEQ ID NO:2的氨基酸20至856组成。
在第二个方面,本发明涉及具有β-葡糖苷酶活性的分离的多肽,所述分离的多肽由多核苷酸编码,所述多核苷酸在优选非常低严格条件下,优选低严格条件下,更优选中严格条件下,更优选中-高严格条件下,甚至更优选高严格条件下,并且最优选非常高严格条件下,与以下杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链(J.Sambrook,E.F.Fritsch,和T.Maniatis,1989,Molecular Cloning,A Laboratory Manual,第2版,Cold Spring Harbor,New York)。
SEQ ID NO:1的核苷酸序列或其亚序列,以及SEQ ID NO:2的氨基酸序列或其片段,可用于设计核酸探针,以根据本领域内公知的方法从不同属和种的菌株鉴定和克隆编码具有β-葡糖苷酶活性的多肽的DNA。具体而言,根据标准的Southern印迹方法,可将这些探针用于与感兴趣的属或种的基因组或cDNA杂交,以鉴定和从其中分离相应的基因。这些探针可明显短于完整序列,但长度上应为至少14,优选至少25,更优选至少35,并且最优选至少70个核苷酸。然而,优选所述核酸探针是至少100个核苷酸长度。例如,所述核酸探针的长度可为至少200个核苷酸,优选至少300个核苷酸,更优选至少400个核苷酸,或最优选至少500个核苷酸。可使用甚至更长的探针,例如长度为优选至少600个核苷酸,更优选至少700个核苷酸,甚至更优选至少800个核苷酸,或最优选至少900个核苷酸的核酸探针。DNA和RNA探针二者均可使用。通常将探针标记以探测相应的基因(例如,用32P、3H、35S、生物素或抗生物素蛋白(avidin)标记)。这些探针涵盖于本发明中。
因而,可从由这些其它生物体制备的基因组DNA或cDNA文库中筛选DNA,所述DNA与上述探针杂交并且编码具有β-葡糖苷酶活性的多肽。可以通过琼脂糖或聚丙烯酰胺凝胶电泳,或通过其它分离技术分离来自这些其它生物体的基因组或其它DNA。可以将来自文库的DNA或分离的DNA转移至硝化纤维素(nitrocellulose)或其它合适的载体材料并且固定于其上。为了鉴定与SEQ ID NO:1或其亚序列同源的克隆或DNA,将所述载体材料优选用在Sounthern印迹中。
就本发明而言,杂交表示核苷酸序列在非常低至非常高的严格条件下与标记的核酸探针杂交,所述核酸探针对应于SEQ ID NO:1的成熟多肽编码序列;包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列;其全长互补链;或它们的亚序列。可使用例如X射线片(X-ray film)检测在这些条件下与核酸探针杂交的分子。
在一个优选的方面,核酸探针是SEQ ID NO:1的成熟多肽编码序列。在另一个优选方面,核酸探针是SEQ ID NO:1的核苷酸58至2568。在另一个优选方面,核酸探针是编码SEQ ID NO:2的多肽的多核苷酸序列,或其亚序列。在另一个优选方面,核酸探针是SEQ ID NO:1。在另一个优选方面,核酸探针是包含在大肠杆菌(E.coli)NRRL B-50214中的质粒pAHYG-33中含有的多核苷酸序列,其中所述其多核苷酸序列编码具有β-葡糖苷酶活性的多肽。在另一个优选方面,核酸探针是包含在大肠杆菌NRRL B-50214中的质粒pAHYG-33中含有的成熟多肽编码区。
对于长度至少100个核苷酸的长探针,将非常低至非常高的严格条件定义为在42℃,在5X SSPE、0.3%SDS、200μg/ml已剪切并且变性的鲑精DNA中,并且对于非常低和低严格性为25%的甲酰胺、对于中和中-高严格性为35%的甲酰胺、或对于高和非常高严格性为50%的甲酰胺,根据标准的Southern印迹法进行预杂交和杂交最佳12至24小时。
对于长度为至少100个核苷酸的长探针,使用2X SSC、0.2%SDS优选至少在45℃(非常低严格性),更优选至少在50℃(低严格性),更优选至少在55℃(中严格性),更优选至少在60℃(中-高严格性),甚至更优选至少在65℃(高严格性),并且最优选至少在70℃(非常高严格性)将载体材料最终洗涤三次,每次15分钟。
对于长度大约15个核苷酸至大约70个核苷酸的短探针,将严格条件定义为在比根据Bolton和McCarthy计算法(1962,Proceedings of the NationalAcademy of Sciences USA 48:1390)得出的Tm低大约5℃至大约10℃,在0.9MNaCl,0.09M Tris-HCl pH 7.6,6mM EDTA,0.5%NP-40,1×Denhardt溶液,1mM焦磷酸钠(sodium pyrophosphate),1mM磷酸二氢钠(sodium monobasicphosphate),0.1mMATP和0.2mg每ml的酵母RNA中,根据标准的Southern印迹步骤进行预杂交、杂交和杂交后洗涤最佳12至24小时。
对于长度大约15个核苷酸至大约70个核苷酸的短探针,将所述载体材料在6×SSC加0.1%SDS中洗涤一次15分钟,并用6×SSC在比计算的Tm低5℃至10℃的温度洗涤两次,每次15分钟。
在第三个方面,本发明涉及由包含核苷酸序列或由核苷酸序列组成的多核苷酸编码的具有β-葡糖苷酶活性的分离的多肽,所述核苷酸序列与SEQ IDNO:1的成熟多肽编码序列具有优选至少60%,更优选至少65%,更优选至少70%,更优选至少75%,更优选至少80%,更优选至少85%,甚至更优选至少90%,最优选至少95%,并且甚至最优选至少96%,至少97%,至少98%,或者至少99%的序列同一性程度,其编码具有β-葡糖苷酶活性的多肽。参见本文的多核苷酸部分。
在第四个方面,本发明涉及SEQ ID NO:2的成熟多肽或其同源序列的包含取代、缺失和/或插入一个或多个(几个)氨基酸的人工变体。优选地,氨基酸改变对性质是较不重要的(of a minor nature),即保守的氨基酸取代或插入,其不显著影响蛋白质的折叠和/或活性;通常为1至大约30个氨基酸的小缺失;小的氨基或羧基末端延伸,如氨基末端甲硫氨酸残基;多至大约20-25个残基的小接头肽;或通过改变净电荷或其它功能来促进纯化的小延伸,如多组氨酸序列(poly histidine tract)、抗原表位(antigenic epitope)或结合域(binding domain)。
保守取代的实例是在以下组之内:碱性氨基酸组(精氨酸、赖氨酸和组氨酸)、酸性氨基酸组(谷氨酸和天冬氨酸)、极性氨基酸组(谷氨酰胺和天冬酰胺)、疏水性氨基酸组(亮氨酸、异亮氨酸和缬氨酸)、芳族氨基酸组(苯丙氨酸、色氨酸和酪氨酸)和小氨基酸组(甘氨酸、丙氨酸、丝氨酸、苏氨酸和甲硫氨酸)。通常不改变比活性(specific activity)的氨基酸取代是本领域已知的,并且由例如H.Neurath和R.L.Hill,1979,于The Proteins,Academic Press,New York中描述。最普遍发生的交换是Ala/Ser、Val/Ile、Asp/Glu、Thr/Ser、Ala/Gly、Ala/Thr、Ser/Asn、Ala/Val、Ser/Gly、Tyr/Phe、Ala/Pro、Lys/Arg、Asp/Asn、Leu/Ile、Leu/Val、Ala/Glu和Asp/Gly。
除了20个基本氨基酸,非基本氨基酸(例如4-羟脯氨酸、6-N-甲基赖氨酸、2-氨基异丁酸、异缬氨酸和α-甲基丝氨酸)可以取代野生型多肽的氨基酸残基。有限数量的非保守氨基酸、不由遗传密码编码的氨基酸和非天然氨基酸可以取代氨基酸残基。“非天然氨基酸”在蛋白质合成后已经过修饰,和/或在它们的侧链具有不同于基本氨基酸的化学结构。非天然氨基酸能够以化学方法合成,并且优选是商业上能够获得的,包括六氢吡啶羧酸(pipecolic acid)、噻唑烷羧酸(thiazolidinecarboxylic acid)、脱氢脯氨酸、3-和4-甲基脯氨酸,和3,3-二甲基脯氨酸。
可供选择的是,氨基酸改变具有这样的性质以使多肽的物理化学性质改变。例如,氨基酸改变可改进多肽的热稳定性,改变底物特异性,改变最适pH等。
能够根据本领域已知的方法,例如定位诱变或丙氨酸分区诱变法(Cunningham和Wells,1989,Science 244:1081-1085)来鉴定亲本多肽中的必需氨基酸。在后一技术中,将单一丙氨酸突变引入到分子中的每个残基,并且测试所得突变分子的生物活性(即,β-葡糖苷酶活性)以鉴定对于所述分子的活性关键的氨基酸残基。同样参见Hilton等,1996,J.Biol.Chem.271:4699-4708。酶的活性部位或其它的生物相互作用也能够通过结构的物理分析而测定,如通过以下这些技术:如核磁共振、晶体学、电子衍射或光亲和标记,连同推定的接触位点氨基酸的突变来测定。参见例如de Vos等,1992,Science 255:306-312;Smith等,1992,J.Mol.Biol.224:899-904;Wlodaver等,1992,FEBSLett.309:59-64。必需氨基酸的身份(identity)也能够从与多肽的同一性分析来推断,所述多肽与根据本发明的多肽相关。
能够使用已知的诱变、重组和/或改组(shuffling)方法,然后是有关的筛选方法,例如那些由Reidhaar-Olson和Sauer,1988,Science 241:53-57;Bowie和Sauer,1989,Proc.Natl.Acad.Sci.USA 86:2152-2156;WO 95/17413;或WO95/22625公开的那些方法来进行并测试单个或多个氨基酸取代。能够使用的其它方法包括易错PCR、噬菌体展示(例如,Lowman等,1991,Biochem.30:10832-10837;美国专利No.5,223,409;WO 92/06204)和区域定向的诱变(Derbyshire等,1986,Gene 46:145;Ner等,1988,DNA 7:127)。
诱变/改组方法能够与高通量、自动化的筛选方法组合以检测由宿主细胞表达的克隆的、诱变的多肽的活性(Ness等,1999,Nature Biotechnology 17:893-896)。能够从宿主细胞回收编码活性多肽的诱变的DNA分子,并且使用本领域内标准方法快速测序。这些方法允许快速测定感兴趣的多肽中单个氨基酸残基的重要性,并且能够应用于未知结构的多肽。
SEQ ID NO:2的成熟多肽的氨基酸取代、缺失和/或插入的总数是10,优选9,更优选8,更优选7,更优选6,更优选5,更优选4,甚至更优选3,最优选2,并且甚至最优选1。
本发明的多肽还包括融合多肽或可切割的融合多肽,其中将另外的多肽融合到所述多肽或其片段的N末端或C末端。通过将编码另一个多肽的核苷酸序列(或其部分)融合于本发明的核苷酸序列(或其部分)来产生融合的多肽。产生融合多肽的技术是本领域已知的,并包括连接编码多肽的编码序列以使它们在阅读框中,并且使融合多肽的表达在相同启动子和终止子的控制下。
融合多肽还可以包括切割位点。一旦分泌了融合多肽,就切割所述位点,从融合蛋白质释放具有β-葡糖苷酶活性的多肽。切割位点的实例包括,但不限于,编码二肽Lys-Arg的Kex2位点(Martin等,2003,J.Ind.Microbiol.Biotechnol.3:568-76;Svetina等,2000,J.Biotechnol.76:245-251;Rasmussen-Wilson等,1997,Appl.Environ.Microbiol.63:3488-3493;Ward等,1995,Biotechnology 13:498-503;和Contreras等,1991,Biotechnology 9:378-381);Ile-(Glu或Asp)-Gly-Arg位点,其在精氨酸残基后通过Factor Xa蛋白酶切割(Eaton等,1986,Biochem.25:505-512);Asp-Asp-Asp-Asp-Lys位点,其在赖氨酸后通过肠激酶切割(Collins-Racie等,1995,Biotechnology 13:982-987);His-Tyr-Glu位点或His-Tyr-Asp位点,其通过Genenase I切割(Carter等,1989,Proteins:Structure,Function,and Genetics 6:240-248);Leu-Val-Pro-Arg-Gly-Ser位点,其在Arg后通过凝血酶切割(Stevens,2003,Drug Discovery World 4:35-48);Glu-Asn-Leu-Tyr-Phe-Gln-Gly位点,其在Gln后通过TEV 蛋白酶切割(Stevens,2003,见上文);和Leu-Glu-Val-Leu-Phe-Gln-Gly-Pro位点,其在Gln后通过基因工程形式的人鼻病毒3C蛋白酶切割(Stevens,2003,见上文)。
具有β-葡糖苷酶活性的多肽的来源
本发明的具有β-葡糖苷酶活性的多肽可以获得自任何属的微生物。就本发明而言,用于本文与给定的来源有关的术语“获得自”,意思是核苷酸序列编码的多肽由所述来源产生,或由其中插入了来自所述来源的核苷酸序列的菌株产生。在一个优选的方面,获得自给定来源的多肽是胞外分泌的。
本发明具有β-葡糖苷酶活性的多肽可以是细菌多肽。例如,所述多肽可以是具有β-葡糖苷酶活性的革兰氏阳性细菌多肽例如芽孢杆菌属(Bacillus)、链球菌属(Streptococcus)、链霉菌属(Streptomyces)、葡萄球菌属(Staphylococcus)、肠球菌属(Enterococcus)、乳杆菌属(Lactobacillus)、乳球菌属(Lactococcus)、梭菌属(Clostridium)、地芽孢杆菌属(Geobacillus)或海洋芽孢杆菌属(Oceanobacillus)多肽;或具有β-葡糖苷酶活性的革兰氏阴性细菌多肽,如大肠杆菌(E.coli)、假单胞菌属(Pseudomonas)、沙门氏菌属(Salmonella)、弯曲杆菌属(Campylobacter)、螺杆菌属(Helicobacter)、黄杆菌属(Flavobacterium)、梭杆菌属(Fusobacterium)、泥杆菌属(Ilyobacter)、奈瑟氏菌属(Neisseria)或脲原体属(Ureaplasma)属多肽。
在一个优选方面,所述多肽是具有β-葡糖苷酶活性的嗜碱芽孢杆菌(Bacillus alkaloph ilus)、解淀粉芽孢杆菌(Bacillus amyloliquefaciens)、短芽孢杆菌(Bacillus brevis)、环状芽孢杆菌(Bacillus circulans)、克劳氏芽孢杆菌(Bacillusclausii)、凝结芽孢杆菌(Bacillus coagulans)、坚强芽孢杆菌(Bacillus firmus)、灿烂芽孢杆菌(Bacillus lautus)、迟缓芽孢杆菌(Bacillus lentus)、地衣芽孢杆菌(Bacillus licheniformis)、巨大芽孢杆菌(Bacillus megaterium)、短小芽孢杆菌(Bacillus pumilus)、嗜热脂肪芽孢杆菌(Bacillus stearothermophilus)、枯草芽孢杆菌(Bacillus subtilis)或苏云金芽孢杆菌(Bacillus thuringiensis)多肽。
在另一个优选的方面,所述多肽是具有β-葡糖苷酶活性的似马链球菌(Streptococcus equisimilis)、酿脓链球菌(Streptococcus pyogenes)、乳房链球菌(Streptococcus uberis)或马链球菌兽瘟亚种(Streptococcus equi subsp.Zooepidemicus)多肽。
在另一个优选的方面,所述多肽是具有β-葡糖苷酶活性的不产色链霉菌(Streptomyces achromogenes)、除虫链霉菌(Streptomyces avermitilis)、天蓝链霉菌(Streptomyces coelicolor)、灰色链霉菌(Streptomyces griseus)或浅青紫链霉菌(Streptomyces lividans)多肽。
本发明具有纤维素分解增强活性的多肽也可以是真菌多肽,并且更优选具有β-葡糖苷酶活性的酵母多肽如假丝酵母属(Candida)、克鲁维酵母属(Kluyveromyces)、毕赤酵母属(Picha)、酵母属(Saccharomyces)、裂殖酵母属(Schizosaccharomyces)或西洋蓍霉属(Yarrowia)多肽;或更优选具有β-葡糖苷酶活性的丝状真菌多肽如枝顶孢霉属(Acremonium)、伞菌属(Agaricus)、链格孢属(Alternaria)、曲霉属(Aspergillus)、短梗霉属(Aureobasidium)、Botryospaeria、拟蜡菌属(Ceriporiopsis)、毛喙壳属(Chaetomidium)、金孢子菌属(Chrysosporium)、Claviceps、Cochliobolus、鬼伞属(Coprinopsis)、Coptotermes、棒囊壳属(Corynascus)、隐丛赤壳菌属(Cryphonectria)、隐球菌属(Cryptococcus)、色二孢属(Diplodia)、黑耳属(Exidia)、Filibasidium、镰孢属(Fusarium)、赤霉属(Gibberella)、全鞭毛虫属(Holomastigotoides)、腐质霉属(Humicola)、耙齿菌属(Irpex)、蘑菇属(Lentinula)、Leptospaeria、梨孢菌属(Magnaporthe)、Melanocarpus、多孔菌属(Meripilus)、毛霉属(Mucor)、毁丝霉属(Myceliophthora)、新考玛脂霉属(Neocallimastix)、脉孢菌属(Neurospora)、拟青霉属(Paecilomyces)、青霉属(Penicillium)、平革菌属(Phanerochaete)、瘤胃壶菌属(Piromyces)、Poitrasia、假黑盘菌属(Pseudoplectania)、Pseudotrichonympha、根毛霉属(Rhizomucor)、裂褶菌属(Schizophyllum)、柱顶孢属(Scytalidium)、踝节菌属(Talaromyces)、嗜热子囊菌属(Thermoascus)、梭孢壳属(Thielavia)、弯颈霉属(Tolypocladium)、木霉属(Trichoderma)、长毛盘菌属(Trichophaea)、轮枝孢属(Verticillium)、包脚菇属(Volvariella)或炭角菌属(Xylaria)多肽。
在一个优选的方面,所述多肽是具有β-葡糖苷酶活性的卡尔酵母(Saccharomyces carlsbergensis)、酿酒酵母(Saccharomyces cerevisiae)、糖化酵母(Saccharomyces diastaticus)、道格拉氏酵母(Saccharomyces douglasii)、克鲁弗酵母(Saccharomyces kluyveri)、诺地酵母(Saccharomyces norbensis)或卵形酵母(Saccharomyces oviformis)多肽。
在另一个优选的方面,所述多肽是具有β-葡糖苷酶活性的解纤维枝顶孢霉(Acremonium cellulolyticus)、棘孢曲霉(Aspergillus aculeatus)、泡盛曲霉(Aspergillus awamori)、烟曲霉(Aspergillus fumigatus)、臭曲霉(Aspergillusfoetidus)、日本曲霉(Aspergillus japonicus)、构巢曲霉(Aspergillus nidulans)、黑曲霉(Aspergillus niger)、米曲霉(Aspergillus oryzae)、嗜角质金孢子菌(Chrysosporiumkeratinophilum)、Chrysosporium lucknowense、热带金孢子菌(Chrysosporiumtropicum)、Chrysosporium merdarium、Chrysosporium inops、毡金孢子菌(Chrysosporium pannicola)、Chrysosporium queenslandicum、Chrysosporiumzonatum、杆孢状镰孢(Fusarium bactridioides)、禾谷镰孢(Fusarium cerealis)、库威镰孢(Fusarium crookwellense)、大刀镰孢(Fusarium culmorum)、禾本科镰孢(Fusarium graminearum)、禾赤镰孢(Fusarium graminum)、异孢镰孢(Fusariumheterosporum)、合欢木镰孢(Fusarium negundi)、尖镰孢(Fusarium oxysporum)、多枝镰孢(Fusarium reticulatum)、粉红镰孢(Fusarium roseum)、接骨木镰孢(Fusarium sambucinum)、肤色镰孢(Fusarium sarcochroum)、拟分枝孢镰孢(Fusarium sporotrichioides)、硫色镰孢(Fusarium sulphureum)、圆镰孢(Fusariumtorulosum)、拟丝孢镰孢(Fusarium trichothecioides)、镶片镰孢(Fusariumvenenatum)、灰腐质霉(Humicola grisea)、特异腐质霉(Humicola insolens)、疏棉状腐质霉(Humicola lanuginosa)、白耙齿菌(Irpex lacteus)、米黑毛霉(Mucormiehei)、嗜热毁丝霉(Myceliophthora thermophila)、粗糙脉孢菌(Neurosporacrassa)、绳状青霉(Penicillium funiculosum)、产紫青霉(Penicillium purpurogenum)、黄孢平革菌(Phanerochaete chrysosporium)、无色梭孢壳(Thielavia achromatica)、Thielavia albomyces、Thielavia albopilosa、澳洲梭孢壳(Thielavia australeinsis)、Thielavia fimeti、小孢梭孢壳(Thielavia microspora)、卵孢梭孢壳(Thielaviaovispora)、Thielavia peruviana、瘤孢梭孢壳(Thielavia spededonium)、毛梭孢壳(Thielavia setosa)、Thielavia subthermophila、土生梭孢霉(Thielavia terrestris)、哈茨木霉(Trichoderma harzianum)、康宁木霉(Trichoderma koningii)、长枝木霉(Trichoderma longibrachiatum)、里氏木霉(Trichoderma reesei)或绿色木霉(Trichoderma viride)多肽。
在另一个优选的方面,所述多肽是具有β-葡糖苷酶活性的Trichophaeaabundans、Trichophaea Trichophaea contradicta、Trichophaea hemisphaerioides、Trichophaea minuta或Trichophaea saccata多肽
在一个更优选的方面,所述多肽是具有β-葡糖苷酶活性的Trichophaeasaccata多肽。在一个最优选的方面,所述多肽是具有β-葡糖苷酶活性的Trichophaea saccata CBS 804.70多肽,例如包含SEQ ID NO:2的氨基酸序列的多肽。
可理解的是对于前述的种,本发明包含完全和不完全阶段(perfect andimperfect states),和其它分类学的等同物(equivalent),例如无性型(anamorph),而无论它们已知的种名。本领域熟练技术人员将容易地识别适合的等同物的同一性。
这些种的菌株在许多培养物保藏中心对于公众能够容易地取得,所述保藏中心诸如美国典型培养物保藏中心(the American Type Culture Collection)(ATCC)、德意志微生物和细胞培养物保藏中心(Deutsche Sammlung vonMikroorganismen und Zellkulturen GmbH)(DSM)、真菌菌种保藏中心(Centraalbureau Voor Schimmelcultures)(CBS)和农业研究机构专利培养物保藏中心北区研究中心(Agricultural Research Service Patent Culture Collection,Northern Regional Research Center)(NRRL)。
此外,可以使用上述的探针从其它来源,包括从自然界(例如,土壤、堆肥、水等)分离的微生物鉴定和获得这些多肽。用于从天然生境(habitat)分离微生物的技术是本领域内公知的。随后可通过相似地筛选这种微生物的基因组或cDNA文库来获得所述多核苷酸。一旦用所述探针检测到编码多肽的多核苷酸,就能够使用本领域普通技术人员熟知的技术将所述多核苷酸分离或克隆(参见,例如,Sambrook等,1989,见上文)。
多核苷酸
本发明还涉及分离的多核苷酸,其包含编码本发明具有β-葡糖苷酶活性的多肽的核苷酸序列,或由编码本发明具有β-葡糖苷酶活性的多肽的核苷酸序列组成。
在一个优选的方面,核苷酸序列包含SEQ ID NO:1或由SEQ ID NO:1组成。在另一个更优选的方面,核苷酸序列包含大肠杆菌NRRL B-50214中所含质粒pAHYG-33中所含的序列,或由大肠杆菌NRRL B-50214中所含质粒pAHYG-33中所含的序列组成。在另一个优选方面,核苷酸序列包含SEQ ID NO:1的成熟多肽编码序列或由SEQ ID NO:1的成熟多肽编码序列组成。在另一个优选方面,核苷酸序列包含SEQ ID NO:1的核苷酸58至2568或由SEQ ID NO:1的核苷酸58至2568组成。在另一个更优选的方面,核苷酸序列包含大肠杆菌NRRL B-50214中所含质粒pAHYG-33中含有的成熟多肽编码序列或由大肠杆菌NRRL B-50214中所含质粒pAHYG-33中含有的成熟多肽编码序列组成。本发明还涵盖编码多肽的核苷酸序列,所述多肽包含SEQ ID NO:2的氨基酸序列或其成熟多肽,或由SEQ ID NO:2的氨基酸序列或其成熟多肽组成;由于遗传密码的简并性,所述核苷酸序列分别不同于SEQ ID NO:1或其成熟多肽编码序列。本发明还涉及SEQ ID NO:1的亚序列,所述亚序列编码具有β-葡糖苷酶活性的SEQ ID NO:2的片段。
本发明还涉及突变多核苷酸,所述突变多核苷酸在SEQ ID NO:1的成熟多肽编码序列中包含至少一个突变,或由在SEQ ID NO:1的成熟多肽编码序列中具有至少一个突变的核苷酸组成,其中所述突变核苷酸序列编码SEQ IDNO:2的成熟多肽。
用于分离或克隆编码多肽的多核苷酸的技术是本领域内已知的,包括从基因组DNA分离,从cDNA制备,或其组合。可通过例如使用熟知的聚合酶链式反应(PCR)或表达文库的抗体筛选来检测具有共有结构特性的克隆DNA片段,从而实现从这种基因组DNA克隆本发明的多核苷酸。参见,例如,Innis等,1990,PCR:A Guide to Methods and Application,Academic Press,New York。可以使用其它核酸扩增方法,如连接酶链式反应(LCR)、连接活化转录(ligatedactivated transcription;LAT)和基于核苷酸序列的扩增(NASBA)。可以从长毛盘菌属(Trichophaea)菌株,或其它或相关生物体克隆多核苷酸,并且因此可为例如所述核苷酸序列的多肽编码区的等位基因变体或种变体(species variant)。
本发明还涉及包含核苷酸序列或由核苷酸序列组成的分离的多核苷酸,所述核苷酸序列与SEQ ID NO:1的成熟多肽编码序列具有优选至少60%,更优选至少65%,更优选至少70%,更优选至少75%,更优选至少80%,更优选至少85%,甚至更优选至少90%,最优选至少95%,并且甚至最优选至少96%,至少97%,至少98%,或者至少99%的序列同一性程度,其编码具有β-葡糖苷酶活性的多肽。
修饰编码本发明多肽的核苷酸序列对于合成与所述多肽基本上相似的多肽可为必需的。术语与所述多肽“基本上相似”指多肽的非天然存在的形式。这些多肽可能以一些工程改造的方式而不同于从其天然来源分离的多肽,例如,比活性、热稳定性、最适pH等方面不同的人工变体。可以在作为SEQ ID NO:1的多肽编码序列存在的核苷酸序列,例如其亚序列的基础上,和/或通过引入如下核苷酸取代来构建变体序列:所述取代不产生由核苷酸序列编码的多肽的另外的氨基酸序列,但是符合意欲产生酶的宿主生物体的密码子选择;或者所述取代可产生不同的氨基酸序列。关于核苷酸取代的概述,参见,例如,Ford等,1991,Protein Expression and Purification 2:95-107。
对于本领域技术人员显而易见的是,这些取代能够在对于分子功能重要的区域之外进行,并且仍然产生活性多肽。对于由本发明的分离的多核苷酸编码的多肽活性关键的并且因此优选不进行取代的氨基酸残基,可以根据本领域公知的方法,例如定位诱变或丙氨酸分区诱变法(参见,例如,Cunningham和Wells,1989,见上文)来鉴定。在后一技术中,将突变引入到分子中的每个荷正电的残基处,并且测试所得突变分子的β-葡糖苷酶活性,以鉴定对于所述分子的活性关键的氨基酸残基。底物-酶相互作用的位点也能够通过分析三维结构测定,通过如核磁共振分析、晶体学或光亲和标记这样的技术来测定(参见,例如,de Vos等,1992,见上文;Smith等,1992,见上文;Wlodaver等,1992,见上文)。
本发明还涉及编码本发明多肽的分离的多核苷酸,所述分离的多核苷酸在优选非常低严格条件下,更优选低严格条件,更优选中等严格条件,更优选中-高严格条件,甚至更优选高严格条件,并且最优选非常高的严格条件下,与以下序列杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链;或它们的等位变体和亚序列(Sambrook等,1989,见上文),如本文所定义的。
本发明还涉及分离的多核苷酸,所述分离的多核苷酸通过以下方法获得:(a)在优选非常低严格条件下,更优选低严格条件,更优选中等严格条件,更优选中-高严格条件,甚至更优选高严格条件,并且最优选非常高的严格条件下,将DNA群体与以下杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链;和(b)分离杂交的多核苷酸,其编码具有β-葡糖苷酶活性的多肽。
核酸构建体
本发明还涉及包含本发明的分离的多核苷酸的核酸构建体,所述分离的多核苷酸与一个或多个(几个)调控序列可操作地连接,所述调控序列在合适的宿主细胞中在与该调控序列相容的条件下指导编码序列的表达。
可以用许多方式操作编码本发明多肽的分离的多核苷酸以提供多肽的表达。依赖于表达载体,在将多核苷酸的序列插入载体之前对其进行操作可能是理想的或必需的。使用重组DNA方法修饰多核苷酸序列的技术是本领域熟知的。
调控序列可以是适当的启动子序列,其是由用于表达编码本发明多肽的多核苷酸的宿主细胞识别的核苷酸序列。启动子序列含有介导多肽的表达的转录调控序列。启动子可以是在所选的宿主细胞中显示转录活性的任何核苷酸序列,包括突变的、截短的和杂合的启动子,并且可以从编码与宿主细胞同源或异源的胞外或胞内多肽的基因获得。
用于指导本发明的核酸构建体转录,特别是在细菌宿主细胞中转录的合适启动子的实例是从下述获得的启动子:大肠杆菌lac操纵子、天蓝链霉菌琼脂糖酶基因(dagA)、枯草芽孢杆菌果聚糖蔗糖酶基因(sacB)、地衣芽孢杆菌α-淀粉酶基因(amyL)、嗜热脂肪芽孢杆菌产麦芽淀粉酶基因(amyM)、解淀粉芽孢杆菌α-淀粉酶基因(amyQ)、地衣芽孢杆菌青霉素酶基因(penP)、枯草芽孢杆菌xylA和xylB基因和原核β-内酰胺酶基因(Villa-Kamaroff等,1978,Proceedings ofthe National Academy of Sciences USA 75:3727-3731),以及tac启动子(DeBoer等,1983,Proceedings of the National Academy of Sciences USA 80:21-25)。另外的启动子在″Useful proteins from recombinant bacteria″于Scientific American,1980,242:74-94中;和在Sambrook等,1989,见上文中描述。
用于指导本发明的核酸构建体在丝状真菌宿主细胞中转录的合适启动子的实例是从下列酶的基因获得的启动子:米曲霉TAKA淀粉酶、曼赫根毛霉(Rhizomucor miehei)天冬氨酸蛋白酶、黑曲霉中性α-淀粉酶、黑曲霉酸稳定性α-淀粉酶、黑曲霉或泡盛曲霉葡糖淀粉酶(glaA)、曼赫根毛霉脂肪酶、米曲霉碱性蛋白酶、米曲霉丙糖磷酸异构酶、构巢曲霉乙酰胺酶、镶片镰孢淀粉葡糖苷酶(WO 00/56900)、镶片镰孢Daria(WO 00/56900)、镶片镰孢Quinn(WO 00/56900)、尖镰孢胰蛋白酶样蛋白酶(WO 96/00787)、里氏木霉β-葡糖苷酶、里氏木霉纤维二糖水解酶I、里氏木霉纤维二糖水解酶II、里氏木霉内切葡聚糖酶I、里氏木霉内切葡聚糖酶II、里氏木霉内切葡聚糖酶III、里氏木霉内切葡聚糖酶IV、里氏木霉内切葡聚糖酶V、里氏木霉木聚糖酶I、里氏木霉木聚糖酶II、里氏木霉β-木糖苷酶,以及NA2-tpi启动子(一种修饰的启动子,其包含在曲霉属中编码中性α-淀粉酶的基因,其中未翻译的前导序列由在曲霉属中编码丙糖磷酸异构酶的基因的未翻译的前导序列所替代;非限制性实例包括修饰的启动子,其包含在黑曲霉中编码中性α-淀粉酶的基因,其中未翻译的前导序列由在构巢曲霉和米曲霉中编码丙糖磷酸异构酶的基因的未翻译的前导序列所替代);和它们的突变的、截短的和杂合的启动子。
在酵母宿主中,有用的启动子从如下酶的基因获得:酿酒酵母烯醇化酶(ENO-1)、酿酒酵母半乳糖激酶(GAL1)、酿酒酵母醇脱氢酶/甘油醛-3-磷酸脱氢酶(ADH1,ADH2/GAP)、酿酒酵母丙糖磷酸异构酶(TPI)、酿酒酵母金属硫蛋白(CUP1)和酿酒酵母3-磷酸甘油酸激酶。对于酵母宿主细胞其它有用的启动子由Romanos等,1992,Yeast 8:423-488描述。
调控序列也可以是合适的转录终止子序列,是由宿主细胞识别以终止转录的序列。所述终止子序列与编码所述多肽的核苷酸序列的3’末端可操作地连接。可以将在所选宿主细胞中有功能的任何终止子用在本发明中。
对于丝状真菌宿主细胞优选的终止子从如下酶的基因获得:米曲霉TAKA淀粉酶、黑曲霉葡糖淀粉酶、构巢曲霉邻氨基苯甲酸合酶、黑曲霉α-葡糖苷酶和尖镰孢胰蛋白酶样蛋白酶。
对于酵母宿主细胞优选的终止子从如下酶的基因获得:酿酒酵母烯醇化酶、酿酒酵母细胞色素C(CYC1)和酿酒酵母甘油醛-3-磷酸脱氢酶。对于酵母宿主细胞其它有用的终止子由Romanos等,1992,见上文描述。
调控序列还可以是合适的前导序列,当被转录时其为对于宿主细胞的翻译重要的mRNA非翻译区。前导序列可操作地连接于编码多肽的核苷酸序列的5’-末端。可以将在所选宿主细胞中有功能的任何前导序列用在本发明中。
对于丝状真菌宿主细胞优选的前导序列从如下酶的基因获得:米曲霉TAKA淀粉酶和构巢曲霉丙糖磷酸异构酶。
对于酵母宿主细胞合适的前导序列从如下酶的基因获得:酿酒酵母烯醇化酶(ENO-1)、酿酒酵母3-磷酸甘油酸激酶、酿酒酵母α因子和酿酒酵母醇脱氢酶/甘油醛-3-磷酸脱氢酶(ADH2/GAP)。
调控序列也可以是聚腺苷酸化序列,其是与核苷酸序列的3’末端可操作地连接的序列,并且在转录时,宿主细胞将其识别为将聚腺苷残基添加至转录的mRNA的信号。可以将在所选宿主细胞中有功能的任何聚腺苷酸化序列在本发明中使用。
对于丝状真菌宿主细胞优选的聚腺苷酸化序列从如下酶的基因获得:米曲霉TAKA淀粉酶、黑曲霉葡糖淀粉酶、构巢曲霉邻氨基苯甲酸合酶、尖镰孢胰蛋白酶样蛋白酶和黑曲霉α-葡糖苷酶。
对于酵母宿主细胞有用的聚腺苷酸化序列由Guo和Sherman,1995,Molecular Cellular Biology 15:5983-5990描述。
调控序列还可以是信号肽编码区,其编码与多肽的氨基末端相连的信号肽,并且指导编码的多肽进入细胞分泌途径。核苷酸序列的编码序列5’端可固有地包含信号肽编码区,其与编码分泌多肽的编码区片段一起天然地连接在翻译阅读框中。可供选择的是,编码序列5’端可含有对于所述编码序列异源的信号肽编码区。异源信号肽编码区在编码序列不天然地含有信号肽编码区时可为必需的。或者,外源信号肽编码区可以简单地取代天然信号肽编码区以增强多肽的分泌。然而,指导表达的多肽进入所选宿主细胞的分泌途径(即,分泌至培养基中)的任何信号肽编码区可在本发明中使用。
对于细菌宿主细胞有效的信号肽编码区是从如下酶的基因获得的信号肽编码区:芽孢杆菌属NCIB 11837产麦芽糖淀粉酶、嗜热脂肪芽孢杆菌α-淀粉酶、地衣芽孢杆菌枯草杆菌蛋白酶(subtilisin)、地衣芽孢杆菌β-内酰胺酶、嗜热脂肪芽孢杆菌中性蛋白酶(nprT,nprS,nprM)和枯草芽孢杆菌prsA。另外的信号肽由Simonen和Palva,1993,Microbiological Reviews 57:109-137描述。
对于丝状真菌宿主细胞有效的信号肽编码区是从如下酶的基因获得的信号肽编码区:米曲霉TAKA淀粉酶、黑曲霉中性淀粉酶、黑曲霉葡糖淀粉酶、曼赫根毛霉天冬氨酸蛋白酶、特异腐质霉纤维素酶、特异腐质霉内切葡聚糖酶V和疏棉状腐质霉脂肪酶。
对于酵母宿主细胞有用的信号肽从酿酒酵母α因子和酿酒酵母转化酶的基因获得。其它有用的信号肽编码区由Romanos等,1992,见上文描述。
在一个优选的方面,信号肽包含SEQ ID NO:2的氨基酸1至19,或者由SEQ ID NO:2的氨基酸1至19组成。在另一个优选方面,信号肽编码序列包含SEQ ID NO:1的核苷酸1至57,或者由SEQ ID NO:1的核苷酸1至57组成。
调控序列还可以是前肽编码区,其编码位于多肽氨基末端的前肽。所得多肽称为酶原(proenzyme)或前多肽(propolypeptide)(或在某些情况下称为酶原(zymogen))。前多肽通常是无活性的并且能够通过前肽的催化或自催化切割从前多肽转化为成熟活性多肽。可以从枯草芽孢杆菌碱性蛋白酶(aprE)、枯草芽孢杆菌中性蛋白酶(nprT)、酿酒酵母α因子、曼赫根毛霉天冬氨酸蛋白酶和嗜热毁丝霉漆酶(WO 95/33836)的基因获得前肽编码区。
当信号肽和前肽区二者均出现在多肽的氨基末端时,将前肽区置于紧接着(next to)多肽氨基末端,并且将信号肽区置于紧接着前肽区的氨基末端。
同样理想的是添加调节序列,其允许相对于宿主细胞的生长来调节多肽的表达。调节系统的实例是引起基因表达响应化学或物理刺激物,包括调节化合物的存在而开启或关闭的那些系统。原核系统中的调节系统包括lac、tac和trp操纵基因系统。在酵母中,可以使用ADH2系统或GAL1系统。在丝状真菌中,可以使用TAKA α-淀粉酶启动子、黑曲霉葡糖淀粉酶启动子和米曲霉葡糖淀粉酶启动子作为调节序列。调节序列的其它实例是那些允许基因扩增的序列。在真核系统中,这些序列包括在氨甲蝶呤(methotrexate)存在下扩增的二氢叶酸还原酶基因,和以重金属(with heavy metal)扩增的金属硫蛋白基因。在这些情况下,编码多肽的核苷酸序列将与调节序列可操作地连接。
表达载体
本发明还涉及重组表达载体,所述重组表达载体包含本发明的多核苷酸、启动子和转录和翻译终止信号。本文所述的多种核酸和调控序列可以结合在一起以产生重组表达载体,所述表达载体可以包括一个或多个(几个)方便的限制位点以允许在这些位点插入或取代编码多肽的核苷酸序列。可供选择的是,可以通过在适当的用于表达的载体中插入包含所述序列的核苷酸序列或核酸构建体来表达本发明的核苷酸序列。在制备表达载体的过程中,将编码序列置于载体中,从而将该编码序列与适当的表达调控序列可操作地连接。
重组表达载体可以是任何载体(例如,质粒或病毒),其能够方便地进行重组DNA步骤,并且能够产生核苷酸序列的表达。载体的选择将通常依赖于载体与将引入该载体的宿主细胞的相容性。载体可以是线状或闭合环状质粒。
载体可以是自主复制载体,即,作为染色体外实体(entity)存在的载体,其复制独立于染色体复制,例如,质粒、染色体外元件、微型染色体(minichromosome)或人工染色体。载体可以含有任何用于确保自复制的手段(means)。或者,载体可以是一种当被引入宿主细胞中时,整合到基因组中并且与整合了该载体的染色体一起复制的载体。此外,可以使用单独的载体或质粒或两个或更多个载体或质粒,其共同含有待引入宿主细胞基因组的完整DNA(total DNA),或可以使用转座子(transposon)。
本发明的载体优选地含有一个或多个(几个)选择性标记,其允许简单选择经转化、转染、转导等的细胞。选择性标记是基因,其产物提供杀生物剂或病毒抗性、对重金属的抗性、对营养缺陷型的原养性(prototrophy to auxotrophs)等。
细菌选择性标记的实例是来自枯草芽孢杆菌或地衣芽孢杆菌的dal基因,或赋予抗生素抗性的标记,所述抗生素抗性例如氨苄青霉素、卡那霉素、氯霉素或四环素抗性。对于酵母宿主细胞合适的标记是ADE2、HIS3、LEU2、LYS2、MET3、TRP1和URA3。用于丝状真菌宿主细胞的选择性标记包括但不限于amdS(乙酰胺酶)、argB(鸟氨酸氨甲酰基转移酶)、bar(草铵膦(phosphinothricin)乙酰转移酶)、hph(潮霉素磷酸转移酶)、niaD(硝酸还原酶)(nitrate reductase)、pyrG(乳清酸核苷-5’-磷酸脱羧酶)(orotidine-5’-phosphatedecarboxylase)、sC(硫酸腺苷酰转移酶)和trpC(邻氨基苯甲酸合酶(anthranilatesynthase))以及它们的等同物。优选用在曲霉属细胞中的是构巢曲霉或米曲霉的amdS和pyrG基因和吸水链霉菌(Streptomyces hygroscopicus)的bar基因。
本发明的载体优选含有元件,其允许载体整合入宿主细胞基因组或载体在细胞中独立于基因组的自主复制。
为了整合入宿主细胞基因组,载体可依赖编码多肽的多核苷酸的序列或用于通过同源或非同源重组整合入基因组的任何其它载体元件。或者,载体可以含有额外的核苷酸序列,用于指导通过同源重组整合入宿主细胞基因组染色体中的精确位置。为了增加在精确位置整合的可能性,整合元件应优选含有足够数量的核酸,如100至10,000碱基对,优选400至10,000碱基对,并且最优选800至10,000碱基对,其与相应的目标序列具有高度序列同一性以增强同源重组的概率。整合元件可以是任何序列,其与宿主细胞基因组中的目标序列同源。此外,整合元件可以是非编码或编码的核苷酸序列。另一方面,可以将载体通过非同源重组整合到宿主细胞的基因组中。
为了自主复制,载体可以进一步包含复制起点,其使载体能够在所述的宿主细胞中自主地复制。复制起点可以是介导自主复制的任何质粒复制子(replicator),其在细胞中发挥功能。术语“复制起点”或“质粒复制子”在本文定义为能够使质粒或载体体内复制的核苷酸序列。
细菌复制起点的实例是允许在大肠杆菌中复制的质粒pBR322、pUC19、pACYC177和pACYC184的复制起点,和允许在芽孢杆菌属中复制的质粒pUB110、pE194、pTA1060和pAMβ1的复制起点。
用于酵母宿主细胞中的复制起点的实例是2微米复制起点,ARS1,ARS4,ARS1和CEN3的组合,和ARS4和CEN6的组合。
在丝状真菌细胞中有用的复制起点的实例是AMA1和ANS1(Gems等,1991,Gene 98:61-67;Cullen等,1987,Nucleic Acids Research 15:9163-9175;WO 00/24883)。分离AMA1基因和构建包含该基因的质粒或载体能够根据公开于WO 00/24883中的方法完成。
可以将多于一个拷贝的本发明的多核苷酸插入宿主细胞以增加基因产物的产生。多核苷酸拷贝数的增加可通过如下方法获得:将至少一个额外拷贝的序列整合入宿主细胞基因组,或将可扩增的选择性标记基因包括于多核苷酸,其中可通过在合适的选择剂(selectable agent)存在下培养细胞来选择含有选择性标记基因的扩增拷贝,且由此含有多核苷酸的额外拷贝的细胞。
用于连接上述元件以构建本发明的重组表达载体的方法是本领域技术人员熟知的(参见,例如,Sambrook等,1989,见上文)。
宿主细胞
本发明还涉及重组宿主细胞,其包含本发明的分离的多核苷酸可操作地连接于一个或多个(几个)指导具有β-葡糖苷酶活性的多肽的产生的调控序列。将包含本发明多核苷酸的构建体或载体导入宿主细胞,使构建体或载体如前所述作为染色体整体或者作为自复制的染色体外载体维持。术语“宿主细胞”包括亲本细胞的任何后代,其由于复制过程中发生的突变而不同于亲本细胞。宿主细胞的选择将在很大程度上依赖于编码多肽的基因及其来源。
宿主细胞可以是在本发明的多肽的重组产生中有用的任何细胞,例如,原核或真核细胞。
原核宿主细胞可以是任何革兰氏阳性细菌或革兰氏阴性细菌。革兰氏阳性细菌包括但不限于,芽孢杆菌属、链球菌属、链霉菌属、葡萄球菌属、肠球菌属、乳酸菌属、乳球菌属、梭菌属、地芽孢杆菌属和海洋芽孢杆菌属。革兰氏阴性细菌包括但不限于,大肠杆菌、假单胞菌属、沙门氏菌属、弯曲杆菌属、螺杆菌属、黄杆菌属、梭杆菌属、泥杆菌属、奈瑟氏菌属和脲原体属。
细菌宿主细胞可以是任何芽孢杆菌属细胞。在本发明的实施中有用的芽孢杆菌属细胞包括但不限于,嗜碱芽孢杆菌、解淀粉芽孢杆菌、短芽孢杆菌、环状芽孢杆菌、克劳氏芽孢杆菌、凝结芽孢杆菌、坚强芽孢杆菌、灿烂芽孢杆菌、迟缓芽孢杆菌、地衣芽孢杆菌、巨大芽孢杆菌、短小芽孢杆菌、嗜热脂肪芽孢杆菌、枯草芽孢杆菌和苏云金芽孢杆菌细胞。
在一个优选的方面,细菌宿主细胞是解淀粉芽孢杆菌。在另一个更优选的方面,细菌宿主细胞是克劳氏芽孢杆菌细胞。在另一个更优选的方面,细菌宿主细胞是迟缓芽孢杆菌。在另一个更优选的方面,细菌宿主细胞是地衣芽孢杆菌细胞。在另一个更优选的方面,细菌宿主细胞是嗜热脂肪芽孢杆菌。在另一个更优选的方面,细菌宿主细胞是枯草芽孢杆菌细胞。
细菌宿主细胞还可以是任何链球菌属细胞。在本发明的实施中有用的链球菌属细胞包括但不限于,似马链球菌、酿脓链球菌、乳房链球菌和马链球菌兽瘟亚种细胞。
在一个优选的方面,细菌宿主细胞是似马链球菌细胞。在另一个优选的方面,细菌宿主细胞是酿脓链球菌细胞。在另一个优选的方面,细菌宿主细胞是乳房链球菌细胞。在另一个优选的方面,细菌宿主细胞是马链球菌兽瘟亚种细胞。
细菌宿主细胞还可以是任何链霉菌属细胞。在本发明的实施中有用的链霉菌属细胞包括但不限于,不产色链霉菌、除虫链霉菌、天蓝链霉菌、灰色链霉菌和浅青紫链霉菌细胞。
在一个优选的方面,细菌宿主细胞是不产色链霉菌细胞。在另一个优选的方面,细菌宿主细胞是除虫链霉菌细胞。在另一个优选的方面,细菌宿主细胞是天蓝链霉菌细胞。在另一个优选的方面,细菌宿主细胞是灰色链霉菌细胞。在另一个优选的方面,细菌宿主细胞是浅青紫链霉菌细胞。
可通过如下方法实现将DNA引入到芽孢杆菌属细胞:例如原生质体转化(参见,例如,Chang和Cohen,1979,MolecularGeneral Genetics 168:111-115),使用感受态细胞(参见,例如,Young和Spizizen,1961,Journal of Bacteriology 81:823-829或Dubnau和Davidoff-Abelson,1971,Journal of Molecular Biology 56:209-221),电穿孔(参见,例如,Shigekawa和Dower,1988,Biotechniques 6:742-751)或接合(参见,例如,Koehler和Thorne,1987,Journal of Bacteriology 169:5771-5278)。可通过如下方法实现将DNA引入到大肠杆菌细胞:例如原生质体转化(参见,例如,Hanahan,1983,J.Mol.Biol.166:557-580)或电穿孔(参见,例如,Dower等,1988,Nucleic Acids Res.16:6127-6145)。可通过如下方法实现将DNA引入到链霉菌属细胞:例如原生质体转化和电穿孔(参见,例如,Gong等,2004,Folia Microbiol.(Praha)49:399-405),接合(参见,例如,Mazodier等,1989,J.Bacteriol.171:3583-3585),或转导(参见,例如,Burke等,2001,Proc.Natl.Acad.Sci.USA 98:6289-6294)。可通过如下方法实现将DNA引入到假单胞菌属细胞:例如电穿孔(参见,例如,Choi等,2006,J.Microbiol.Methods 64:391-397)或接合(参见,例如,Pinedo和Smets,2005,Appl.Environ.Microbiol.71:51-57)。可通过如下方法实现将DNA引入到链球菌属细胞:例如天然感受态(naturalcompetence)(参见,例如,Perry和Kuramitsu,1981,Infect.Immun.32:1295-1297),原生质体转化(参见,例如,Catt和Jollick,1991,Microbios.68:189-207),电穿孔(参见,例如,Buckley等,1999,Appl.Environ.Microbiol.65:3800-3804)或接合(参见,例如,Clewell,1981,Microbiol.Rev.45:409-436)。然而,可以使用本领域已知的将DNA引入宿主细胞的任何方法。
宿主细胞还可以是真核生物,如哺乳动物、昆虫、植物或真菌细胞。
在一个优选的方面,宿主细胞是真菌细胞。“真菌”用在本文包括以下门:子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、壶菌门(Chytridiomycota)和接合菌门(Zygomycota)(如由Hawksworth等,于Ainsworth and Bisby’sDictionary of The Fungi,第8版,1995,CAB International,University Press,Cambridge,UK中所定义)以及卵菌门(Oomycota)(如Hawksworth等,1995,见上,171页中所引用),和所有有丝分裂孢子真菌(mitosporic fungi)(Hawksworth等,1995,见上文)。
在一个更优选的方面,真菌宿主细胞是酵母细胞。“酵母”用在本文包括产子囊酵母(ascosporogenous yeast)(内孢霉目(Endomycetales))、产担子酵母(basidiosporogenous yeast)和属于半知菌类(Fungi Imperfecti)(芽孢纲(Blastomycetes))的酵母。由于酵母的分类在未来可能改变,就本发明而言,将酵母定义为如Biology and Activities of Yeast(Skinner,F.A.,Passmore,S.M.,和Davenport,R.R.编,Soc.App.Bacteriol.Symposium Series No.9,1980)中所述。
在一个甚至更加优选的方面,酵母宿主细胞是假丝酵母属、汉逊酵母属(Hansenula)、克鲁维酵母属、毕赤酵母属、酵母属、裂殖酵母属或西洋蓍霉属细胞。
在一个最优选的方面,酵母宿主细胞是卡尔酵母细胞。在另一个最优选的方面,酵母宿主细胞是酿酒酵母细胞。在另一个最优选的方面,酵母宿主细胞是糖化酵母细胞。在另一个最优选的方面,酵母宿主细胞是道格拉氏酵母细胞。在另一个最优选的方面,酵母宿主细胞是克鲁弗酵母细胞。在另一个最优选的方面,酵母宿主细胞是诺地酵母细胞。在另一个最优选的方面,酵母宿主细胞是卵形酵母细胞。在另一个最优选的方面,酵母宿主细胞是乳酸克鲁维酵母(Kluyveromyces lactis)细胞。在另一个最优选的方面,酵母宿主细胞是解脂西洋蓍霉(Yarrowia lipolytica)细胞。
在另一个更优选的方面,真菌宿主细胞是丝状真菌细胞。“丝状真菌”包括真菌门(Eumycota)和卵菌门的亚门(如由Hawksworth等,1995,见上文,所定义)的所有丝状形式。丝状真菌通常的特征在于由壳多糖(chitin)、纤维素、葡聚糖、壳聚糖(chitosan)、甘露聚糖和其它复杂多糖组成的菌丝体壁。通过菌丝延伸进行营养生长,而碳分解代谢是专性需氧的。相反,酵母例如酿酒酵母的营养生长通过单细胞菌体的出芽生殖(budding)进行,而碳分解代谢可以是发酵的。
在甚至更优选的方面,丝状真菌宿主细胞是枝顶孢霉属、曲霉属、短梗霉属、烟管霉属(Bjerkandera)、拟蜡菌属、金孢子菌属、鬼伞属(Coprinus)、革盖菌属(Coriolus)、隐球菌属、Filibasidium、镰孢属、腐质霉属、梨孢菌属、毛霉属、毁丝霉属、新考玛脂霉属、脉孢菌属、拟青霉属、青霉属、平革菌属(Phanerochaete)、射脉菌属(Phlebia)、瘤胃壶菌属、侧耳属(Pleurotus)、裂褶菌属、踝节菌属、嗜热子囊菌属、梭孢壳属、弯颈霉属、栓菌属(Trametes)或木霉属细胞。
在最优选的方面,丝状真菌宿主细胞是泡盛曲霉、烟曲霉、臭曲霉、日本曲霉、构巢曲霉、黑曲霉或米曲霉细胞。在另一个最优选方面,丝状真菌宿主细胞是杆孢状镰孢、禾谷镰孢、库威镰孢、大刀镰孢、禾本科镰孢、禾赤镰孢、异孢镰孢、合欢木镰孢、尖镰孢、多枝镰孢、粉红镰孢、接骨木镰孢、肤色镰孢、拟分枝孢镰孢、硫色镰孢、圆镰孢、拟丝孢镰孢或镶片镰孢细胞。在另一个最优选的方面,丝状真菌宿主细胞是黑刺烟管菌(Bjerkandera adusta)、干拟蜡菌(Ceriporiopsis aneirina)、干拟蜡菌、Ceriporiopsis caregiea、Ceriporiopsisgilvescens、Ceriporiopsis pannocinta、Ceriporiopsis rivulosa、Ceriporiopsis subrufa、虫拟蜡菌(Ceriporiopsis subvermispora)、嗜角质金孢子菌、Chrysosporiumlucknowense、热带金孢子菌、Chrysosporium merdarium、Chrysosporium inops、毡金孢子菌、Chrysosporium queenslandicum、Chrysosporium zonatum、灰盖鬼伞(Coprmus cinereus)、毛革盖菌(Coriolus hirsutus)、特异腐质霉、疏棉状腐质霉、米黑毛霉、嗜热毁丝霉、粗糙脉孢菌、产紫青霉、黄孢平革菌(Phanerochaetechrysosporium)、辐射射脉菌(Phlebia radiata)、刺芹侧耳(Pleurotus eryngii)、土生梭孢霉、长绒毛栓菌(Trametes villosa)、变色栓菌(Trametes versicolor)、哈茨木霉、康宁木霉、长枝木霉、里氏木霉或绿色木霉细胞。
在另一个优选实施方案中,所述丝状真菌宿主细胞是黑曲霉细胞。在另一个最优选的方面,所述丝状真菌宿主细胞是米曲霉细胞。在另一个最优选的方面,所述丝状真菌宿主细胞是Chrysosporium lucknowense细胞。在另一个最优选的方面,所述丝状真菌宿主细胞是镶片镰孢细胞。在另一个最优选的方面,所述丝状真菌宿主细胞是嗜热毁丝霉细胞。在另一个最优选的方面,所述丝状真菌宿主细胞是里氏木霉细胞。
可以将真菌细胞通过涉及原生质体形成、原生质体转化和细胞壁重建的方法以本身公知的方式转化。用于转化曲霉属和木霉属宿主细胞的合适方法在EP 238 023和Yelton等,1984,Proceedings of the National Academy ofSciences USA 81:1470-1474中描述。用于转化镰孢属菌种的合适方法由Malardier等,1989,Gene 78:147-156和WO 96/00787描述。可以使用由如下文献描述的方法转化酵母:Becker和Guarente,于Abelson,J.N.和Simon,M.I.编,Guide to Yeast Genetics and Molecular Biology,Methods in Enzymology,Volume 194,pp 182-187,Academic Press,Inc.,New York;Ito等,1983,Journalof Bacteriology 153:163;和Hinnen等,1978,Proceedings of the NationalAcademy of Sciences USA 75:1920。
产生方法
本发明还涉及用于产生本发明多肽的方法,其包括:(a)在有助于产生多肽的条件下培养细胞,所述细胞以其野生型形式产生所述多肽;和(b)回收所述多肽。在一个优选的方面,所述细胞是长毛盘菌属的细胞。在更优选的方面,所述细胞是Trichophaea saccata。在最优选的方面,所述细胞是Trichophaea saccataCBS 804.70。
本发明还涉及用于产生本发明的多肽的方法,其包括:(a)如本文所述,在有助于产生多肽的条件下培养重组宿主细胞;和(b)回收所述多肽。
本发明还涉及用于产生本发明的多肽的方法,包括:(a)在有助于产生多肽的条件下培养重组宿主细胞,其中所述宿主细胞包含突变核苷酸序列,其在SEQ ID NO:1的成熟多肽编码序列中具有至少一个突变,其中所述突变核苷酸序列编码多肽,该多肽包含SEQ ID NO:2的成熟多肽或由SEQ ID NO:2的成熟多肽组成,和(b)回收所述多肽。
在本发明的产生方法中,使用本领域熟知的方法在适合于产生所述多肽的营养培养基中培养细胞。例如,可以通过在合适培养基中和允许表达和/或分离所述多肽的条件下进行的摇瓶培养,和实验室或工业发酵罐中的小规模或大规模发酵(包括连续、分批、补料分批或固态发酵)来培养细胞。使用本领域已知的方法在合适的营养培养基中进行培养,所述营养培养基包含碳源和氮源和无机盐。合适的培养基能够从商业供应商获得或可以根据公开的组成制备(例如,在美国典型培养物保藏中心的目录中)。如果多肽分泌到营养培养基中,该多肽能够从所述培养基中直接回收。如果多肽不分泌到培养基中,其能够从细胞裂解物(lysate)回收。
可以使用本领域已知的对于所述多肽是特异性的方法来检测多肽。这些检测方法可包括特异性抗体的使用、酶产物的形成或酶底物的消失。例如,酶试验(enzyme assay)可用于测定如本文所述的多肽的活性。
所得多肽可以使用本领域已知的方法回收。例如,多肽可以通过常规方法从营养培养基中回收,所述常规方法包括但不限于离心、过滤、提取、喷雾干燥、蒸发或沉淀。
本发明的多肽可以通过多种本领域已知的方法纯化以获得基本上纯的多肽,所述方法包括但不限于层析(例如,离子交换、亲和、疏水、层析聚焦和大小排阻)、电泳方法(例如,制备型(preparative)等电聚焦)、差示溶解度(例如,硫酸铵沉淀)、SDS-PAGE或提取(参见,例如,Protein Purification,J.-C.Janson和Lars Ryden,editors,VCH Publishers,New York,1989)。
植物
本发明还涉及植物,例如,转基因植物、植物部分或植物细胞,其包含分离的多核苷酸,所述多核苷酸编码本发明具有β-葡糖苷酶活性的多肽,从而以可回收的量表达和产生所述多肽。多肽可从植物或植物部分回收。或者,同样可以将含有该重组多肽的植物或植物部分用于改进食品或饲料的质量,例如,改进营养价值、适口性(palatability)和流变性质(rheological properties),或用于破坏抗营养因子。
转基因植物可以是双子叶的(双子叶植物)或单子叶的(单子叶植物)。单子叶植物的实例是草(grasses),如草地早熟禾(meadow grass)(蓝草(blue grass),早熟禾属(Poa));饲用牧草(forage grass)如羊茅属(Festuca)、黑麦草属(Lolium);寒地型牧草(temperate grass),如Agrostis(翦股颖属);和谷类,例如,小麦、燕麦、黑麦、大麦、稻(rice)、高粱和玉蜀黍(maize)(玉米)。
双子叶植物的实例是烟草(tobacco),豆类(legumes),如羽扇豆(lupins),马铃薯,糖甜菜(sugar beet),豌豆,豆(bean)和大豆(soybean)和十字花科的(cruciferous)植物(十字花科(family Brassicaceae)),如花椰菜(cauliflower),油菜籽(rape seed)和紧密相关的模型生物体拟南芥(Arabidopsis thaliana)。
植物部分的实例是茎(stem)、愈伤组织(callus)、叶(leaf)、根(root)、果实(fruit)、种子(seed)和块茎(tuber),以及包含这些部分的独立组织,例如,表皮(epidermis)、叶肉(mesophyll)、薄壁组织(parenchyme)、维管组织(vasculartissue)、分生组织(meristem)。具体的植物细胞区室(compartments),如叶绿体(chloroplast)、质外体(apoplast)、线粒体(mitochondria)、液泡(vacuole)、过氧化物酶体(peroxisome)和细胞质(cytoplasm)也被认为是植物部分。此外,任何植物细胞,无论什么组织来源,都被认为是植物部分。同样地,植物部分,如分离以促进本发明的应用的具体组织和细胞也被认为是植物部分,例如胚(embryo)、胚乳(endosperm)、糊粉(aleurone)和种皮(seed coat)。
同样包含于本发明范围内的还有这些植物、植物部分和植物细胞的子代。
表达本发明多肽的转基因植物或植物细胞可以依照本领域已知方法构建。简而言之,通过如下方法构建所述植物或植物细胞:将编码本发明多肽的一个或多个(几个)表达构建体并入植物宿主基因组或叶绿体基因组,并且将所得的修饰植物或植物细胞繁殖为转基因植物或植物细胞。
表达构建体便利地是包含编码本发明多肽的多核苷酸的核酸构建体,所述多核苷酸与在选择的植物或植物部分中表达该多核苷酸序列所需的适当的调节序列可操作地连接。此外,表达构建体可以包含对于鉴定宿主细胞有用的选择性标记,在所述宿主细胞中整合了表达构建体和将该构建体引入到所述植物中所必需的DNA序列(后者依赖于使用的DNA引入方法)。
调节序列的选择,例如启动子和终止子序列和任选地信号或转运序列的选择,举例来说,基于期望何时、何处以及如何表达多肽而确定。例如,编码本发明多肽的基因的表达可以是组成型的或诱导型的,或可以是发育、阶段或组织特异性的,并且基因产物可以靶向特定的组织或植物部分例如种子或叶。调节序列由例如Tague等,1988,Plant Physiology 86:506所述。
对于组成性表达,可以使用35S-CaMV、玉米泛素1和稻肌动蛋白1启动子(Franck等,1980,Cell 21:285-294,Christensen等,1992,Plant Mo.Biol.18:675-689;Zhang等,1991,Plant Cell 3:1155-1165)。器官特异性启动子可以是例如来自贮藏库组织(storage sink tissue)例如种子、马铃薯块茎和果实的启动子(Edwards和Coruzzi,1990,Ann.Rev.Genet.24:275-303),或来自代谢库组织(metabolic sink tissue)例如分生组织的启动子(Ito等,1994,Plant Mol.Biol.24:863-878),种子特异性启动子诸如来自稻的谷蛋白(glutelin)、醇溶蛋白(prolamin)、球蛋白(globulin)或白蛋白(albumin)启动子(Wu等,1998,Plant and CellPhysiology 39:885-889),来自豆球蛋白(legumin)B4和蚕豆(Vicia faba)的未知的种子蛋白基因的蚕豆启动子(Conrad等,1998,Journal of Plant Physiology152:708-711)、来自种子油体蛋白(oil body protein)的启动子(Chen等,1998,Plantand Cell Physiology 39:935-941),来自欧洲油菜(Brassica napus)的贮藏蛋白napA启动子,或本技术领域公知的任何其他种子特异性的启动子,例如,在WO91/14772中所描述的。此外,启动子可为叶特异性的启动子,如来自稻或番茄的rbcs启动子(Kyozuka等,1993,Plant Physiology 102:991-1000),小球藻病毒(chlorella virus)腺嘌呤甲基转移酶(adenine methyltransferase)基因启动子(Mitra和Higgins,1994,Plant Molecular Biology 26:85-93),或来自稻的aldP基因启动子(Kagaya等,1995,Molecular and general genetics 248:668-674),或伤口诱导的启动子,如马铃薯pin2启动子(Xu等,1993,Plant Molecular Biology 22:573-588)。同样地,所述启动子可通过非生物的处理诱导,所述非生物的处理诸如温度、干旱或盐度变化,或通过外源施加的激活所述启动子的物质诱导,例如乙醇、雌激素(oestrogens)、植物激素(plant hormones)如乙烯、脱落酸(abscisic acid)和赤霉酸(gibberellic acid),和重金属。
启动子增强子元件也可以用于实现本发明多肽在植物中的较高表达。例如,启动子增强子元件可以是内含子,其置于启动子和编码本发明多肽的核苷酸序列之间。例如Xu等,1993,见上,公开了使用稻肌动蛋白1基因的第一内含子以增强表达。
选择性标记基因和表达构建体的任何其它部分可以选自本领域内可用的那些。
将核酸构建体根据本领域已知的常规技术并入植物基因组,所述常规技术包括土壤杆菌属(Agrobacterium)介导的转化、病毒介导的转化、显微注射(microinjection)、粒子轰击、生物射弹转化和电穿孔(Gasser等,1990,Science 244:1293;Potrykus,1990,Bio/Technology 8:535;Shimamoto等,1989,Nature 338:274)。
目前,根癌土壤杆菌(Agrobacterium tumefaciens)介导的基因转移(genetransfer),是产生转基因双子叶植物的优选方法(为了参考,见Hooykas和Schilperoort,1992,Plant Molecular Biology 19:15-38),而且它也可以用于转化单子叶植物,虽然对于这些植物其他的转化方法是常用的。目前,产生转基因单子叶植物的优选的方法,是用粒子(用转化DNA涂覆的微观的金或钨粒子)轰击胚愈伤组织(embryonic calli)或发育中的胚(developing embryos)(Christou,1992,Plant Journal 2:275-281;Shimamoto,1994,Current Opinion Biotechnology 5:158-162;Vasil等,1992,Bio/Technology 10:667-674)。转化单子叶植物的可供选择的方法是基于原生质体转化,如由Omirulleh等,1993,Plant Molecular Biology21:415-428所描述的。其他用于依照本公开使用的转化方法包括那些描述于美国专利6,395,966和7,151,204的那些(两者均通过全文提述并入本文)。
转化之后,根据本领域熟知的方法选择具有并入的表达构建体的转化体并且再生成为完整植物。通常设计转化方法用于通过如下方法在再生期间或在后续世代中选择性消除选择基因:例如,使用带有两个独立的T-DNA构建体的共转化或通过特异性重组酶位点特异性地切除选择基因。
本发明还涉及用于产生本发明多肽的方法,其包括:(a)在有助于产生多肽的条件下培养包含多核苷酸的转基因植物或植物细胞,所述多核苷酸编码本发明具有β-葡糖苷酶活性的多肽;和(b)回收所述多肽。
除了直接用根据本发明制备的构建体直接转化具体植物基因型之外,还可通过将具有本发明的构建体的植物与缺乏该构建体的第二植物杂交来制备转基因植物。举例而言,可将编码具有β-葡糖苷酶活性的多肽的构建体或其部分通过杂交而引入特定植物品种,而根本无需直接转化该给定品种的植物。因此,本发明不仅涵盖从依照本发明经转化的细胞直接再生的植物,还包括此类植物的后裔(progeny)。如用于本文,后裔可指依照本发明制备的亲本植物任何世代的后代(offspring)。此种后裔可包含依据本发明制备的DNA构建体,或依据本发明制备的DNA构建体的一部分。杂交导致本发明的转基因通过将起始种系与包含本发明的转基因的供体植物种系交叉授粉而引入植物种系。此类步骤的非限制性实例进一步阐述于美国专利7,151,204号。
涵盖了包含本发明的具有β-葡糖苷酶活性的多肽的植物包括通过回交转化方法生成的植物。举例而言,本发明的植物包括称作回交转化的基因型、种系、近交体(inbred)或杂交体(hybrid)的植物。
可使用遗传标记以协助本发明的一种或多种转基因从一个遗传背景基因渗入(introgression)至另一个。标记协助的选择提供了相对于常规育种的优势,在于其可用于避免由表型变异导致的错误。进一步,遗传标记可在特定杂交的个体后裔中提供有关良种种质相对程度的数据。举例而言,当本不(otherwise)具有非农艺学所需的遗传背景但具有所需性状的植物与良种亲本杂交时,可使用遗传标记来选择不仅具有目标性状,还具有相对较大比例所需种质的后裔。以此方式,使一种或多种性状基因渗入特定遗传背景所需的世代数得到最小化。
去除或减少β-葡糖苷酶活性
本发明还涉及用于产生亲本细胞突变体的方法,其包括破坏或缺失编码本发明的多肽的多核苷酸或其部分,所述方法导致在相同条件下培养时,与亲本细胞相比突变的细胞产生较少的所述多肽。
可以使用本领域熟知的方法(例如,插入、破坏、替代或缺失)通过减少或消除编码本发明多肽的核苷酸序列的表达来构建突变细胞。在一个优选的方面,所述核苷酸序列是失活的。待修饰或失活的核苷酸序列可以是,例如,编码区或其对活性关键的部分,或表达编码区所需的调节元件。这种调节或调控序列的实例可以是启动子序列或其功能部分,即,足以影响核苷酸序列表达的部分。用于可能的修饰的其它调控序列包括但不限于前导序列、聚腺苷酸化序列、前肽序列、信号肽序列、转录终止子和转录激活子。
可以通过向亲本细胞施以诱变,并且选择其中已将所述核苷酸序列的表达减少或消除的突变细胞来进行核苷酸序列的修饰或失活。诱变可能是特异性的或随机的,可以通过例如使用合适的物理或化学诱变剂进行,通过使用合适的寡核苷酸进行,或通过将所述DNA序列进行PCR产生的诱变。此外,可以通过使用这些诱变剂的任何组合来进行诱变。
适合于本发明目的的物理或化学诱变剂的实例包括紫外线(UV)照射、羟胺、N-甲基-N′-硝基-N-亚硝基胍(MNNG)、O-甲基羟胺、亚硝酸、乙基甲烷磺酸酯(ethyl methane sulphonate)(EMS)、亚硫酸氢钠、甲酸和核苷酸类似物。
当使用这些试剂时,通常通过如下方法来进行所述诱变:在合适条件下存在优选的诱变剂时温育待诱变的亲本细胞,并筛选和/或选择显示基因表达减少的或无基因表达的突变体细胞。
通过导入、取代或去除基因中的一个或多个(几个)核苷酸或其转录或翻译所需的调控元件可以实现所述核苷酸序列的修饰或失活。例如,可以插入或去除核苷酸从而导致引入终止密码子,去除起始密码子,或改变开放阅读框。按照本领域已知的方法通过定位诱变或PCR产生的诱变可以实现这种修饰或失活。尽管在理论上所述修饰可以在体内进行,即,直接在表达待修饰核苷酸序列的细胞上进行,但优选如下面所示例的那样在体外进行所述修饰。
消除或减少核苷酸序列通过细胞表达的便利方式的例子是基于基因取代,基因缺失,或基因破坏的技术。例如,在基因破坏方法中,将相应于内源核苷酸序列的核酸序列在体外进行诱变以产生缺陷性的核酸序列,然后将其转化入亲本细胞中以产生缺陷基因。通过同源重组,所述缺陷性核酸序列替代了内源性核苷酸序列。理想的是所述缺陷性核苷酸序列还编码标记,其可用于选择其中核苷酸序列被修饰或破坏的转化子。在特别优选的方面,用可选择的标记(如本文所述的那些)来破坏所述核苷酸序列。
或者,可以使用与所述核苷酸序列互补的序列通过确定的反义或RNAi技术来进行所述核苷酸序列的修饰或失活。更具体地,通过导入与所述基因的核苷酸序列互补的序列可以减少或消除所述核苷酸序列通过细胞的表达,所述序列可以在细胞中转录并且能够与细胞中产生的mRNA杂交。由此在允许所述互补反义核苷酸序列与mRNA杂交的条件下,减少或消除翻译的蛋白质的量。
本发明进一步涉及亲本细胞的突变体细胞,其包含编码多肽的核苷酸序列或其调控序列的破坏或缺失,这导致与亲本细胞相比突变体细胞产生更少的多肽或不产生多肽。
这样生成的多肽缺陷型突变细胞作为表达天然和/或异源多肽的宿主细胞特别有用。所以,本发明进一步涉及生产天然或异源多肽的方法,其包括:(a)在有助于生产多肽的条件下培养突变细胞;和(b)回收所述多肽。术语“异源多肽”在本文中定义为对宿主细胞不是天然的多肽,进行了修饰以改变天然序列的天然蛋白,或作为通过重组DNA技术对宿主细胞操作的结果其表达在量上改变的天然蛋白。
在另一方面,本发明涉及通过发酵可产生本发明的多肽以及目标蛋白产物的细胞来生产基本上无β-葡糖苷酶活性的蛋白质产品的方法:在发酵之前、过程中或发酵完成之后向发酵液(fermentation broth)中添加有效量的能够抑制β-葡糖苷酶活性的试剂,从发酵液中回收目标产物,并且任选地将回收的产物进行进一步纯化。
在另一方面,本发明涉及如下生产基本上无β-葡糖苷酶活性的蛋白质产品的方法:在允许产物表达的条件下培养细胞,将得到的培养液进行组合的pH和温度处理以基本上减少β-葡糖苷酶活性,和从发酵液中回收产物。或者,可以将从培养液回收的酶制备物进行组合的pH和温度处理。所述组合的pH和温度处理可任选地与用β-葡糖苷酶抑制剂处理组合使用。
依照本发明的这个方面,可能去除至少60%,优选至少75%,更优选至少85%,还更优选至少95%,并且最优选至少99%的β-葡糖苷酶活性。可使用此方法获得β-葡糖苷酶活性的完全去除。
组合的pH和温度处理优选在2-4或9-11范围内的pH和至少60-70℃范围内的温度进行一段足够的时间以达到期望的效果,通常30至60分钟是足够的。
用于培养和纯化感兴趣的产物的方法可以通过本领域已知的方法进行。
本发明用于产生基本上无β-葡糖苷酶活性的产物的方法在真核多肽,特别是真菌蛋白质例如酶的产生中是特别令人有兴趣的。所述酶可以选自,例如,淀粉分解酶(amylolytic enzyme)、脂肪分解酶、蛋白水解酶、纤维素分解酶(cellulolytic enzyme)、氧化还原酶或植物细胞壁降解酶。这些酶的实例包括氨肽酶、淀粉酶、淀粉葡糖苷酶、糖酶、羧肽酶、过氧化氢酶、纤维二糖水解酶、纤维素酶、几丁质酶(chitinase)、角质酶(cutinase)、环糊精糖基转移酶、脱氧核糖核酸酶、内切葡聚糖酶、酯酶、半乳糖苷酶、β-半乳糖苷酶、葡糖淀粉酶、葡糖氧化酶、葡糖苷酶、卤素过氧化物酶、半纤维素酶、转化酶、异构酶、漆酶、连接酶、脂肪酶、裂合酶、甘露糖苷酶、氧化酶、果胶分解酶、过氧化物酶、肌醇六磷酸酶、酚氧化酶、多酚氧化酶、蛋白水解酶、核糖核酸酶、转移酶、转谷氨酰胺酶或木聚糖酶。β-葡糖苷酶-缺陷细胞也可以用于表达在制药上感兴趣的异源蛋白质例如激素、生长因子、受体等。
可理解的是术语“真核多肽”不仅包括天然多肽,也包括多肽例如酶,其通过氨基酸取代、缺失或添加或其它这样的修饰而被修饰以增强活性、热稳定性、pH耐受性等。
在另外的方面,本发明涉及基本上无β-葡糖苷酶活性的蛋白质产物,其通过本发明的方法产生。
抑制具有β-葡糖苷酶活性的多肽表达的方法
本发明还涉及抑制具有β-葡糖苷酶活性的多肽在细胞中表达的方法,其包括对细胞施用双链RNA(dsRNA)分子或者在细胞中表达双链RNA(dsRNA)分子,其中dsRNA包含本发明多核苷酸的亚序列。在一个优选的方面,dsRNA长约15、16、17、18、19、20、21、22、23、24、25或更多的双链体核苷酸。
dsRNA优选是小干扰RNA(siRNA)或微RNA(miRNA)。在一个优选的方面,dsRNA是用于抑制转录的小干扰RNA(siRNA)。在另一个优选的方面,dsRNA是用于抑制翻译的小RNA(miRNA)。
本发明还涉及用于抑制细胞中具有β-葡糖苷酶活性的多肽表达的这些双链RNA(dsRNA)分子,其包含SEQ ID NO:1的成熟多肽编码序列的部分。本发明不受限于任何特定作用机制,dsRNA能进入细胞,并引起相似或相同序列的单链RNA(ssRNA)的降解,所述RNA包括内源mRNA。当细胞接触dsRNA时,来自同源基因的mRNA选择性地受到称为RNA干扰(RNAi)的过程的降解。
本发明的dsRNA可以用于基因沉默。在一个方面,本发明提供使用本发明的dsRNAi选择性地降解RNA的方法。所述过程可以在体外、在活体外或离体(ex vivo)或在体内进行。在一个方面,dsRNA分子能够用于产生细胞、器官或动物中的功能缺失突变(loss-of-function mutation)。制备或使用选择性降解RNA的dsRNA分子的方法在本领域中公知,参见,例如,美国专利No.6,506,559;美国专利No.6,511,824;美国专利No.6,515,109;和美国专利No.6,489,127。
组合物
本发明还涉及包含本发明的多肽的组合物。优选地,所述组合物富含这种多肽。术语“富含”表示所述组合物的β-葡糖苷酶活性,例如,以至少1.1的富集因数(enrichment factor)增加。
所述组合物可以包含本发明的多肽作为主要酶成分,例如,单成分组合物。或者,所述组合物可以包含多种酶活性,如氨肽酶、淀粉酶、糖酶、羧肽酶、过氧化氢酶、纤维素酶、几丁质酶、角质酶、环糊精糖基转移酶、脱氧核糖核酸酶、酯酶、α-半乳糖苷酶、β-半乳糖苷酶、葡糖淀粉酶、α-葡糖苷酶、β-葡糖苷酶、卤素过氧化物酶、转化酶、漆酶、脂肪酶、甘露糖苷酶、氧化酶、果胶分解酶、肽谷氨酰胺酶(peptidoglutaminase)、过氧化物酶、肌醇六磷酸酶、多酚氧化酶、蛋白水解酶、核糖核酸酶、转谷氨酰胺酶或木聚糖酶。额外的酶可以通过例如属于以下属或种的微生物产生:曲霉属,优选棘孢曲霉、泡盛曲霉、烟曲霉、臭曲霉、日本曲霉、构巢曲霉、黑曲霉或米曲霉;镰孢属,优选杆孢状镰孢、禾谷镰孢、库威镰孢、大刀镰孢、禾本科镰孢、禾赤镰孢、异孢镰孢、合欢木镰孢、尖镰孢、多枝镰孢、粉红镰孢、接骨木镰孢、肤色镰孢、硫色镰孢、圆镰孢、拟丝孢镰孢或镶片镰孢;腐质霉属,优选特异腐质霉或疏棉状腐质霉;或木霉属,优选哈茨木霉、康宁木霉、长枝木霉、里氏木霉或绿色木霉。
可以依照本领域内已知的方法制备多肽组合物,并且可以是液体或干组合物的形式。例如,所述多肽组合物可以是颗粒(granulate)或微粒(microgranulate)的形式。可以依照本领域内已知方法将包括于所述组合物中的多肽稳定化。
下面给出的是本发明多肽组合物的优选的用途的实例。本发明的多肽组合物的剂量和使用该组合物的其它条件可以基于本领域内已知的方法确定。
纤维素材料的加工
本发明还涉及降解或转化纤维素材料的方法,其包括:在存在本发明的具有β-葡糖苷酶活性的多肽的情况下,用酶组合物处理纤维素材料。在一个优选方面,所述方法还包括回收已降解或转化的纤维素材料。
本发明还涉及产生发酵产物的方法,其包括:(a)在存在本发明的具有β-葡糖苷酶活性的多肽的条件下,用酶组合物糖化纤维素材料;(b)用一种或多种(几种)发酵微生物发酵经糖化的纤维素材料以产生发酵产物;和(c)从发酵回收发酵产物。
本发明还涉及发酵纤维素材料的方法,其包括:用一种或多种(几种)发酵微生物发酵纤维素材料,其中在存在具有β-葡糖苷酶活性的多肽的条件下,用酶组合物糖化纤维素材料。在一个优选的方面,纤维素材料的发酵产生发酵产物。在另一个优选的方面,方法进一步包括从发酵回收发酵产物。
本发明的方法可以用于将纤维素材料糖化成可发酵糖,并且将可发酵糖转化成很多有用的物质,例如燃料、饮用乙醇和/或发酵产物(例如酸、醇、酮、气体等)。从纤维素材料产生期望的发酵产物通常涉及预处理、酶水解(糖化)和发酵。
根据本发明的纤维素材料的处理可以使用本领域的常规过程完成。此外,本发明的方法能使用经配置以依照发明操作的任何常规生物质处理设备进行。
水解(糖化)和发酵,分别或同时,包括但不限于,分离的水解和发酵(SHF)、同步糖化和发酵(SSF)、同步糖化和共发酵(SSCF)、混合的水解和发酵(HHF)、分离的水解和共发酵(SHCF)、混合的水解和共发酵(HHCF),和直接微生物转化(DMC)。SHF使用分离的处理步骤以首先将纤维素材料酶水解为可发酵糖,例如,葡萄糖,纤维二糖、纤维三糖和戊糖,然后将可发酵糖发酵成为乙醇。在SSF中,纤维素材料的酶水解和糖变为乙醇的发酵在一个步骤中组合(Philippidis,G.P.,1996,Cellulose bioconversion technology,于Handbook onBioethanol:Production and Utilization,Wyman,C.E编,Taylor & Francis,Washington,DC,179-212)。SSCF包括多种糖的共发酵(Sheehan,J.,和Himmel,M.,1999,Enzymes,energy and the environment:A strategic perspective on theU.S.Department of Energy’s research and development activities for bioethanol,Biotechnol.Prog.15:817-827)。HHF在同步糖化和水解步骤之外,还涉及单独的水解步骤,所述步骤可以在同一个反应器中进行。HHF过程中的步骤可以在不同的温度,即,高温酶糖化,然后在发酵菌株能够耐受的较低温度进行SSF。DMC在一个或多个(几个)步骤中组合了所有三个过程(酶产生、水解和发酵),其中使用相同的生物体产生用于将木素纤维素转化成可发酵糖并将可发酵糖转化成终产物的酶(Lynd,L.R.,Weimer,P.J.,van Zyl,W.H.,和Pretorius,I.S.,2002,Microbial cellulose utilization:Fundamentals andbiotechnology,Microbiol.Mol.Biol.Reviews 66:506-577)。在本文可以理解的是,本领域中任何已知的方法,包括预处理、酶水解(糖化)、发酵,或它们的组合,可用于实施本发明的方法。
常规设备包括补料批式搅拌反应器、批式搅拌反应器、具有超滤的连续流搅拌反应器和/或连续活塞流柱式反应器(Fernanda de Castilhos Corazza,Flávio Faria de Moraes,Gisella Maria Zanin和Ivo Neitzel,2003,Optimal controlin fed-batch reactor for the cellobiose hydrolysis,Acta Scientiarum.Technology25:33-38;Gusakov,A.V.,和Sinitsyn,A.P.,1985,Kinetics of the enzymatichydrolysis of cellulose:1.A mathematical model for a batch reactor process,Enz.Microb.Technol.7:346-352)、研磨反应器(Ryu,S.K.,和Lee,J.M.,1983,Bioconversion of waste cellulose by using an attrition bioreactor,Biotechnol.Bioeng.25:53-65),或者具有由电磁场引起的强烈搅拌的反应器(Gusakov,A.V.,Sinitsyn,A.P.,Davydkin,I.Y.,Davydkin,V.Y.,Protas,O.V.,1996,Enhancement of enzymatic cellulose hydrolysis using a novel type of bioreactorwith intensive stirring induced by electromagnetic field,Appl.Biochem.Biotechnol.56:141-153)。其它反应器类型包括:流化床、升流层(upflowblanket)、固定化和用于水解和/或发酵的挤出机型的反应器。
预处理。在本发明的方法的实施中,可以使用本领域已知的任何预处理过程破坏植物细胞壁的纤维素材料组分(Chandra等,2007,Substratepretreatment:The key to effective enzymatic hydrolysis of lignocellulosics?Adv.Biochem.Engin./Biotechnol.108:67-93;Galbe和Zacchi,2007,Pretreatment oflignocellulosic materials for efficient bioethanol production,Adv.Biochem.Engin./Biotechnol.108:41-65;Hendriks和Zeeman,2009,Pretreatments to enhance thedigestibility of lignocellulosic biomass,Bioresource Technol.100:10-18;Mosier等,2005,Features of promising technologies for pretreatment of lignocellulosicbiomass,Bioresource Technol.96:673-686;Taherzadeh和Karimi,2008,Pretreatment of lignocellulosic wastes to improve ethanol and biogas production :A review,Int.J.of Mol.Sci.9:1621-1651;Yang和Wyman,2008,Pretreatment:the key to unlocking low-cost cellulosic ethanol,Biofuels Bioproducts andBiorefining-Biofpr.2:26-40)。
纤维素材料也可以在预处理之前使用本领域中已知的方法进行粒度减小、预浸泡、润湿、洗涤或调节。
常规的预处理包括但不限于,蒸汽预处理(伴随或不伴随爆炸)、稀酸预处理、热水预处理、碱性预处理、石灰预处理、湿氧化、湿爆炸、氨纤维爆炸、有机溶剂预处理和生物预处理。其它预处理包括氨渗滤、超声、电穿孔、微波、超临界CO2、超临界H2O、臭氧和γ辐射预处理。
可以在水解和/或发酵之前预处理纤维素材料。预处理优选在水解前进行。或者,预处理可以与酶水解同时进行以释放可发酵糖,如葡萄糖、木糖和/或纤维二糖。在大多数情况下,预处理步骤本身使一些生物质转化成可发酵糖(甚至在不存在酶的情况下)。
蒸汽预处理。在蒸汽预处理中,加热纤维素材料以破坏植物细胞壁成分,包括木质素、半纤维素和纤维素,使酶可接触纤维素和其它部分,例如,半纤维素。将纤维素材料通过或穿过反应容器,其中注入蒸汽以增加温度至需要的温度和压力,并且在其中保持期望的反应时间。蒸汽预处理优选在140-230℃,更优选160-200℃,并且最优选170-190℃进行,其中最优的温度范围依赖于加入的任何化学催化剂。蒸汽预处理的停留时间优选1-15分钟,更优选3-12分钟,并且最优选4-10分钟,其中最优的停留时间依赖于温度范围和加入的任何化学催化剂。蒸汽预处理允许相对较高的固体加载量,使纤维素材料在预处理过程中通常仅仅变得潮湿。蒸汽预处理经常与预处理后的物质的爆炸放料(explosive discharge)组合,这称为蒸汽爆炸,即,快速闪变至大气压和物质的湍流,以通过破碎增加可接触的表面积(Duff和Murray,1996,Bioresource Technology 855:1-33;Galbe和Zacchi,2002,Appl.Microbiol.Biotechnol.59:618-628;美国专利申请No.20020164730)。在蒸汽预处理过程中,切割半纤维素乙酰基团,并且得到的酸自催化纤维素部分水解成单糖和寡糖。去除木质素至有限的程度。
经常在蒸汽预处理之前加入催化剂如H2SO4或SO2(通常0.3至3%w/w),其可减少时间,降低温度,增加回收率,并改进酶水解(Ballesteros等,2006,Appl.Biochem.Biotechnol.129-132:496-508;Varga等,2004,Appl.Biochem.Biotechnol.113-116:509-523;Sassner等.,2006,EnzymeMicrob.Technol.39:756-762)。
化学预处理:术语“化学处理“指能促进纤维素、半纤维素和/或木质素分离和/或释放的任何化学处理。合适的化学预处理步骤的实例包括例如稀酸预处理、石灰预处理、湿氧化、氨纤维/冷冻爆炸(AFEX)、氨渗滤(APR)和有机溶剂预处理。
在稀酸预处理中,将纤维素材料与稀酸(通常是H2SO4)和水混合以形成浆料,由蒸汽加热至期望的温度,并在一段停留时间后闪变至大气压。可以用很多反应器设计进行稀酸预处理,例如,活塞流式反应器、逆流反应器或连续逆流收缩床反应器(Duff和Murray,1996,supra;Schell等,2004,BioresourceTechnol.91:179-188;Lee等,1999,Adv.Biochem.Eng.Biotechnol.65:93-115)。
还可以使用碱性条件下的几种预处理方法。这些碱预处理包括,但不限于,石灰预处理、湿氧化、氨渗滤(APR)和氨纤维/冷冻爆炸(AFEX)。
用碳酸钙、氢氧化钠或氨,在85-150℃的低温进行石灰预处理,停留时间从1小时到几天(Wyman等,2005,Bioresource Technol.96:1959-1966;Mosier等,2005,Bioresource Technol.96:673-686)。WO 2006/110891、WO 2006/110899、WO 2006/110900和WO 2006/110901公开了使用氨的预处理方法。
湿法氧化是热预处理,通常在180-200℃进行5-15分钟,加入氧化剂如过氧化氢或过压氧(Schmidt和Thomsen,1998,Bioresource Technol.64:139-151;Palonen等,2004,Appl.Biochem.Biotechnol.117:1-17;Varga等,2004,Biotechnol.Bioeng.88:567-574;Martin等,2006,J.Chem.Technol.Biotechnol.81:1669-1677)。预处理以优选1-40%干物质,更优选2-30%干物质,并且最优性5-20%干物质进行,并且由于加入碱如碳酸钠,初始pH经常会增加。
湿法氧化预处理方法的修改方法,称为湿爆炸(湿氧化和蒸汽爆炸的组合),能够处理高达30%的干物质。在湿爆炸中,在预处理过程中,在一定的停留时间后引入氧化剂。然后通过闪变至大气压而结束预处理(WO 2006/032282)。
氨纤维爆炸(AFEX)涉及在温和温度如90-100℃和高压如17-20bar,用液体或气体氨将纤维素材料处理5-10分钟,其中干物质含量可以高达60%(Gollapalli等,2002,Appl.Biochem.Biotechnol.98:23-35;Chundawat等,2007,Biotechnol.Bioeng.96:219-231;A1izadeh等,2005,Appl.Biochem.Biotechnol.121:1133-1141;Teymouri等,2005,Bioresource Technol.96:2014-2018)。AFEX预处理导致纤维素解聚,和半纤维素的部分水解。木质素-糖复合物得到切割。
有机溶剂预处理通过用含水乙醇(40-60%乙醇)在160-200℃提取30-60分钟而将纤维素材料去木质素化(Pan等,2005,Biotechnol.Bioeng.90:473-481;Pan等,2006,Biotechnol.Bioeng.94:851-861;Kurabi等,2005,Appl.Biochem.Biotechnol.121:219-230)。经常加入硫酸作为催化剂。在有机溶剂预处理中,去除大部分半纤维素。
合适的预处理方法的其他实例如Schell等,2003,Appl.Biochem andBiotechn.Vol.105-108:69-85,和Mosier等,2005,Bioresource Technology 96:673-686,和美国专利公开申请2002/0164730所述。
在一个方面,化学预处理优选作为酸处理,并且更优选作为连续稀酸和/或弱酸(mild acid)处理进行。酸通常是硫酸,但也可以使用其它酸,如乙酸、柠檬酸、硝酸、磷酸、酒石酸、琥珀酸、氯化氢或其混合物。弱酸处理在优选1-5,更优选1-4,并且最优选1-3的pH范围内进行。在一个方面,酸浓度在优选0.01至20wt%酸,更优选0.05至10wt%酸,甚至更优选0.1至5wt%酸,并且最优选0.2至2.0wt%酸的范围内。酸与纤维素材料接触,并在优选160-220℃,和更优选165-195℃范围内的温度保持数秒到数分钟,例如1秒-60分钟的时间。
在另一个方面,预处理作为氨纤维爆炸步骤(AFEX预处理步骤)进行。
在另一个方面,预处理发生在含水浆料中。在优选的方面,在预处理过程中纤维素材料以优选10-80wt%,更优选20-70wt%,并且最优性30-60wt%,如约50wt%的量存在。预处理的纤维素材料可以不洗涤或者使用本领域任何已知的方法洗涤,例如,用水洗涤。
机械预处理:术语“机械预处理”指各种类型的磨制(grinding)或粉碎(milling)(例如,干磨、湿磨或振动球磨)。
物理预处理:术语“物理预处理”指促进纤维素、半纤维素和/或木质素从纤维素材料分离和/或释放的任何预处理。例如,物理预处理可涉及辐射(例如,微波辐射)、汽蒸/蒸汽爆炸、水热解(hydrothermolysis)和它们的组合。
物理预处理可以涉及高压和/或高温(蒸汽爆炸)。在一个方面,高压指优选约300至约600psi,更优选约350至约550psi,并且最优选约400至约500psi,如约450psi的压力。在另一个方面,高温指约100至300℃,优选约140至235℃的温度。在一个优选的方面,机械预处理在使用如上所定义的高温和高压的分批过程、蒸汽枪水解器系统,例如来自Sunds DefiratorAB,Sweden的Sunds Hydrolyzer中进行。
组合的物理和化学预处理:可以对纤维素材料进行物理和化学预处理。例如,预处理步骤可以涉及稀酸或弱酸处理和高的温度和/或压力处理。根据需要,可以顺序或同时进行物理和化学预处理。还可以包括机械预处理。
因此,在一个优选的方面,对纤维素材料进行机械、化学或物理预处理,或者它们的任何组合,以促进纤维素、半纤维素和/或木质素的分离和/或释放。
生物预处理:术语“生物预处理”指可以促进纤维素、半纤维素和/或木质素从纤维素材料分离和/或释放的任何生物预处理。生物预处理技术可以包括应用溶解木质素的微生物(参见,例如,Hsu,T.-A.,1996,Pretreatment of biomass,于Handbook on Bioethanol:Production and Utilization,Wyman,C.E编,Taylor&Francis,Washington,DC,179-212;Ghosh和Singh,1993,Physicochemical andbiological treatments for enzymatic/microbial conversion of lignocellulosic biomass,Adv.Appl.Microbiol.39:295-333;McMillan,J.D.,1994,Pretreating lignocellulosicbiomass:a review,于Enzymatic Conversion of Biomass for Fuels Production,Himmel,M.E.,Baker,J.O.,和Overend,R.P.,编,ACS Symposium Series 566,American Chemical Society,Washington,DC,第15章;Gong,C.S.,Cao,N.J.,Du,J.,和Tsao,G.T.,1999,Ethanol production from renewable resources,于Advances inBiochemical Engineering/Biotechnology,Scheper,T.,编,Springer-Verlag BerlinHeidelberg,Germany,65:207-241;Olsson和Hahn-Hagerdal,1996,Fermentation oflignocellulosic hydrolysates for ethanol production,Enz.Microb.Tech.18:312-331;和Vallander和Eriksson,1990,Production of ethanol from lignocellulosic materials:State ofthe art,Adv.Biochem.Eng./Biotechnol.42:63-95)。
糖化。在水解(也称作糖化)步骤中,将例如经预处理的纤维素材料水解以将纤维素和任选地也将半纤维素分解成可发酵糖,如葡萄糖、纤维二糖、木糖、木酮糖、阿拉伯糖、甘露糖、半乳糖和/或可溶的寡糖。水解由酶组合物以酶法在本发明具有β-葡糖苷酶活性的多肽的存在下进行。所述组合物可以进一步包含一种或多种(几种)半纤维素分解酶。组合物的酶还可以顺序加入。
酶水解优选在易于由本领域技术人员确定的条件下,在合适的含水环境中进行。在一个优选的方面,水解在适于酶的活性,即对于酶最优的条件下进行。水解可以以补料批式或连续的过程进行,其中将预处理的纤维素材料(底物)逐渐补入,例如,含酶的水解溶液中。
糖化通常在搅拌釜反应器或发酵罐中,在受控的pH、温度和混合条件下进行。合适的处理时间、温度和pH条件可以由本领域技术人员容易地确定。例如,糖化可以持续长达200小时,但是通常优选进行约12至约96小时,更优选约16至约72小时,并且最优选约24至约48小时。温度优选约25℃至约70℃,更优选约30℃至约65℃,并且最优选约40℃至约60℃,特别是约50℃。pH优选约3至约8,更优选约3.5至约7,并且最优选约4至约6,特别是约pH 5。干燥固体含量优选约5至约50wt%,更优选约10至约40wt%,并且最优选约20至约30wt%。
所述酶组合物优选包含具有纤维素分解活性和/或木聚糖降解活性。在一个方面,所述酶组合物包含一种或多种(几种)纤维素分解酶。在另一个方面,所述酶组合物包含一种或多种(几种)木聚糖降解酶。在另一个方面,所述酶组合物包含一种或多种(几种)纤维素分解酶和一种或多种(几种)木聚糖降解酶。
所述一种或多种(几种)纤维素分解酶优选选自下组:内切葡聚糖酶、纤维二糖水解酶和β-葡糖苷酶。所述一种或多种(几种)木聚糖降解酶优选选自下组:木聚糖酶、乙酰木聚糖酯酶、阿魏酸酯酶、阿拉伯呋喃糖苷酶、木糖苷酶和葡糖醛酸糖苷酶。
在另一个方面,所述酶组合物另外或甚至还包含具有纤维素分解增强活性的多肽(参见,例如WO 2005/074647、WO 2005/074656和WO 2007/089290)。在另一个方面,所述酶组合物可进一步或甚至进一步包含一种或多种(几种)其他酶活性以改善含纤维素材料的降解。其他优选的酶为半纤维素酶(例如α-D-葡糖醛酸糖苷酶、α-L-阿拉伯呋喃糖苷酶、内切甘露聚糖酶、β-甘露糖苷酶、α-半乳糖苷酶、内切-α-L-阿拉伯聚糖酶、β-半乳糖苷酶),糖酯酶(例如乙酰木聚糖酯酶、乙酰甘露聚糖酯酶、阿魏酸酯酶、香豆酸酯酶、葡糖醛酸酯酶(glucuronoylesterases)),果胶酶,蛋白酶,木质素水解酶(ligninolytic enzyme)(例如漆酶、锰过氧化物酶、木质素过氧化物酶、H2O2产生酶、氧化还原酶),棒曲霉素(expansin)、膨胀素(swollenin)或其组合。在本发明的方法中,其他酶可在发酵之前或之中添加,例如在糖化之中或在发酵微生物繁殖之中或之后添加。
所述酶组合物的一种或多种(几种)组分可为野生型蛋白、重组蛋白或野生型蛋白和重组蛋白的组合。举例而言,一种或多种(几种)组分可为细胞的天然蛋白,其用作宿主细胞以重组表达一种或多种(几种)酶组合物的其他组分。酶组合物的一种或多种(几种)组分可作为单组分产生,然后将其组合以形成酶组合物。所述酶组合物可为多组分和单组分蛋白制备物的组合。
用于本发明方法中的酶可为任何适用于本文中所述工艺的形式,如例如去除或未去除细胞的粗发酵液,含或不含细胞碎片的细胞裂解液,半纯化或纯化的酶制备物,或宿主细胞,作为酶的来源。所述酶组合物可为干粉或颗粒,无粉尘的颗粒,液体,稳定化液体或稳定化受保护的酶。液体酶制备物可根据确立的工艺,例如通过添加稳定剂如糖、糖醇或其他多元醇,和/或乳酸或其他有机酸来稳定化。
具有β-葡糖苷酶活性的酶和多肽的最适量取决于几个因素,其包括但不限于,组分纤维素分解酶的混合物、含纤维素底物、含纤维素底物的浓度、含纤维素底物的预处理、温度、时间、pH和包括发酵生物体(例如,同步糖化和发酵的酵母)。
在一个优选的方面,纤维素分解酶对于纤维素材料的有效量是约0.5至约50mg,优选约0.5至约40mg,更优选约0.5至约25mg,更优选约0.75至约20mg,更优选约0.75至约15mg,甚至更优选约0.5至约10mg,并且最优选约2.5至约10mg每g纤维素材料。
在另一个优选的方面,具有β-葡糖苷酶活性的多肽对于纤维素材料的有效量是约0.01至约50.0mg,优选约0.01至约40mg,更优选约0.01至约30mg,更优选约0.01至约20mg,更优选约0.01至约10mg,更优选约0.01至约5mg,更优选约0.025至约1.5mg,更优选约0.05至约1.25mg,更优选约0.075至约1.25mg,更优选约0.1至约1.25mg,甚至更优选约0.15至约1.25mg,并且最优选约0.25至约1.0mg每g纤维素材料。
在另一个优选的方面,具有β-葡糖苷酶活性的多肽对于纤维素分解酶的有效量是约0.005至约1.0g,优选约0.01至约1.0g,更优选约0.15至约0.75g,更优选约0.15至约0.5g,更优选约0.1至约0.5g,甚至更优选约0.1至约0.5g,并且最优选约0.05至约0.2g每g纤维素分解酶。
酶可源自或获得自任何合适的来源,包括细菌、真菌、酵母、植物或哺乳动物来源。术语“获得”在本文中意指该酶可从天然产生该酶作为天然酶的生物分离。术语“获得”在本文中还意指该酶可在宿主生物中使用本文中所述的方法重组产生,其中经重组产生的酶对于宿主生物是天然的或异源的,或具有修饰的氨基酸序列,例如,具有一个或多个(几个)缺失、插入和/或取代的氨基酸,即重组产生的酶,其为天然氨基酸序列的片段和/或突变体或通过本领域已知的氨基酸改组方法产生的酶。天然酶的含义中涵盖的是天然变体,而外来酶的含义中涵盖的是重组(如通过定位诱变或重排)获得的变体
具有纤维素分解酶活性或木聚糖降解活性的多肽可以是细菌多肽。例如,所述多肽可以是革兰氏阳性细菌多肽如芽孢杆菌属(Bacillus)、链球菌属(Streptococcus)、链霉菌属(Streptomyces)、葡萄球菌属(Staphylococcus)、肠球菌属(Enterococcus)、乳杆菌属(Lactobacillus)、乳球菌属(Lactococcus)、梭菌属(Clostridium)、地芽孢杆菌属(Geobacillus)或海洋芽孢杆菌属(Oceanobacillus)多肽,所述多肽具有纤维素分解酶活性或木聚糖降解活性;或革兰氏阴性细菌多肽,如大肠杆菌、假单胞菌属(Pseudomonas)、沙门氏菌属(Salmonella)、弯曲杆菌属(Campylobacter)、螺杆菌属(Helicobacter)、黄杆菌属(Flavobacterium)、梭杆菌属(Fusobacterium)、泥杆菌属(Ilyobacter)、奈瑟氏菌属(Neisseria)或脲原体属(Ureaplasma)多肽,所述多肽具有纤维素分解酶活性或木聚糖降解活性。
在一个优选的方面,所述多肽是具有纤维素分解活性或木聚糖降解活性的嗜碱芽孢杆菌、解淀粉芽孢杆菌、短芽孢杆菌、环状芽孢杆菌、克劳氏芽孢杆菌、凝结芽孢杆菌、坚强芽孢杆菌、灿烂芽孢杆菌、迟缓芽孢杆菌、地衣芽孢杆菌、巨大芽孢杆菌、短小芽孢杆菌、嗜热脂肪芽孢杆菌、枯草芽孢杆菌或苏云金芽孢杆菌多肽。
在另一个优选的方面,所述多肽是具有纤维素分解活性或木聚糖降解活性的似马链球菌、酿脓链球菌、乳房链球菌或马链球菌兽瘟亚种多肽。
在另一个优选的方面,所述多肽是具有纤维素分解活性或木聚糖降解活性的不产色链霉菌、除虫链霉菌、天蓝链霉菌、灰色链霉菌或浅青紫链霉菌多肽。
具有纤维素分解活性或木聚糖降解活性的多肽也可以是真菌多肽,并且更优选酵母多肽如假丝酵母属、克鲁维酵母属、毕赤酵母属、酵母属、裂殖酵母属或西洋蓍霉属多肽,其具有纤维素分解活性或木聚糖降解活性;或更优选丝状真菌多肽如枝顶孢霉属、伞菌属、链格孢属、曲霉属、短梗霉属、Botryospaeria、拟蜡菌属、Chaetomidium、金孢子菌属、Claviceps、Cochliobolus、鬼伞属、Coptotermes、棒囊壳属、隐丛赤壳菌属、隐球菌属、色二孢属、黑耳属、Filibasidium、镰孢属、赤霉属、全鞭毛虫属、腐质霉属、耙齿菌属、蘑菇属、Leptospaeria、梨孢菌属、Melanocarpus、多孔菌属、毛霉属、毁丝霉属、新考玛脂霉属、脉孢菌属、拟青霉属、青霉属、平革菌属、瘤胃壶菌属、Poitrasia、假黑盘菌属、Pseudotrichonympha、根毛霉属、裂褶菌属、柱顶孢属、踝节菌属、嗜热子囊菌属、梭孢壳属、弯颈霉属、木霉属、长毛盘菌属、轮枝孢属、包脚菇属或炭角菌属多肽,其具有纤维素分解活性或木聚糖降解活性。
在一个优选的方面,所述多肽是具有纤维素分解活性或木聚糖降解活性的卡尔酵母、酿酒酵母、糖化酵母、道格拉氏酵母、克鲁弗酵母、诺地酵母或卵形酵母多肽。
在另一个优选的方面,所述多肽是具有纤维素分解活性或木聚糖降解活性的解纤维枝顶孢霉、棘孢曲霉、泡盛曲霉、烟曲霉、臭曲霉、日本曲霉、构巢曲霉、黑曲霉、米曲霉、嗜角质金孢子菌、Chrysosporium lucknowense、热带金孢子菌、Chrysosporium merdarium、Chrysosporium inops、毡金孢子菌、Chrysosporium queenslandicum、Chrysosporim zonatum、杆孢状镰孢、禾谷镰孢、库威镰孢、大刀镰孢、禾本科镰孢、禾赤镰孢、异孢镰孢、合欢木镰孢、尖镰孢、多枝镰孢、粉红镰孢、接骨木镰孢、肤色镰孢、拟分枝孢镰孢、硫色镰孢、圆镰孢、拟丝孢镰孢、镶片镰孢、灰腐质霉、特异腐质霉、疏棉状腐质霉、白耙齿菌、米黑毛霉、嗜热毁丝霉、粗糙脉孢菌、绳状青霉、产紫青霉、黄孢平革菌、Thielavia achromatica、Thielavia albomyces、Thielavia albopilosa、澳洲梭孢壳、Thielavia fimeti、小孢梭孢壳、卵孢梭孢壳、Thielavia peruviana、瘤孢梭孢壳、毛梭孢壳、Thielavia subthermophila、土生梭孢霉、哈茨木霉、康宁木霉、长枝木霉、里氏木霉、绿色木霉或Trichophaea saccata多肽。
还可以使用具有纤维素分解酶活性或木聚糖降解活性的多肽经化学修饰或蛋白质工程改造的突变体。
所述纤维素分解酶组合物的一种或多种(几种)组分可以是重组组分,亦即,通过克隆编码所述单独组分的DNA序列并随后用该DNA序列转化细胞并在宿主中表达(参见,例如,WO91/17243和WO91/17244)产生。所述宿主优选是异源宿主(酶对宿主是异源的),但该宿主在一定条件下也可以是同源宿主(酶对宿主是同源的)。单组分纤维素分解蛋白还可以通过从发酵液中提纯这样的蛋白质来制备。
适用于本发明的商业的纤维素分解蛋白制备物的实例包括,例如,CELLICTMCtec(Novozymes A/S)、CELLUCLASTTM(Novozymes A/S)、NOVOZYMTM 188(NovozymesA/S)、CELLUZYMETM(NovozymesA/S)、CEREFLOTM(NovozymesA/S)和ULTRAFLOTM(NovozymesA/S),ACCELERASETM(Genencor Int.)、LAMINEXTM(Genencor Int.)、SPEZYMETM CP(Genencor Int.),ROHAMENTTM7069W ( GmbH),
GmbH), LDI(Dyadic International,Inc.)、
LDI(Dyadic International,Inc.)、 LBR(Dyadic International,Inc.)或
LBR(Dyadic International,Inc.)或 150L(DyadicInternational,Inc.)。所述纤维素酶以固体的约0.001到约5.0wt.%,更优选固体的0.025到约4.0wt.%,且最优选固体的约0.005到约2.0wt.%的有效量添加。所述纤维素酶以固体的约0.001到约5.0wt.%,更优选固体的0.025到约4.0wt.%,且最优选固体的约0.005到约2.0wt.%的有效量添加。
150L(DyadicInternational,Inc.)。所述纤维素酶以固体的约0.001到约5.0wt.%,更优选固体的0.025到约4.0wt.%,且最优选固体的约0.005到约2.0wt.%的有效量添加。所述纤维素酶以固体的约0.001到约5.0wt.%,更优选固体的0.025到约4.0wt.%,且最优选固体的约0.005到约2.0wt.%的有效量添加。
可以用于本发明的方法的细菌内切葡聚糖酶的实例包括但不仅限于,解纤维热酸菌(Acidothermus cellulolyticus)内切葡聚糖酶(WO 91/05039;WO93/15186;美国专利5,275,944;WO 96/02551;美国专利5,536,655,WO00/70031,WO 05/093050);Thermobifida fusca内切葡聚糖酶III(WO05/093050);和Thermobifida fusca内切葡聚糖酶V(WO 05/093050)。
可以用于本发明的方法的真菌内切葡聚糖酶的实例包括但不仅限于,里氏木霉内切葡聚糖酶I(Penttila等,1986,Gene 45:253-263;GENBANKTM登录号M15665);里氏木霉内切葡聚糖酶II(Saloheimo等,1988,Gene 63:11-22;GENBANKTM登录号M19373);里氏木霉内切葡聚糖酶III(Okada等,1988,Appl.Environ.Microbiol.64:555-563;GENBANKTM登录号AB003694);里氏木霉内切葡聚糖酶IV(Saloheimo等,1997,Eur.J.Biochem.249:584-591;GENBANKTM登录号Y11113);以及里氏木霉内切葡聚糖酶V(Saloheimo等,1994,Molecular Microbiology 13:219-228;GENBANKTM登录号Z33381);棘孢曲霉内切葡聚糖酶(Ooi等,1990,Nucleic Acids Research18:5884);川地曲霉(Aspergillus kawachii)内切葡聚糖酶(Sakamoto等,1995,Current Genetics 27:435-439);胡萝卜软腐欧文氏菌(Erwinia carotovara)内切葡聚糖酶(Saarilahti等,1990,Gene 90:9-14);尖镰孢内切葡聚糖酶(GENBANKTM登录号L29381);灰腐质霉thermoidea变种内切葡聚糖酶(GENBANKTM登录号AB003107);Melanocarpus albomyces内切葡聚糖酶(GENBANKTM登录号MAL515703);粗糙脉孢菌内切葡聚糖酶(GENBANKTM登录号XM 324477);特异腐质霉内切葡聚糖酶V(SEQ ID NO:4);嗜热毁丝霉CBS 117.65内切葡聚糖酶(SEQ IDNO:6);担子菌纲(basidiomycete)CBS 495.95内切葡聚糖酶(SEQ ID NO:8);担子菌纲CBS 494.95内切葡聚糖酶(SEQ ID NO:10);土生梭孢霉NRRL 8126CEL6B内切葡聚糖酶(SEQ ID NO:12);土生梭孢霉NRRL 8126CEL6C内切葡聚糖酶(SEQ ID NO:14);土生梭孢霉NRRL 8126CEL7C内切葡聚糖酶(SEQ ID NO:16);土生梭孢霉NRRL 8126CEL7E内切葡聚糖酶(SEQ ID NO:18);土生梭孢霉NRRL 8126CEL7F内切葡聚糖酶(SEQ ID NO:20);Cladorrhinum foecundissimum ATCC 62373CEL7A内切葡聚糖酶(SEQ ID NO:22);以及里氏木霉菌株VTT-D-80133内切葡聚糖酶(SEQ ID NO:24;GENBANKTM登录号M15665)。上述SEQ ID NO:4、SEQ ID NO:6、SEQ ID NO:8、SEQ ID NO:10、SEQ ID NO:12、SEQ ID NO:14、SEQ ID NO:16、SEQ IDNO:18、SEQ ID NO:20、SEQ ID NO:22和SEQ ID NO:24的内切葡聚糖酶分别由SEQ ID NO:3、SEQ ID NO:5、SEQ ID NO:7、SEQ ID NO:9、SEQ IDNO:11、SEQ ID NO:13、SEQ ID NO:15、SEQ ID NO:17、SEQ ID NO:19、SEQ ID NO:21、SEQ ID NO:23的成熟多肽编码序列编码。
可用于本发明的方法的纤维二糖水解酶的示例包括但不仅限于,里氏木霉纤维二糖水解酶I(SEQ ID NO:26);里氏木霉纤维二糖水解酶II(SEQ IDNO:28);特异腐质霉纤维二糖水解酶I(SEQ ID NO:30)、嗜热毁丝霉纤维二糖水解酶II(SEQ ID NO:32和SEQ ID NO:34)、土生梭孢霉纤维二糖水解酶II(CEL6A)(SEQ ID NO:36)、嗜热毛壳菌(Chaetomium thermophilum)纤维二糖水解酶I(SEQ ID NO:38)以及嗜热毛壳菌纤维二糖水解酶II(SEQ ID NO:40)。上述SEQ ID NO:24、SEQ ID NO:26、SEQ ID NO:28、SEQ ID NO:30、SEQ ID NO:32、SEQ ID NO:34、SEQ ID NO:36、SEQ ID NO:38和SEQ IDNO:40的纤维二糖水解酶分别由SEQ ID NO:23、SEQ ID NO:25、SEQ ID NO:27、SEQ ID NO:29、SEQ ID NO:31、SEQ ID NO:33、SEQ ID NO:35、SEQID NO:37和SEQ ID NO:39的成熟多肽编码序列编码。
可用于本发明的方法的β-葡糖苷酶的实例包括但不仅限于米曲霉β-葡糖苷酶(SEQ ID NO:42);烟曲霉β-葡糖苷酶(SEQ ID NO:44);巴西青霉(Penicilliumbrasilianum)IBT 20888β-葡糖苷酶(SEQ ID NO:46);黑曲霉β-葡糖苷酶(SEQ IDNO:48);以及棘孢曲霉β-葡糖苷酶(SEQ ID NO:50)。上述SEQ ID NO:40、SEQID NO:42、SEQ ID NO:44、SEQ ID NO:46、SEQ ID NO:48和SEQ ID NO:50的β-葡糖苷酶分别由SEQ ID NO:39、SEQ ID NO:41、SEQ ID NO:43、SEQ IDNO:45、SEQ ID NO:47和SEQ ID NO:49的成熟多肽编码序列编码。
具有β-葡糖苷酶活性的米曲霉多肽可根据WO 2002/095014获取。具有β-葡糖苷酶活性的烟曲霉多肽可根据WO 2005/047499获取。具有β-葡糖苷酶活性的巴西青霉多肽可根据WO 2007/019442获取。具有β-葡糖苷酶活性的黑曲霉多肽可根据Dan等,2000,J.Biol.Chem.275:4973-4980获取。具有β-葡糖苷酶活性的棘孢曲霉多肽可根据Kawaguchi等,1996,Gene 173:287-288获取。
所述β-葡糖苷酶可以是融合蛋白。在一个方面,所述β-葡糖苷酶是SEQID NO:52的米曲霉β-葡糖苷酶变体BG融合蛋白或SEQ ID NO:54的米曲霉β-葡糖苷酶融合蛋白。在另一个方面,所述米曲霉β-葡糖苷酶变体BG融合蛋白由SEQ ID NO:51的多核苷酸编码,或所述米曲霉β-葡糖苷酶融合蛋白由SEQ ID NO:53的多核苷酸编码。
其它内切葡聚糖酶、纤维二糖水解酶和β-葡糖苷酶使用根据Henrissat B.,1991,A classification of glycosyl hydrolases based on amino-acid sequencesimilarities,Biochem.J.280:309-316和Henrissat B.和Bairoch A.,1996,Updating the sequence-based classification of glycosyl hydrolases,Biochem.J.316:695-696的分类公开于许多糖基水解酶家族中。
其它可用于本发明的纤维素分解酶描述于EP 495,257、EP 531,315、EP531,372、WO 89/09259、WO 94/07998、WO 95/24471、WO 96/11262、WO96/29397、WO 96/034108、WO 97/14804、WO 98/08940、WO 98/012307、WO98/13465、WO 98/015619、WO 98/015633、WO 98/028411、WO 99/06574、WO 99/10481、WO 99/025846、WO 99/025847、WO 99/031255、WO2000/009707、WO 2002/050245、WO 2002/0076792、WO 2002/101078、WO2003/027306、WO 2003/052054、WO 2003/052055、WO 2003/052056、WO2003/052057、WO 2003/052118、WO 2004/016760、WO 2004/043980、WO2004/048592、WO 2005/001065、WO 2005/028636、WO 2005/093050、WO2005/093073、WO 2006/074005、WO 2006/117432、WO 2007/071818、WO2007/071820、WO 2008/008070、WO 2008/008793、美国专利No.4,435,307、美国专利No.5,457,046、美国专利No.5,648,263、美国专利No.5,686,593、美国专利No.5,691,178、美国专利No.5,763,254以及美国专利No.5,776,757。
在本发明的方法中,可使用任何具有纤维素分解增强活性的多肽。
在一个方面,所述具有纤维素分解增强活性的多肽包含下述基序:
[ILMV]-P-X(4,5)-G-X-Y-[ILMV]-X-R-X-[EQ]-X(4)-[HNQ]和[FW]-[TF]-K-[AIV],
其中X为任意氨基酸,X(4,5)为在4或5个连续位置的任意氨基酸,而X(4)是在4个连续位置的任意氨基酸。
具有上述所示的基序的多肽可进一步包含:
H-X(1,2)-G-P-X(3)-[YW]-[AILMV],
[EQ]-X-Y-X(2)-C-X-[EHQN]-[FILV]-X-[ILV]或
H-X(1,2)-G-P-X(3)-[YW]-[AILMV]和[EQ]-X-Y-X(2)-C-X-[EHQN]-[FILV]-X-[ILV],
其中X为任意氨基酸,X(1,2)为在1个位置或2个连续位置的任意氨基酸,X(3)为3个连续位置的任意氨基酸,而X(2)为2个连续位置的任意氨基酸。在上述基序中,采用公认的IUPAC单字母氨基酸缩写。
在一个优选的方面,所述具有纤维素分解增强活性的多肽进一步包含H-X(1,2)-G-P-X(3)-[YW]-[AILMV]。在另一个优选的方面,具有纤维素分解增强活性的分离的多肽包含[EQ]-X-Y-X(2)-C-X-[EHQN]-[FILV]-X-[ILV]。在另一个优选的方面,具有纤维素分解增强活性的多肽进一步包含H-X(1,2)-G-P-X(3)-[YW]-[AILMV]和[EQ]-X-Y-X(2)-C-X-[EHQN]-[FILV]-X-[ILV]。
在第二个方面,所述具有纤维素分解增强活性的多肽包含下述基序:
[ILMV]-P-x(4,5)-G-x-Y-[ILMV]-x-R-x-[EQ]-x(3)-A-[HNQ],
其中x为任意氨基酸,x(4,5)为在4或5个连续位置的任意氨基酸,而x(3)为3个连续位置的任意氨基酸。在上述基序中,采用公认的IUPAC单字母氨基酸缩写。
具有纤维素分解增强活性的分离的多肽的实例包括具有纤维素分解增强活性的土生梭孢霉(SEQ ID NO:56,SEQ ID NO:58,SEQ ID NO:60,SEQ ID NO:62,SEQ ID NO:64或SEQ ID NO:66的成熟多肽);桔橙热子囊菌(SEQ ID NO:68的成熟多肽);或里氏木霉(SEQ ID NO:70的成熟多肽)。上述具有SEQ ID NO:56、SEQ ID NO:58、SEQ ID NO:60、SEQ ID NO:62、SEQ ID NO:64、SEQ IDNO:66、SEQ ID NO:68和SEQ ID NO:70的具有纤维素分解增强活性的多肽分别由SEQ ID NO:55、SEQ ID NO:57、SEQ ID NO:59、SEQ ID NO:61、SEQ IDNO:63、SEQ ID NO:65、SEQ ID NO:67和SEQ ID NO:69的成熟多肽编码序列编码。参见,例如WO2005/074647、WO2005/074656和WO2007/089290。
适用于本发明的商业性木聚糖降解酶制备物的实例包括,例如SHEARZYMETM(Novozymes A/S)、CELLICTM Htec(Novozymes A/S)、(Novozymes A/S)、 (Novozymes A/S)、
(Novozymes A/S)、 HC(Novozymes A/S)、Xylanase(Genencor)、
HC(Novozymes A/S)、Xylanase(Genencor)、 TX-200A(AB Enzymes)、HSP 6000Xylanase(DSM)、DEPOLTM333P(Biocatalysts Limit,Wales,UK)、DEPOLTM 740L.(Biocatalysts Limit,Wales,UK)和DEPOLTM 762P(Biocatalysts Limit,Wales,UK)。
TX-200A(AB Enzymes)、HSP 6000Xylanase(DSM)、DEPOLTM333P(Biocatalysts Limit,Wales,UK)、DEPOLTM 740L.(Biocatalysts Limit,Wales,UK)和DEPOLTM 762P(Biocatalysts Limit,Wales,UK)。
可用于本发明方法的木聚糖酶的实例包括但不限于棘孢曲霉(Aspergillusaculeatus)木聚糖酶(GeneSeqP:AAR63790;WO 94/21785)、烟曲霉(Aspergillusfumigatus)木聚糖酶(WO 2006/078256)和土生梭孢霉(Thielavia terrestris)NRRL 8126木聚糖酶(WO 2009/079210)。
可用于本发明方法的β-木糖苷酶的实例包括但不限于里氏木霉(Trichoderma reesei)β-木糖苷酶(UniProtKB/TrEMBL登录号Q92458),埃默森踝节菌(Talaromyces emersonii)(SwissProt登录号Q8X212)和粗糙脉孢菌(Neurospora crassa)(SwissProt登录号Q7SOW4)。
可用于本发明方法的乙酰木聚糖酯酶的实例包括但不限于红褐肉座菌(Hypocrea jecorina)乙酰木聚糖酯酶(WO 2005/001036)、粗糙脉孢菌乙酰木聚糖酯酶(UniProt登录号q7s259)、土生梭孢霉NRRL 8126乙酰木聚糖酯酶(WO2009/042846)、球毛壳菌(Chaetomium globosum)乙酰木聚糖酯酶(Uniprot登录号Q2GWX4)、细丽毛壳菌(Chaetomium gracile)乙酰木聚糖酯酶(GeneSeqP登录号AAB82124)、颖枯壳针孢(Phaeosphaeria nodorum)乙酰木聚糖酯酶(Uniprot登录号Q0UHJ1)和特异腐质霉(Humicola insolens)DSM 1800乙酰木聚糖酯酶(WO 2009/073709)。
可用于本发明方法的阿魏酸酯酶的实例包括但不限于特异腐质霉DSM 1800阿魏酸酯酶(WO 2009/076122)、粗糙脉孢菌阿魏酸酯酶(UniProt登录号Q9HGR3)和费希新萨托菌(Neosartorya fischer)阿魏酸酯酶(UniProt登录号A1D9T4)。
可用于本发明方法的阿拉伯呋喃糖苷酶的实例包括但不限于特异腐质霉(Humicola insolens)DSM 1800阿拉伯呋喃糖苷酶(WO 2009/073383)和黑曲霉(Aspergillus niger)阿拉伯呋喃糖苷酶(GeneSeqP登录号AAR94170)。
可用于本发明方法的α-葡糖醛酸糖苷酶的实例包括但不限于棒曲霉(Aspergillus clavatus)α-葡糖醛酸糖苷酶(UniProt登录号alcc12)、里氏木霉(Trichoderma reesei)α-葡糖醛酸糖苷酶(Uniprot登录号Q99024)、埃默森踝节菌(Talaromyces emersonii)α-葡糖醛酸糖苷酶(UniProt登录号Q8X211)、黑曲霉(Aspergillus niger)α-葡糖醛酸糖苷酶(Uniprot登录号Q96WX9)、土曲霉(Aspergillus terreus)α-葡糖醛酸糖苷酶(SwissProt登录号Q0CJP9)和烟曲霉(Aspergillus fumigatus)α-葡糖醛酸糖苷酶(SwissProt登录号Q4WW45)。
用于本发明方法的酶和蛋白可通过在含有合适碳源和氮源和无机盐,使用本领域已知方法(参见,例如Bennett,J.W.和LaSure,L.(编),More GeneManipulations in Fungi,Academic Press,CA,1991)的营养培养基上发酵上述指出的微生物菌株来产生。合适的培养基可从供应商获得,或可根据已公开组合物制备(例如美国典型培养物保藏中心的目录)。适于生长和酶产生的温度范围和其他条件在本领域是已知的(参见,例如Bailey,J.E.和Ollis,D.F.,BiochemicalEngineering Fundamentals,McGraw-Hill Book Company,NY,1986)。
所述发酵可以是任何其结果为酶表达或分离的培养细胞的方法。因此,发酵可以理解为包括在合适的培养基中并在允许所述酶得以表达或分离的条件下进行的摇瓶培养,或在实验室或工业发酵罐中的小-或大规模发酵(包括连续、分批、补料分批或固态发酵)。通过上述方法产生的所得的酶可从发酵培养基回收并通过常规方法纯化。
发酵。可通过一种或多种(几种)能将糖直接或间接发酵成所需发酵产物的发酵微生物发酵自经预处理和水解的纤维素材料获得的可发酵糖。“发酵”或“发酵方法”指任何发酵方法或包含发酵步骤的任何方法。发酵方法还包括用于消费品醇工业(例如,啤酒和葡萄酒)、乳品业(例如,发酵乳产品)、皮革业和烟草业的发酵方法。发酵条件依赖于期望的发酵产物和发酵生物体,并且能由本领域的技术人员容易地确定。
在发酵步骤中,作为预处理和酶水解步骤的结果从纤维素材料释放的糖,通过发酵生物体(如酵母)发酵成为产物,例如,乙醇。如上所述,水解(糖化)和发酵可以是单独或同时的。
在本发明的发酵步骤中可以使用任何合适的经水解的纤维素材料。通常根据所需发酵产品(即,要从发酵获得的物质)和使用的方法来选择所述材料,如本领域中所公知的。
术语“发酵培养基”在本文中可理解为指加入发酵微生物之前的培养基,如,由糖化过程产生的培养基,以及同步的糖化和发酵方法(SSF)中使用的培养基。
“发酵微生物”指适用于理想的发酵方法产生发酵产物的任何微生物,包括细菌和真菌生物体。发酵生物体可以是C6和/或C5发酵生物体,或它们的组合。C6和C5发酵生物体均在本领域公知。合适的发酵微生物能将糖(如葡萄糖、木糖、木酮糖、阿拉伯糖、麦芽糖、甘露糖、半乳糖或寡糖)直接或间接地发酵(即,转化)成所需的发酵产品。
可产生乙醇的细菌和真菌发酵生物体的实例如Lin等,2006,Appl.Microbiol.Biotechnol.69:627-642所述。
能发酵C6糖的发酵微生物的实例包括细菌和真菌生物体,如酵母。优选的酵母包括酵母属菌种,优选酿酒酵母。
能发酵C5糖的发酵生物体的实例包括细菌和真菌生物体,如酵母。优选的C5发酵酵母包括毕赤酵母属,优选树干毕赤酵母(Pichia stipitis)的菌株,如树干毕赤酵母CBS 5773;假丝酵母属,优选博伊丁假丝酵母(Candidaboidinii)、芸薹假丝酵母(Candida brassicae)、休哈塔假丝酵母(Candidasheatae)、迪丹斯假丝酵母(Candida diddensii)、假热带假丝酵母(Candidapseudotropicalis)或产朊假丝酵母(Candida utilis)的菌株。
其它发酵生物体包括发酵单胞菌属(Zymomonas),如运动发酵单胞菌(Zymomonas mobilis);汉逊酵母属,如异常汉逊酵母(Hansenula anomala);克鲁维酵母属,如脆壁克鲁维酵母;裂殖酵母属,如粟酒裂殖酵母(S.pombe);和大肠杆菌的菌株,特别是已经经过遗传修饰而改进乙醇产量的大肠杆菌菌株。
在一个优选的方面,酵母是酵母属菌种。在一个更优选的方面,酵母是酿酒酵母。在另一个更优选的方面,酵母是糖化酵母(Saccharomycesdistaticus)。在另一个更优选的方面,酵母是葡萄汁酵母(Saccharomycesuvarum)。在另一个优选的方面,酵母是克鲁维酵母属。在另一个更优选的方面,酵母是马克斯克鲁维酵母(Kluyveromyces marxianus)。在另一个更优选的方面,酵母是脆壁克鲁维酵母。在另一个优选的方面,酵母是假丝酵母属。在另一个更优选的方面,酵母是博伊丁假丝酵母。在另一个更优选的方面,酵母是芸薹假丝酵母。在另一个更优选的方面,酵母是迪丹斯假丝酵母。在另一个更优选的方面,酵母是假热带假丝酵母。在另一个更优选的方面,酵母是产朊假丝酵母。在另一个优选的方面,酵母是棒孢酵母属(Clavispora)。在另一个更优选的方面,酵母是葡萄牙棒孢酵母(Clavispora lusitaniae)。在另一个更优选的方面,酵母是仙人掌棒孢酵母(Clavispora opuntiae)。在另一个优选的方面,酵母是管囊酵母属(Pachysolen)。在另一个更优选的方面,酵母是嗜鞣管囊酵母(Pachysolen tannophilus)。在另一个优选的方面,酵母是毕赤酵母属。在另一个更优选的方面,酵母是树干毕赤酵母。在另一个优选的方面,酵母是酒香酵母属(Bretannomyces)。在另一个更优选的方面,酵母是克劳森酒香酵母(Bretannomyces clausenii)(Philippidis,G.P.,1996,Cellulosebioconversion technology,于Handbook on Bioethanol:Production andUtilization,Wyman,C.E.编,Taylor&Francis,Washington,DC,179-212)。
能有效地将己糖和戊糖发酵成乙醇的细菌包括,例如,运动发酵单胞菌和热纤维梭菌(Clostridium thermocellum)(Philippidis,1996,见上文)。
在一个优选的方面,细菌是发酵单胞菌属。在更优选的方面,细菌是运动发酵单胞菌。在另一个优选的方面,细菌是梭菌属。在另一个更优选的方面,细菌是热纤维梭菌。
商业上可得到的适合乙醇产生的酵母包括,例如ETHANOL REDTM酵母(可从Red Star/Lesaffre,USA获得)、FALITM(可从Fleischmann’s Yeast,BurnsPhilp Food Inc.,USA的部门获得)、SUPERSTARTTM和THERMOSACCTM新鲜酵母(可从Ethanol Technology,WI,USA获得)、BIOFERMTM AFT和XR(可从NABC-North American Bioproducts Corporation,GA,USA获得)、GERTSTRANDTM(可从Gert Strand AB,Sweden获得)和FERMIOLTM(可从DSMSpecialties获得)。
在一个优选的方面,发酵微生物已经经过遗传修饰,提供发酵戊糖的能力,如利用木糖、利用阿拉伯糖和共同利用木糖和阿拉伯糖的微生物。
通过将异源基因克隆入多种发酵微生物已经构建了能将己糖和戊糖转化成乙醇(共发酵)的生物体(Chen和Ho,1993,Cloning and improving the expression ofPichia stipitis xylose reductase gene in Saccharomyces cerevisiae,Appl.Biochem.Biotechnol.39-40:135-147;Ho等,1998,Genetically engineered Saccharomycesyeast capable of effectively cofermenting glucose and xylose,Appl.Environ.Microbiol.64:1852-1859;Kotter和Ciriacy,1993,Xylose fermentation bySaccharomyces cerevisiae,Appl.Microbiol.Biotechnol.38:776-783;Walfridsson等,1995,Xylose-metabolizing Saccharomyces cerevisiae strains overexpressing theTKL 1and TAL1 genes encoding the pentose phosphate pathway enzymestransketolase and transaldolase,Appl.Environ.Microbiol.61:4184-4190;Kuyper等,2004,Minimal metabolic engineering of Saccharomyces cerevisiae for efficientanaerobic xylose fermentation:a proof of principle,FEMS Yeast Research 4:655-664;Beall等,1991,Parametric studies of ethanol production from xylose and other sugarsby recombinant Escherichia coli,Biotech.Bioeng.38:296-303;Ingram等,1998,Metabolic engineering of bacteria for ethanol production,Biotechnol.Bioeng.58:204-214;Zhang等,1995,Metabolic engineering ofa pentose metabolism pathway inethanologenic Zymomonas mobilis,Science 267:240-243;Deanda等,1996,Development of an arabinose-fermenting Zymomonas mobilis train by metabolicpathway engineering,Appl.Environ.Microbiol.62:4465-4470;WO 2003/062430,xylose isomerase)。
在一个优选的方面,经过遗传修饰的发酵微生物是酿酒酵母。在另一个优选的方面,经过遗传修饰的发酵微生物是运动发酵单胞菌。在另一个优选的方面,经过遗传修饰的发酵微生物是大肠杆菌。在另一个优选的方面,经过遗传修饰的发酵微生物是产酸克雷伯氏菌(Klebsiella oxytoca)。在另一个优选的方面,所述经遗传修饰的发酵微生物是克鲁维酵母菌种。
本领域中公知的是,上述生物体还能用于产生其它物质,如本文所述。
通常向降解的木素纤维素或水解物加入发酵微生物,并进行约8至约96小时,如约24至约60小时发酵。温度通常为约26℃至约60℃,特别是约32℃或50℃,并且在约pH 3至约pH 8,如约pH 4-5、6或7。
在一个优选的方面,对降解的纤维素材料施用酵母和/或另一种微生物,并进行约12至约96小时,如通常为24-60小时发酵。在一个优选的方面,温度优选为约20℃至约60℃,更优选约25℃至约50℃,并且最优选约32℃至约50℃,特别是约32℃或50℃,并且pH通常为约pH 3至约pH 7,优选约pH 4-7。然而,一些发酵生物体例如细菌,具有更高的最适发酵温度。酵母或另一种微生物优选以约105-1012,优选约107-1010,特别是约2x 108活细胞计数每ml发酵液的量施用。关于使用酵母进行发酵的进一步指导可以在例如“The AlcoholTextbook”(K.Jacques,T.P.Lyons和D.R.Kelsall编,Nottingham University Press,United Kingdom 1999)中找到,其通过提述并入本文。
对于乙醇生产,在发酵后蒸馏发酵的浆料以提取乙醇。根据本发明的方法获得的乙醇可以用作,例如燃料乙醇;饮料乙醇,即,中性饮料酒,或工业乙醇。
发酵刺激剂可以与本文所述的任何方法组合使用,以进一步改进发酵工艺,而且特定地,改进发酵微生物的性能,如,速率增加和乙醇得率。“发酵刺激剂”指用于发酵微生物(特别是酵母)生长的刺激剂。优选的用于生长的发酵刺激剂包括维生素和矿物质。维生素的实例包括多种维生素、生物素、泛酸(盐)、烟酸、内消旋肌醇(meso-inositol)、硫胺素、吡哆醇(pyridoxine)、对氨基苯甲酸、叶酸、核黄素和维生素A、B、C、D和E。参见,例如,Alfenore等,Improving ethanol production and viability of Saccharomyces cerevisiae by avitamin feeding strategy during fed-batch process,Springer-Verlag(2002),其通过提述并入本文。矿物质的实例包括能够提供营养物的矿物质和矿物质盐,所述营养物包括P、K、Mg、S、Ca、Fe、Zn、Mn和Cu。
发酵产物:发酵产物可以是源自发酵的任何物质。发酵产物可以是,不限于,醇(例如,阿拉伯醇、丁醇、乙醇、甘油、甲醇、1,3-丙二醇、山梨醇和木糖醇);有机酸(例如,乙酸、醋酮酸、己二酸、抗坏血酸、柠檬酸、2,5-二酮-D-葡糖酸、甲酸、反丁烯二酸、葡糖二酸、葡糖酸、葡糖醛酸、戊二酸、3-羟基丙酸、衣康酸、乳酸、苹果酸、丙二酸、草酸、草酰乙酸、丙酸、琥珀酸和木糖酸);酮(例如,丙酮);氨基酸(例如,天冬氨酸、谷氨酸、甘氨酸、赖氨酸、丝氨酸和苏氨酸);和气体(例如,甲烷、氢气(H2)、二氧化碳(CO2)和一氧化碳(CO))。发酵产物还可以是作为高价值产品的蛋白质。
在一个优选的方面,发酵产物是醇。可理解的是,术语“醇”包括包含一个或多个羟基基团的物质。在更优选的方面,所述醇是阿拉伯醇。在另一个更优选的方面,所述醇是丁醇。在另一个更优选的方面,所述醇是乙醇。在另一个更优选的方面,所述醇是甘油。在另一个更优选的方面,所述醇是甲醇。在另一个更优选的方面,所述醇是1,3-丙二醇。在另一个更优选的方面,所述醇是山梨醇。在另一个更优选的方面,所述醇是木糖醇。参见,例如,Gong,C.S.,Cao,N.J.,Du,J.,和Tsao,G.T.,1999,Ethanol production from renewable resources,于Advances in Biochemical Engineering/Biotechnology,Scheper,T.编,Springer-VerlagBerlin Heidelberg,Germany,65:207-241;Silveira,M.M.,和Jonas,R.,2002,Thebiotechnological production of sorbitol,Appl.Microbiol.Biotechnol.59:400-408;Nigam,P.,和Singh,D.,1995,Processes for fermentative production of xylitol-asugar substitute,Process Biochemistry 30(2):117-124;Ezeji,T.C.,Qureshi,N.和Blaschek,H.P.,2003,Production of acetone,butanol and ethanol by Clostridiumbeijerinckii BA101 and in situ recovery by gas stripping,World Journal ofMicrobiology and Biotechnology 19(6):595-603。
在另一个优选的方面,所述物质是有机酸。在另一个更优选的方面,所述有机酸是乙酸。在另一个更优选的方面,所述有机酸是醋酮酸。在另一个更优选的方面,所述有机酸是己二酸。在另一个更优选的方面,所述有机酸是抗坏血酸。在另一个更优选的方面,所述有机酸是柠檬酸。在另一个更优选的方面,所述有机酸是2,5-二酮-D-葡糖酸。在另一个更优选的方面,所述有机酸是甲酸。在另一个更优选的方面,所述有机酸是反丁烯二酸。在另一个更优选的方面,所述有机酸是葡糖二酸。在另一个更优选的方面,所述有机酸是葡糖酸。在另一个更优选的方面,所述有机酸是葡糖醛酸。在另一个更优选的方面,所述有机酸是戊二酸。在另一个优选的方面,所述有机酸是3-羟基丙酸。在另一个更优选的方面,所述有机酸是衣康酸。在另一个更优选的方面,所述有机酸是乳酸。在另一个更优选的方面,所述有机酸是苹果酸。在另一个更优选的方面,所述有机酸是丙二酸。在另一个更优选的方面,所述有机酸是草酸。在另一个更优选的方面,所述有机酸是丙酸。在另一个更优选的方面,所述有机酸是琥珀酸。在另一个更优选的方面,所述有机酸是木糖酸。参见,例如,Chen,R.,和Lee,Y.Y.,1997,Membrane-mediated extractive fermentation for lactic acidproduction from cellulosic biomass,Appl.Biochem.Biotechnol.63-65:435-448。
在另一个优选的方面,所述物质是酮。可理解的是术语“酮”涵盖含有一个或多个酮基的物质。在另一个更优选的方面,所述酮是丙酮。参见,例如,Qureshi和Blaschek,2003,见上文。
在另一个优选的方面,所述物质是氨基酸。在另一个更优选的方面,所述有机酸是天冬氨酸。在另一个更优选的方面,所述氨基酸是谷氨酸。在另一个更优选的方面,所述氨基酸是甘氨酸。在另一个更优选的方面,所述氨基酸是赖氨酸。在另一个更优选的方面,所述氨基酸是丝氨酸。在另一个更优选的方面,所述氨基酸是苏氨酸。参见,例如,Richard,A.,和Margaritis,A.,2004,Empirical modelingof batch fermentation kinetics for poly(glutamic acid)production and other microbialbiopolymers,Biotechnology and Bioengineering 87(4):501-515。
在另一个优选的方面,所述物质是气体。在另一个更优选的方面,所述气体是甲烷。在另一个更优选的方面,所述气体是H2。在另一个更优选的方面,所述气体是CO2。在另一个更优选的方面,所述气体是CO。参见,例如,Kataoka,N.,A.Miya,和K.Kiriyama,1997,Studies on hydrogen production bycontinuous culture system of hydrogen-producing anaerobic bacteria,WaterScience and Technology 36(6-7):41-47;和Gunaseelan V.N.于Biomass andBioenergy,Vol.13(1-2),pp.83-114,1997,Anaerobic digestion of biomass formethane production:A review。
回收可以使用本领域已知的任何方法,任选地从发酵培养基回收发酵产物,所述方法包括,但不限于,层析、电泳方法、差示溶解度、蒸馏或提取。例如,通过常规蒸馏方法从发酵的纤维素材料分离并纯化醇。可以获得纯度高达约96vol%的乙醇,其能用作,例如,燃料乙醇、饮用乙醇,即,中性饮料酒,或工业乙醇。
信号肽
本发明还涉及编码信号肽的分离的多核苷酸,所述信号肽包含SEQ IDNO:2的氨基酸1至19,或由SEQ ID NO:2的氨基酸1至19组成。
本发明还涉及包含编码蛋白的基因的核酸构建体,其中所述基因可操作地连接于编码信号肽的分离的多核苷酸和编码前多肽的分离的多核苷酸之一或二者,其中所述基因对编码所述信号肽和前肽的多核苷酸是外源的。
在一个优选的方面,所述编码信号肽的分离的多核苷酸包含SEQ ID NO:1的核苷酸1至57,或者由SEQ ID NO:1的核苷酸1至57组成。
本发明还涉及包含这种核酸构建体的重组表达载体和重组宿主细胞。
本发明还涉及用于产生蛋白质的方法,包括:(a)在有助于产生所述蛋白质的条件下培养这样的重组宿主细胞,所述宿主细胞包含编码蛋白的基因,所述基因可操作地连接于此种编码信号肽的多核苷酸,其中所述基因对编码所述信号肽的多核苷酸是外源的;和(b)回收所述蛋白质。
所述蛋白质对于宿主细胞可以是天然的或异源的。术语“蛋白质”在本文的意思不是指特定长度的编码产物,并且因此包含肽、寡肽和蛋白质。术语“蛋白质”还包含组合以形成编码产物的两种以上多肽。所述蛋白质还包括杂合多肽,其包含部分或全部多肽序列的组合,所述多肽序列从至少两种不同的蛋白质获得,其中一个或多个(几个)对于宿主细胞可以是异源或天然的。蛋白质进一步包括上述蛋白质和杂合蛋白质的天然存在的等位基因变异和工程改造的变异。
优选蛋白质是激素或其变体、酶、受体或其部分、抗体或其部分,或报告蛋白(reporter)。在一个更优选的方面,所述蛋白质是氧化还原酶、转移酶、水解酶、裂合酶(lyase)、异构酶或连接酶。在一个更加优选的方面,所述蛋白质是氨肽酶、淀粉酶、糖酶、羧肽酶、过氧化氢酶、纤维素酶、几丁质酶、角质酶、环糊精糖基转移酶、脱氧核糖核酸酶、酯酶、α-半乳糖苷酶、β-半乳糖苷酶、葡糖淀粉酶、α-葡糖苷酶、β-葡糖苷酶、转化酶、漆酶、另外的脂肪酶、甘露糖苷酶、变聚糖酶(mutanase)、氧化酶、果胶分解酶、过氧化物酶、肌醇六磷酸酶、多酚氧化酶、蛋白水解酶、核糖核酸酶、转谷氨酰胺酶或木聚糖酶。
基因可以从任何原核、真核或其它来源获得。
通过以下实施例进一步对本发明进行描述,但不应将其理解为对本发明范围的限制。
实施例
作为缓冲液和底物使用的化学品是至少试剂等级的商业产品。
材料
作为缓冲液和底物使用的化学品是至少试剂等级的商业产品。
菌株
将Trichophaea saccata菌株CBS 804.70用作编码家族GH3Aβ-葡糖苷酶的基因的来源。将米曲霉Jal250菌株(WO 99/61651)用于表达Trichophaeasaccata CBS 804.70β-葡糖苷酶。
培养基
MEX-1培养基由20g大豆粉、15g的麦麸(wheat bran)、10g的微晶纤维素( FMC,Philadelphia,PA,USA),5g的麦芽糊精,3g的细菌蛋白胨,0.2g的Pluronic,1g的橄榄油和去离子水加至1升组成。
FMC,Philadelphia,PA,USA),5g的麦芽糊精,3g的细菌蛋白胨,0.2g的Pluronic,1g的橄榄油和去离子水加至1升组成。
PDA平板由39克的马铃薯右旋糖琼脂和去离子水加至1升组成。
LB培养基由10g的胰蛋白胨、5g的酵母提取物、5g的氯化钠和去离子水加至1升组成。
LB平板由LB培养基和每升15g的细菌琼脂组成。
SOC培养基由2%胰蛋白胨、0.5%酵母提取物、10mM NaCl、2.5mM KCl、10mM MgCl2和10mM MgSO4和去离子水加至1升组成;通过高压灭菌来除菌,然后添加经过滤灭菌的葡萄糖至20mM。
MDU2BP培养基由45g的麦芽糖、1g的MgSO4·7H2O、1g的NaCl、2g的K2HSO4、12g的KH2PO4、2g的尿素、500μl的AMG微量金属溶液、和去离子水加至1升组成。将pH调整至5.0,然后使用0.22μm过滤单元进行过滤灭菌。
AMG微量金属溶液由14.3g的ZnSO4·7H2O、2.5g的CuSO4·5H2O、0.5g的NiCl2·6H2O、13.8g的FeSO4·7H2O、8.5g的MnSO4·7H2O、3g的柠檬酸和去离子水加至1升组成。
2X YT平板由每升的16g的胰蛋白胨、10g的酵母提取物、5g的NaCl、15g的细菌琼脂和去离子水加至1升组成。
YPG培养基由4g的酵母提取物、1g的K2HPO4、0.5g的MgSO4、15.0g的葡萄糖和去离子水加至1升(pH 6.0)组成。
YPM培养基由10g的酵母提取物、20g的细菌蛋白胨、20g的麦芽糖和去离子水加至1升组成。
M410培养基由50g的葡萄糖、50g的麦芽糖、2g的MgSO4·7H2O、2g的KH2PO4、4g的柠檬酸、8g的酵母提取物、2g的尿素、0.5g的CaCl2、0.5ml的AMG微量金属溶液和去离子水加至1升(pH 6.0)组成。
实施例1:制备Trichophaea saccata菌株CBS 804.70菌丝体用于cDNA文库产生
将Trichophaea saccata菌株CBS 804.70接种至PDA平板上,并在28℃温育7日。将几个菌丝体-PDA琼脂栓接种入含有100ml MEX-1培养基的750ml摇瓶。将烧瓶在37℃以150rpm搅拌温育9日。通过经由 (Calbiochem,San Diego,CA,USA)过滤来收获真菌菌丝体,然后将其冻结于液氮中。然后通过将冻结的菌丝体与相同体积的干冰在经液氮预冷却的咖啡磨(coffee grinder)中一同研磨来将菌丝体粉碎为粉末。将粉碎的菌丝体物质维持在-80℃直至使用。
(Calbiochem,San Diego,CA,USA)过滤来收获真菌菌丝体,然后将其冻结于液氮中。然后通过将冻结的菌丝体与相同体积的干冰在经液氮预冷却的咖啡磨(coffee grinder)中一同研磨来将菌丝体粉碎为粉末。将粉碎的菌丝体物质维持在-80℃直至使用。
实施例2:Trichophaea saccata菌株CBS 804.70RNA分离
根据Chirgwin等,1979,Biochemistry 18:5294-5299从冻结的、粉末状Trichophaea saccata菌丝体通过用硫氰酸胍提取然后通过5.7M CsCl垫(cushion)超离心来制备总RNA。根据Aviv等,1972,Proc.Natl.Acad.Sci.USA69:1408-1412通过寡聚(dT)-纤维素亲和层析来分离富含polyA的RNA。
实施例3:Trichophaea saccata菌株CBS 804.70cDNA文库的构建
双链cDNA是根据Gubler和Hoffman,1983,Gene 25:263-269;Sambrook,J.,Fritsch,E.F.和Maniantis,T.MolecularCloning:A Laboratory Manual,第2版,1989,Cold Spring Harbor Laboratory,Cold Spring Harbor,New York;以及Kofod等,1994,J.Biol.Chem.269:29182-29189的一般方法,使用多聚A-Not I引物(Promega Corp.,Madison,Wisconsin,USA)合成的。在合成之后,cDNA用绿豆核酸酶处理,用T4DNA聚合酶平端化,并连接于50倍摩尔过量的Eco RI衔接头(Invitrogen Corp.,Carlsbad,CA,USA)。将cDNA用Not I切割,并将cDNA通过0.8%琼脂糖凝胶电泳使用44mM Tris碱、44mM硼酸和0.5mM EDTA(TBE)缓冲液进行大小分级。将700bp和更大的cDNA级分从凝胶切出,并使用 PCR DNA和Gel Band Purification Kit (GE Healthcare LifeSciences,United Kingdom)依照生产商的指示进行纯化。
PCR DNA和Gel Band Purification Kit (GE Healthcare LifeSciences,United Kingdom)依照生产商的指示进行纯化。
然后对制备的cDNA进行定向克隆,即使用Rapid Ligation Kit(RocheDiagnostics GmbH,Penzberg,Germany)依照生产商的指示将其连接入经EcoRI-Not I切割的pMHas5(WO 03/044049)。将连接混合物使用 和Pulse Controller(Bio-Rad Laboratories,Inc.,Hercules,CA,USA)以50μF,25mAmp,1.8kV和2mm间隙(gap)小杯依照生产商的指示电穿孔入大肠杆菌DH10B细胞(Invitrogen Corp.,Carlsbad,CA,USA)。
和Pulse Controller(Bio-Rad Laboratories,Inc.,Hercules,CA,USA)以50μF,25mAmp,1.8kV和2mm间隙(gap)小杯依照生产商的指示电穿孔入大肠杆菌DH10B细胞(Invitrogen Corp.,Carlsbad,CA,USA)。
将经电穿孔的细胞铺板于补充每ml 50μg卡那霉素的LB平板。从起始cDNA-pMHas5载体连接液的总共大约30000个转化体制备cDNA质粒汇集库(pool)。直接从菌落的汇集库使用 Spin Midi/Maxiprep Kit(QIAGEN GmbH Corporation,Hilden,Germany)制备质粒DNA。cDNA文库命名为SBL521-2。
Spin Midi/Maxiprep Kit(QIAGEN GmbH Corporation,Hilden,Germany)制备质粒DNA。cDNA文库命名为SBL521-2。
实施例4:构建含有β-内酰胺酶报道基因的SigA4转座子
从含有WO 01/77315中描述的质粒的pSigA2转座子构建含有命名为pSigA4的质粒的转座子,以创建具有减少的选择背景的信号俘获转座子(signal trapping transposon)pSigA2的改善形式。所述pSigA2转座子含有在该转座子自身上编码的无信号的β-内酰胺酶构建体。使用校正读码的 DNA聚合酶(QIAGEN GmbH Corporation,Hilden,Germany)和下述5’磷酸化的引物(TAG Copenhagen,Denmark)采用PCR以产生存在于质粒骨架上的完整β-内酰胺酶基因的缺失:
DNA聚合酶(QIAGEN GmbH Corporation,Hilden,Germany)和下述5’磷酸化的引物(TAG Copenhagen,Denmark)采用PCR以产生存在于质粒骨架上的完整β-内酰胺酶基因的缺失:
SigA2NotU-P:
5’-TCGCGATCCGTTTTCGCATTTATCGTGAAACGCT-3’(SEQ ID NO:71)
SigA2NotD-P:
5’-CCGCAAACGCTGGTGAAAGTAAAAGATGCTGAA-3’(SEQ ID NO:72)
所述扩增反应物由1μl的pSigA2(10ng/μl)、5μl的10X PROOFSTARTTMBuffer(QIAGEN GmbH Corporation,Hilden,Germany)、2.5μl的dNTP混合物(20mM)、0.5μl的SigA2NotU-P(10mM)、0.5μl的SigA2NotD-P(10mM)、10μl的Q溶液(QIAGEN GmbH Corporation,Hilden,Germany)和31.25μl的去离子水。使用DNA ENGINETM Thermal Cycler(MJ Research Inc.,Waltham,MA,USA)进行扩增,其程序为1循环,在95℃进行5分钟;和20个循环,每循环在94℃进行30秒,62℃进行30秒和72℃进行4分钟。
通过0.8%琼脂糖凝胶电泳使用40mM Tris碱-20mM乙酸钠-1mM EDTA二钠(TAE)缓冲液和每升0.1μg的溴乙锭分离3.9kb PCR反应产物。DNA条带用Eagle Eye Imaging System(Stratagene,La Jolla,CA,USA)协助在360nm显现。将3.9kb DNA条带从凝胶切出,并使用 PCR DNA和Gel BandPurification Kit依照生产商的指示进行纯化。
PCR DNA和Gel BandPurification Kit依照生产商的指示进行纯化。
将该3.9kb片段在16℃用10单位T4DNA连接酶(New England Biolabs,Inc.,Beverly,MA,USA)、9μl的该3.9kb PCR片段和1μl的10X连接缓冲液(New England Biolabs,Inc.,Beverly,MA,USA)自连接过夜。将连接物在65℃热失活10分钟,然后用DpnI在37℃消化2小时。在温育之后,使用 PCR DNA和Gel Band Purification Kit纯化消化液。
PCR DNA和Gel Band Purification Kit纯化消化液。
然后将纯化的物质依照生产商的指示转化入大肠杆菌TOP10感受态细胞(Invitrogen Corp.,Carlsbad,CA,USA)。将转化混合物铺板于补充每ml 25μg的氯霉素的LB平板之上。从几个转化体制备质粒小量制备物,并用Bgl II消化。选择一个具有正确构建的质粒并命名为pSigA4。质粒pSigA4含有与WO 01/77315中公开的相同的侧翼为Bgl II的转座子SigA2(SEQ ID NO:71)。
将质粒pSigA4DNA(0.3μg/μl)的60μl样品用Bgl II消化,并使用TAE缓冲液通过0.8%琼脂糖凝胶电泳进行分离。将2kb的SigA2转座子DNA条带用200μl的EB缓冲液(QIAGEN GmbH Corporation,Hilden,Germany)洗脱,并使用 PCR DNA和Gel Band Purification Kit依照生产商的指示进行纯化,并在200μl的EB缓冲液中洗脱。将SigA2用于转座子协助的信号俘获。
PCR DNA和Gel Band Purification Kit依照生产商的指示进行纯化,并在200μl的EB缓冲液中洗脱。将SigA2用于转座子协助的信号俘获。
实施例5:Trichophaea saccata CBS 804.70的转座子协助的信号俘获
对于转座子协助的信号俘获的完整描述可见于WO 2001/77315。从起始cDNA-pMHas5载体连接液的总共30000个转化体制备cDNA质粒汇集库。质粒DNA是从由固体LB选择性培养基回收的菌落汇集物使用 Spin Midi/Maxiprep Kit直接制备的。用转座子SigA2和MuA转座酶(Finnzymes OY,Espoo,Finland)依照生产商的指示处理所述质粒汇集库。
Spin Midi/Maxiprep Kit直接制备的。用转座子SigA2和MuA转座酶(Finnzymes OY,Espoo,Finland)依照生产商的指示处理所述质粒汇集库。
对于Trichophaea saccata CBS 804.70cDNA文库的体外转座子标记,将含有大约2.6μg的DNA的4或8μl SigA2转座子与含有2μg的DNA,2μl的MuA转座酶(0.22μg/μl)和5μl的5X缓冲液(Finnzymes OY,Espoo,Finland)的Trichophaea saccata CBS 804.70cDNA文库的1μl质粒DNA汇集库在总体积50μl中混合,并在30℃温育3.5小时,然后在75℃热失活10分钟。将DNA通过添加5μl的3M乙酸钠pH 5和110μl的96%乙醇沉淀,并以10000x g离心30分钟。将沉淀在70%乙醇中洗涤,并在室温干燥,并重悬于10μl的10mM Tris,pH 8,1mMEDTA(TE)缓冲液。
将1.5μl体积的转座子标记质粒汇集库依照生产商的指示使用GENE 和Pulse Controller(Bio-Rad Laboratories,Inc.,Hercules,CA,USA)以50μF,25mAmp,1.8kV用2mm间隔小杯电穿孔入20μl的大肠杆菌DH10B超感受态(ultracompetent)细胞(Gibco-BRL,Gaithersburg MD,USA)。
和Pulse Controller(Bio-Rad Laboratories,Inc.,Hercules,CA,USA)以50μF,25mAmp,1.8kV用2mm间隔小杯电穿孔入20μl的大肠杆菌DH10B超感受态(ultracompetent)细胞(Gibco-BRL,Gaithersburg MD,USA)。
将经电穿孔的细胞在28℃在SOC培养基中以250rpm搅拌温育2小时,然后铺板于下述选择培养基:补充了50μg每ml卡那霉素的LB培养基;补充了50μg每ml卡那霉素和15μg每ml氯霉素的LB培养基;和/或补充了50μg每ml卡那霉素、15μg每ml氯霉素和12.5μg每ml氨苄青霉素的LB培养基。
从将电穿孔液稀释铺板于补充了卡那霉素和氯霉素培养基的LB培养基,确定了每个电穿孔液存在大约72000个含有具SigA2转座子的cDNA质粒文库的菌落,并在三重选择(LB,卡那霉素,氯霉素,氨苄青霉素)下回收了大约69个菌落。进行进一步的电穿孔和铺板实验直至从该三重选择下回收了总共445个菌落。使用 96Turbo Miniprep Kit(QIAGEN GmbH Corporation,Hilden,Germany)小量制备所述菌落。用下述所示的转座子正向和反向引物(引物A和B)依照WO 2001/77315(28页)公开的步骤对质粒进行测序。
96Turbo Miniprep Kit(QIAGEN GmbH Corporation,Hilden,Germany)小量制备所述菌落。用下述所示的转座子正向和反向引物(引物A和B)依照WO 2001/77315(28页)公开的步骤对质粒进行测序。
引物A:
5’-AGCGTTTGCGGCCGCGATCC-3’(SEQ ID NO:73)
引物B:
5’-TTATTCGGTCGAAAAGGATCC-3’(SEQ ID NO:74)
实施例6:序列汇编(assembly)和注释(annotataion)
获取DNA序列以供使用ABI PRISM Automated DNA Sequencer Model3700(Applied Biosystems,Foster City,CA,USA)的反应。修整对于每个质粒读取的(reads for each plasmid)引物A和引物B序列以去除载体和转座子序列。这产生了225个经汇编的序列,通过使用程序PhredPhrap(Ewing等,1998,Genome Research 8:175-185;Ewing和Green,1998,Genome Research 8:186-194)将其分组为148个重叠群(contig)。随后将所有148个重叠群与在标准的公共DNA和蛋白序列数据库(TrEMBL,SWALL,PDB,EnsemblPep,GeneSeqP)可获得的序列通过使用程序BLASTX 2.0a19MP-WashU[14-Jul-1998][Build linux-x8618:51:4430-Jul-1998](Gish等,1993,Nat.Genet.3:266-72)进行比较。通过分析BlastX结果直接鉴定了家族GH3Aβ-葡糖苷酶。
从随机位于沿编码区的15个独立转座事件的汇编提取所述DNA重叠群的开读框。Trichophaea saccata β-葡糖苷酶的核苷酸序列(SEQ ID NO:1)和推定的氨基酸序列(SEQ ID NO:2)示于图1A和1B。所述cDNA片段编码856个氨基酸的多肽。所述基因的%G+C含量为58.1%。使用SignalP软件程序(Nielsen等,1997,Protein Engineering 10:1-6),预测了19个残基的单个肽。该预测的成熟蛋白包含具有90.35kDa的分子量的837个氨基酸。
β-葡糖苷酶家族3序列的比较性比对是使用如EMBOSS的Needle程序中执行的Needleman-Wunsch算法(Needleman和Wunsch,1970,J.Mol.Biol.48:443-453)以缺口开放罚分为10,缺口延伸罚分为0.5和EBLOSUM62矩阵来确定的。该比对显示Trichophaea saccata Cel3A成熟多肽的推定的氨基酸序列与来自核盘菌(Sclerotinia sclerotiorum)的假设预测的蛋白的推定氨基酸序列(Uniprot accession number A7EI88)共享59.3%的同一性(排除缺口)。
实施例7:将Trichophaea saccata家族GH3A基因克隆入米曲霉表达载体
设计了如下所示的两个合成寡核苷酸引物以从cDNA PCR扩增Trichophaea saccata gh3a基因。使用IN-FUSIONTM PCR Cloning Kit(ClontechLaboratories,Inc.,Mountain View,CA,USA)将片段直接克隆入表达载体pAILo2(WO 2005/074647)。
正向引物:
5’-ACTGGATTTACC -3’(SEQ ID NO:75)
-3’(SEQ ID NO:75)
反向引物:
5’-TCACCTCTAGTTAATTA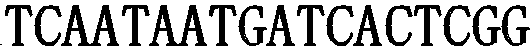 -3’(SEQ ID NO:76)
-3’(SEQ ID NO:76)
粗体字母代表编码序列。剩余序列与pAlLo2的插入位点同源。
将五十皮摩尔的每种上述引物用于含有1μl(239μg)的SBL521-2cDNA文库,含MgCl2的1X High Fidelity缓冲液(Roche Diagnostics Corp.,Indianapolis,IN,USA),1μl的10mM dATP、dTTP、dGTP和dCTP的混合物和2.6单位的
High Fidelity缓冲液(Roche Diagnostics Corp.,Indianapolis,IN,USA),1μl的10mM dATP、dTTP、dGTP和dCTP的混合物和2.6单位的 High Fidelity酶混合物(Roche,Indianapolis,IN,USA),最终体积为50μl的PCR反应。使用
High Fidelity酶混合物(Roche,Indianapolis,IN,USA),最终体积为50μl的PCR反应。使用  epgradient S(Eppendorf Scientific,Inc.,Westbury,NY,USA)扩增片段,程序为一个循环94℃进行2分钟,10个循环,每个循环在94℃进行15秒,58℃进行30秒和72℃进行2分钟;20个循环,每个在94℃进行15秒,58℃进行30秒和72℃进行120秒(每个后续循环加5秒)。然后将加热块维持在72℃进行7分钟,然后进行4℃浸泡循环。
epgradient S(Eppendorf Scientific,Inc.,Westbury,NY,USA)扩增片段,程序为一个循环94℃进行2分钟,10个循环,每个循环在94℃进行15秒,58℃进行30秒和72℃进行2分钟;20个循环,每个在94℃进行15秒,58℃进行30秒和72℃进行120秒(每个后续循环加5秒)。然后将加热块维持在72℃进行7分钟,然后进行4℃浸泡循环。
将反应产物通过使用TAE缓冲液的1.0%琼脂糖凝胶电泳分离,并将大约2.5kb产物条带从凝胶切出,并使用 Gel Extraction Kit(QIAGENInc.,Valencia,CA,USA)依照生产商的指示进行纯化。
Gel Extraction Kit(QIAGENInc.,Valencia,CA,USA)依照生产商的指示进行纯化。
然后将纯化的PCR片段使用IN-FUSIONTM PCR Cloning Kit克隆入pAlLo2。用Nco I和Pac I消化载体。将经消化的片段通过如上所述的凝胶电泳纯化,从凝胶切出,并使用 Gel Extraction Kit(QIAGEN Inc.,Valencia,CA,USA)纯化。将基因片段和经消化的载体在反应中合并,得到表达质粒pAG39,其中gh3a基因的转录是处于NA2-tpi启动子(来自黑曲霉中性α-淀粉酶和构巢曲霉丙糖磷酸异构酶的基因的启动子的杂合体)的控制下。重组反应物(20μl)是由1X IN-FUSIONTM Buffer(Clontech,Mountain View,CA,USA),1X B SA(Clontech,Mountain View,CA,USA),1μl的IN-FUSIONTM酶(1∶10稀释)(Clontech,Mountain View,CA,USA),249ng的经Nco I和Pac I消化的pAlLo2和82.5ng的纯化的Trichophaea saccata gh3a PCR产物组成的。将所述反应物在37℃温育15分钟,然后在50℃温育15分钟。将反应物用40μl的TE缓冲液稀释,并将2.5μl的经稀释的反应物用于转化大肠杆菌TOP 10感受态细胞(Invitrogen,Carlsbad,CA,USA)。
Gel Extraction Kit(QIAGEN Inc.,Valencia,CA,USA)纯化。将基因片段和经消化的载体在反应中合并,得到表达质粒pAG39,其中gh3a基因的转录是处于NA2-tpi启动子(来自黑曲霉中性α-淀粉酶和构巢曲霉丙糖磷酸异构酶的基因的启动子的杂合体)的控制下。重组反应物(20μl)是由1X IN-FUSIONTM Buffer(Clontech,Mountain View,CA,USA),1X B SA(Clontech,Mountain View,CA,USA),1μl的IN-FUSIONTM酶(1∶10稀释)(Clontech,Mountain View,CA,USA),249ng的经Nco I和Pac I消化的pAlLo2和82.5ng的纯化的Trichophaea saccata gh3a PCR产物组成的。将所述反应物在37℃温育15分钟,然后在50℃温育15分钟。将反应物用40μl的TE缓冲液稀释,并将2.5μl的经稀释的反应物用于转化大肠杆菌TOP 10感受态细胞(Invitrogen,Carlsbad,CA,USA)。
使用 9600(QIAGEN Inc.,Valencia,CA,USA)制备质粒DNA,并用Bam HI消化推定的pAG39克隆。然后对来自这些克隆的质粒DNA进行测序以鉴定不含PCR诱导的错误的克隆。测序反应物含有1.5μl的质粒DNA,4.5μl的水和4μl的测序主混合物(sequencing master-mix),所述主混合物含有1μl的5X测序缓冲液(Millipore,Billerica,MA,USA),1μl的BIGDYETM终止物(terminator)(Applied Biosystems,Inc.,Foster City,CA,USA),1μl的水和每反应3.2皮摩尔的下述引物之一。
9600(QIAGEN Inc.,Valencia,CA,USA)制备质粒DNA,并用Bam HI消化推定的pAG39克隆。然后对来自这些克隆的质粒DNA进行测序以鉴定不含PCR诱导的错误的克隆。测序反应物含有1.5μl的质粒DNA,4.5μl的水和4μl的测序主混合物(sequencing master-mix),所述主混合物含有1μl的5X测序缓冲液(Millipore,Billerica,MA,USA),1μl的BIGDYETM终止物(terminator)(Applied Biosystems,Inc.,Foster City,CA,USA),1μl的水和每反应3.2皮摩尔的下述引物之一。
引物pAlLo25’:
5’-TGTCCCTTGTCGATGCG-3’(SEQ ID NO:77)
引物pAlLo23’:
5’-CACATGACTTGGCTTCC-3’(SEQ ID NO:78)
引物Ts3AF1:
5’-GTCTACCTCTGGCCATTCCA-3’(SEQ ID NO:79)
引物Ts3AF2:
5’-TACCGCAATCCTCCAGAACA-3’(SEQ ID NO:80)
引物Tsac gh3a 1675-17000Rev:
5’-ACGCTTGTCGGCCCATTTCTCCATGA-3’(SEQ ID NO:81)
引物Tsac gh3a 925-950Rev:
5’-TGGGCCCAGCAAGGGCACACGATCAT-3’(SEQ ID NO:82)
实施例8:在米曲霉JaL355中表达编码具有β-葡糖苷酶活性的GH3A多肽的Trichophaea saccata cDNA
米曲霉JaL355原生质体是根据Christensen等,1988,Bio/Technology 6:1419-1422的方法制备的。将五μg的pAG39用于转化米曲霉JaL355。将二十四个转化体分离至单个PDA平板。
将24个转化体的汇合的PDA平板用5ml的0.01% 20洗涤,并收集孢子。将八μl的每个孢子储备分别添加至24孔板中1ml的YPG、YPM和M410培养基,并在34℃温育。在温育三日之后,使用
20洗涤,并收集孢子。将八μl的每个孢子储备分别添加至24孔板中1ml的YPG、YPM和M410培养基,并在34℃温育。在温育三日之后,使用 无染色,8-16%梯度SDS-PAGE凝胶(BioRad Laboratories,Inc.,Hercules,CA,USA)依照生产商的指示分析7.5μl来自培养的上清。培养的SDS-PAGE分布显示几个转化体具有大约100kDa的新的主要条带,并在M410培养基中具有最佳表达。在总共五日的温育之后,对所有M410培养进行取样,并如上所述通过SDS-PAGE进行分析,此时选择显示最佳表达的转化体。
无染色,8-16%梯度SDS-PAGE凝胶(BioRad Laboratories,Inc.,Hercules,CA,USA)依照生产商的指示分析7.5μl来自培养的上清。培养的SDS-PAGE分布显示几个转化体具有大约100kDa的新的主要条带,并在M410培养基中具有最佳表达。在总共五日的温育之后,对所有M410培养进行取样,并如上所述通过SDS-PAGE进行分析,此时选择显示最佳表达的转化体。
用5ml的0.01% 20洗涤最佳转化体的汇集PDA平板,并接种入五个含有100ml M410培养基的500ml摇瓶以生成用于表征该酶的培养液。在第5日收获烧瓶。使用0.22μm stericup吸滤器(Millipore,Bedford,MA,USA)过滤培养液。
20洗涤最佳转化体的汇集PDA平板,并接种入五个含有100ml M410培养基的500ml摇瓶以生成用于表征该酶的培养液。在第5日收获烧瓶。使用0.22μm stericup吸滤器(Millipore,Bedford,MA,USA)过滤培养液。
实施例9:表征Trichophaea saccata GH3β-葡糖苷酶
对纤维二糖和4-硝基苯基-β-D-葡萄糖吡喃糖苷:将纤维二糖溶解于50mM乙酸钠-0.01% 20pH 5缓冲液至2g/L。使用相同缓冲液将酶以多种浓度稀释。向50μl的酶稀释物(0.04-5μg/ml)添加100μl的纤维二糖溶液(2g/L缓冲液)。将反应在50℃温育30分钟,接着用50μl的0.5M氢氧化钠停止反应。使用Glucose Oxidase试剂(Pointe Scientific,Inc.,Canton,MI,USA)依照生产商的指示测量产生的葡萄糖。
20pH 5缓冲液至2g/L。使用相同缓冲液将酶以多种浓度稀释。向50μl的酶稀释物(0.04-5μg/ml)添加100μl的纤维二糖溶液(2g/L缓冲液)。将反应在50℃温育30分钟,接着用50μl的0.5M氢氧化钠停止反应。使用Glucose Oxidase试剂(Pointe Scientific,Inc.,Canton,MI,USA)依照生产商的指示测量产生的葡萄糖。
将4-硝基苯基-β-D-葡萄糖吡喃糖苷溶解于DMSO至100mM。在即将进行测定前,将100mM 4-硝基苯基-β-D-葡萄糖吡喃糖苷储液在50mM乙酸钠-0.01% 20缓冲液中稀释100X(1mM 4-硝基苯基-β-D-葡萄糖吡喃糖苷)。使用相同缓冲液将酶以多种浓度稀释。向20μl的每个酶稀释物,添加100μl的4-硝基苯基-β-D-葡萄糖吡喃糖苷溶液。将反应在40℃温育20分钟,然后用50μl的1M碳酸钠pH 10缓冲液停止反应。在405nm处通过分光光度法确定释放的4-硝基苯酚。
20缓冲液中稀释100X(1mM 4-硝基苯基-β-D-葡萄糖吡喃糖苷)。使用相同缓冲液将酶以多种浓度稀释。向20μl的每个酶稀释物,添加100μl的4-硝基苯基-β-D-葡萄糖吡喃糖苷溶液。将反应在40℃温育20分钟,然后用50μl的1M碳酸钠pH 10缓冲液停止反应。在405nm处通过分光光度法确定释放的4-硝基苯酚。
结果说明Trichophaea saccata GH3β-葡糖苷酶对两种底物均具有活性。
热稳定性:将Trichophaea saccata GH3A β-葡糖苷酶在50mM乙酸钠-0.01% 20pH 5缓冲液中稀释至1g/L,然后在50℃温育72小时,和在60℃温育3小时或24小时。将相同的样品储藏于4℃充当对照。在温育之后,使用4-硝基苯基-β-D-葡萄糖吡喃糖苷作为底物根据上述测定法,但使用在该测定法中给出少于5%转化的一种酶加载量来测量样品的残余活性。在4℃的样品的活性标准化为100%,而将在其他温育条件的样品的活性与4℃活性相比较。Trichophaea saccata GH3A β-葡糖苷酶的热稳定性示于下述表I。
20pH 5缓冲液中稀释至1g/L,然后在50℃温育72小时,和在60℃温育3小时或24小时。将相同的样品储藏于4℃充当对照。在温育之后,使用4-硝基苯基-β-D-葡萄糖吡喃糖苷作为底物根据上述测定法,但使用在该测定法中给出少于5%转化的一种酶加载量来测量样品的残余活性。在4℃的样品的活性标准化为100%,而将在其他温育条件的样品的活性与4℃活性相比较。Trichophaea saccata GH3A β-葡糖苷酶的热稳定性示于下述表I。
表I:Trichophaea saccata GH3A β-葡糖苷酶的热稳定性
| 温育条件 | 残余活性 |
| 4℃ | 100% |
| 50℃,72小时 | 95% |
| 60℃,3小时 | 23% |
| 60℃,24小时 | 6% |
pH分布:Trichophaea saccata GH3A β-葡糖苷酶的pH活性分布是使用上述用于确定活性的相同实验方案确定的,只是酶是在五个不同pH温育的:pH4、5、6、7、8,以及使用在该测定中给出少于5%底物转化的一种酶加载量。将Britton-Robinson缓冲液用作缓冲液系统。为了制备所述Britton-Robinson缓冲液,制备由在1升去离子水中0.1摩尔硼酸,0.1摩尔乙酸和0.1摩尔磷酸组成的100mM储液。然后将100mM储液使用5M氢氧化钠滴定至一定pH(4~8),然后稀释至40mM。用该缓冲液制备4-硝基苯基-β-D-葡萄糖吡喃糖苷,并在50℃测量其活性。将最高的活性标准化为100%,并将在其他pH值处的活性与该最高活性相比较并表示为%活性。Trichophaea saccata GH3β-葡糖苷酶的pH活性分布示于表II。
表II:pH活性分布
| pH值 | 活性 |
| 4.0 | 100% |
| 5.0 | 94% |
| 6.0 | 64% |
| 7.0 | 26% |
| 8.0 | 1% |
生物材料的保藏
下述的生物材料已经依据布达佩斯条约的条款保藏于农业研究机构专利培养物保藏中心/农业研究培养物保藏中心(Agricultural Research ServicePatent Culture Collection),北区研究中心(Northern Regional Research Center),1815University Street,Peoria,IL,USA,并给予下述的登录号:
保藏物 登录号 保藏日期
大肠杆菌pAHYG-33 NRRL B-50214 2008年12月22日
所述菌株于下述条件下保藏:确保在本专利申请未决期间,依据该外国专利法律的授权的人能够获得所述培养物。所述保藏物为所保藏菌株的基本上纯的培养物。在提交了该申请的副本,或其后续文本的国家,依据该外国专利法律可以获得所述保藏物。然而,应当理解,保藏物的获得并不构成对实施本发明的许可,实施本发明是对政府行为所授予的专利权的侵犯。
通过下述编号段落进一步描述本发明:
[1]具有β-葡糖苷酶活性的分离的多肽,其选自下组:
(a)包含氨基酸序列的多肽,所述氨基酸序列与SEQ ID NO:2的成熟多肽具有至少60%序列同一性;
(b)由多核苷酸编码的多肽,所述多核苷酸在至少中严格条件下与以下杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链;
(c)由多核苷酸编码的多肽,所述多核苷酸包含与SEQ ID NO:1的成熟多肽编码序列具有至少60%序列同一性的核苷酸序列;和
(d)SEQ ID NO:2的成熟多肽的包含取代、缺失和/或插入一个或多个(几个)氨基酸的变体。
[2]段1的多肽,其包含氨基酸序列,所述氨基酸序列与SEQ ID NO:2的氨基酸序列具有至少60%序列同一性。
[3]段2的多肽,其包含氨基酸序列,所述氨基酸序列与SEQ ID NO:2的氨基酸序列具有至少65%序列同一性。
[4]段3的多肽,其包含氨基酸序列,所述氨基酸序列与SEQ ID NO:2的氨基酸序列具有至少70%序列同一性。
[5]段4的多肽,其包含氨基酸序列,所述氨基酸序列与SEQ ID NO:2的氨基酸序列具有至少75%序列同一性。
[6]段5的多肽,其包含氨基酸序列,所述氨基酸序列与SEQ ID NO:2的氨基酸序列具有至少80%序列同一性。
[7]段6的多肽,其包含氨基酸序列,所述氨基酸序列与SEQ ID NO:2的氨基酸序列具有至少85%序列同一性。
[8]段7的多肽,其包含氨基酸序列,所述氨基酸序列与SEQ ID NO:2的氨基酸序列具有至少90%序列同一性。
[9]段8的多肽,其包含氨基酸序列,所述氨基酸序列与SEQ ID NO:2的氨基酸序列具有至少95%序列同一性。
[10]段9的多肽,其包含氨基酸序列,所述氨基酸序列与SEQ ID NO:2的氨基酸序列具有至少97%序列同一性。
[11]段1的多肽,其包含SEQ ID NO:2的氨基酸序列或其具有β-葡糖苷酶活性的片段或由SEQ IDNO:2的氨基酸序列或其具有β-葡糖苷酶活性的片段组成。
[12]段11的多肽,其包含SEQ ID NO:2的氨基酸序列或由SEQ ID NO:2的氨基酸序列组成。
[13]段1的多肽,其包含SEQ ID NO:2的成熟多肽或由SEQ ID NO:2的成熟多肽组成。
[14]段1的多肽,其由多核苷酸编码,所述多核苷酸在至少中严格条件下与下述杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链。
[15]段14的多肽,其由多核苷酸编码,所述多核苷酸在至少中-高严格条件下与下述杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ IDNO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链。
[16]段15的多肽,其由多核苷酸编码,所述多核苷酸在至少高严格条件下与下述杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链。
[17]段16的多肽,其由多核苷酸编码,所述多核苷酸在至少非常高严格条件下与下述杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ IDNO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链。
[18]段1的多肽,其由包含核苷酸序列的多核苷酸编码,所述核苷酸序列与SEQ ID NO:1的成熟多肽编码序列具有至少60%序列同一性。
[19]段18的多肽,其由包含核苷酸序列的多核苷酸编码,所述核苷酸序列与SEQ ID NO:1的成熟多肽编码序列具有至少65%序列同一性。
[20]段19的多肽,其由包含核苷酸序列的多核苷酸编码,所述核苷酸序列与SEQ ID NO:1的成熟多肽编码序列具有至少70%序列同一性。
[21]段20的多肽,其由包含核苷酸序列的多核苷酸编码,所述核苷酸序列与SEQ ID NO:1的成熟多肽编码序列具有至少75%序列同一性。
[22]段21的多肽,其由包含核苷酸序列的多核苷酸编码,所述核苷酸序列与SEQ ID NO:1的成熟多肽编码序列具有至少80%序列同一性。
[23]段22的多肽,其由包含核苷酸序列的多核苷酸编码,所述核苷酸序列与SEQ ID NO:1的成熟多肽编码序列具有至少85%序列同一性。
[24]段23的多肽,其由包含核苷酸序列的多核苷酸编码,所述核苷酸序列与SEQ ID NO:1的成熟多肽编码序列具有至少90%序列同一性。
[25]段24的多肽,其由包含核苷酸序列的多核苷酸编码,所述核苷酸序列与SEQ ID NO:1的成熟多肽编码序列具有至少95%序列同一性。
[26]段25的多肽,其由包含核苷酸序列的多核苷酸编码,所述核苷酸序列与SEQ ID NO:1的成熟多肽编码序列具有至少97%序列同一性。
[27]段1的多肽,其由多核苷酸编码,所述多核苷酸包含SEQ ID NO:1的核苷酸序列或其编码具有β-葡糖苷酶活性的片段的亚序列或由SEQ ID NO:1的核苷酸序列或其编码具有β-葡糖苷酶活性的片段的亚序列组成。
[28]段27的多肽,其由多核苷酸编码,所述多核苷酸包含SEQ ID NO:1的核苷酸序列或由SEQ ID NO:1的核苷酸序列组成。
[29]段1的多肽,其由多核苷酸编码,所述多核苷酸包含SEQ ID NO:1的成熟多肽编码序列或由SEQ ID NO:1的成熟多肽编码序列组成。
[30]段1的多肽,其中所述多肽是SEQ ID NO:2的成熟多肽的包含一个或多个(几个)氨基酸的取代、缺失和/或插入的变体。
[31]段1的多肽,所述多肽由质粒pAHYG-33中含有的多核苷酸编码,所述质粒pAHYG-33包含在大肠杆菌NRRL B-50214中。
[32]段1-31中任一项的多肽,其中成熟多肽是SEQ ID NO:2的氨基酸20至856。
[33]段1-32中任一项的多肽,其中成熟多肽编码序列是SEQ ID NO:1的核苷酸58至2568。
[34]分离的多核苷酸,其包含编码段1-33中任一项的多肽的核苷酸序列。
[35]段34的分离的多核苷酸,其在SEQ ID NO:1的成熟多肽编码序列中包含至少一个突变,其中所述突变核苷酸序列编码SEQ ID NO:2的成熟多肽。
[36]核酸构建体,其包含段34或35的多核苷酸,所述多核苷酸与一个或多个(几个)调控序列可操作地连接,所述调控序列指导多肽在表达宿主中产生。
[37]重组表达载体,其包含段34或35的多核苷酸。
[38]重组宿主细胞,其包含段34或35的多核苷酸,所述多核苷酸与一个或多个(几个)调控序列可操作地连接,所述调控序列指导具有α-葡糖醛酸糖苷酶的产生。
[39]用于产生段1-33中任一项的多肽的方法,其包括:(a)在有助于所述多肽产生的条件下培养细胞,所述细胞以其野生型形式产生所述多肽;和(b)回收所述多肽。
[40]用于产生段1-33中任一项的多肽的方法,其包括:(a)在有助于所述多肽产生的条件下培养包含核酸构建体的宿主细胞,所述核酸构建体包含编码所述多肽的核苷酸序列;和(b)回收所述多肽。
[41]用于产生亲本细胞的突变体的方法,所述方法包括破坏或缺失编码段1-33中任一项的多肽或其一部分的多核苷酸,其导致突变体与亲本细胞相比产生较少的所述多肽。
[42]由段41的方法产生的突变细胞。
[43]段42的突变细胞,其进一步包含编码天然或异源蛋白的基因。
[44]产生蛋白的方法,其包括:(a)在有助于所述蛋白产生的条件下培养段43的突变细胞;和(b)回收所述蛋白。
[45]段34或35的分离的多核苷酸,其通过下述方法获得:(a)在至少中严格条件下将DNA群体与下述杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链;和(b)分离杂交的多核苷酸,其编码具有β-葡糖苷酶活性的多肽。
[46]段45的分离的多核苷酸,其通过下述方法获得:(a)在至少中-高严格条件下将DNA群体与下述杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链;和(b)分离杂交的多核苷酸,其编码具有β-葡糖苷酶活性的多肽。
[47]段46的分离的多核苷酸,其通过下述方法获得:(a)在至少高严格条件下将DNA群体与下述杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链;和(b)分离杂交的多核苷酸,其编码具有β-葡糖苷酶活性的多肽。
[48]段47的分离的多核苷酸,其通过下述方法获得:(a)在至少非常高严格条件下将DNA群体与下述杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链;和(b)分离杂交的多核苷酸,其编码具有β-葡糖苷酶活性的多肽。
[49]段45-48的分离的多核苷酸,其中成熟多肽编码序列是SEQ ID NO:1的核苷酸58至2568。
[50]用于产生包含突变核苷酸序列的多核苷酸的方法,所述突变核苷酸序列编码具有β-葡糖苷酶活性的多肽,所述方法包括:(a)向SEQ ID NO:1的成熟多肽编码序列中引入至少一个突变,其中突变核苷酸序列编码包含SEQ ID NO:2的成熟多肽或者由SEQ ID NO:2的成熟多肽组成的多肽;和(b)回收所述包含突变核苷酸序列的多核苷酸。
[51]由段50的方法产生的突变多核苷酸。
[52]产生多肽的方法,其包括:(a)在有助于所述多肽产生的条件下培养细胞,所述细胞包含段51的编码所述多肽的突变多核苷酸;和(b)回收所述多肽。
[53]产生段1-33中任一项的多肽的方法,其包括:(a)在有助于所述多肽产生的条件下培养转基因植物或植物细胞,所述转基因植物或植物细胞包含编码所述多肽的多核苷酸;和(b)回收所述多肽。
[54]转基因植物、植物部分或植物细胞,其用编码段1-33中任一项的多肽的多核苷酸转化。
[55]双链抑制RNA(dsRNA)分子,其包含段34或35的多核苷酸的亚序列,其中dsRNA任选地为siRNA或miRNA分子。
[56]段55的双链抑制RNA(dsRNA)分子,其为长约15、16、17、18、19、20、21、22、23、24、25或更长的双链体核苷酸。
[57]用于抑制具有β-葡糖苷酶活性的多肽在细胞中表达的方法,其包括对细胞施用双链RNA(dsRNA)分子,或者在细胞中表达双链RNA(dsRNA)分子,其中所述dsRNA包括段34或35的多核苷酸的亚序列。
[58]段57的方法,其中dsRNA为长约15、16、17、18、19、20、21、22、23、24、25或更长的双链体核苷酸。
[59]分离的多核苷酸,其编码信号肽,所述信号肽包含SEQ ID NO:2的氨基酸1至19或由SEQ ID NO:2的氨基酸1至19组成。
[60]核酸构建体,其包含编码蛋白质的基因,所述基因可操作地连接于段59的多核苷酸,其中所述基因对于所述多核苷酸是外源的。
[61]重组表达载体,其包含编码蛋白质的基因,所述基因可操作地连接于段59的多核苷酸,其中所述基因对于编码信号肽的多核苷酸是外源的。
[62]重组宿主细胞,其包含编码蛋白质的基因,所述基因可操作地连接于段59的多核苷酸,其中所述基因对于编码信号肽的多核苷酸是外源的。
[63]用于产生蛋白质的方法,其包括:(a)在有助于所述蛋白质产生的条件下培养重组宿主细胞,所述宿主细胞包含编码蛋白质的基因,所述基因可操作地连接于段59的多核苷酸,其中所述基因对于所述编码信号肽的多核苷酸是外源的;和(b)回收所述蛋白质。
[64]组合物,其包含段1-33中任一项的多肽。
[65]降解或转化纤维素材料的方法,其包括:在段1-33中任一项的具有β-葡糖苷酶活性的多肽存在下用酶组合物处理所述纤维素材料。
[66]段65的方法,其中所述纤维素材料经过预处理。
[67]段65或66的方法,其中酶组合物包含选自下组的一种或多种纤维素分解酶:内切葡聚糖酶、纤维二糖水解酶和β-葡糖苷酶。
[68]段65-67中任一项的方法,其中所述酶组合物进一步包含具有纤维素分解增强活性的多肽。
[69]段65-68中任一项的方法,其中所述酶组合物进一步包含一种或多种选自下组的酶:木聚糖酶、半纤维素酶、酯酶、蛋白酶、漆酶或过氧化物酶。
[70]段65-69中任一项的方法,其进一步包括回收已降解的纤维素材料。
[71]段70的方法,其中已降解的纤维素材料是糖。
[72]段71的方法,其中所述糖选自下组:葡萄糖、木糖、甘露糖、半乳糖,和阿拉伯糖。
[73]产生发酵产物的方法,其包括:(a)在段1-33中任一项的具有β-葡糖苷酶活性的多肽存在下,用酶组合物糖化纤维素材料;(b)用一种或多种发酵微生物发酵经糖化的纤维素材料以产生发酵产物;和(c)从所述发酵回收所述发酵产物。
[74]段73的方法,其中所述纤维素材料经预处理。
[75]段73或74的方法,其中所述酶组合物包含选自下组的一种或多种纤维素分解酶:内切葡聚糖酶、纤维二糖水解酶和β-葡糖苷酶。
[76]段73-75中任一项的方法,其中所述酶组合物进一步包含具有纤维素分解增强活性的多肽。
[77]段73-76中任一项的方法,其中所述酶组合物进一步包含选自下组的一种或多种酶:木聚糖酶、半纤维素酶、酯酶、蛋白酶、漆酶或过氧化物酶。
[78]段73-77中任一项的方法,其中步骤(a)和(b)在同步糖化和发酵中同时进行。
[79]段73-78中任一项的方法,其中发酵产物是醇、有机酸、酮、氨基酸或气体。
[80]发酵纤维素材料的方法,其包括:用一种或多种发酵微生物发酵纤维素材料,其中所述纤维素材料在段1-33中任一项的具有β-葡糖苷酶活性的多肽的存在下用酶组合物糖化。
[81]段80的方法,其中所述纤维素材料的发酵产生发酵产物。
[82]段81的方法,其进一步包括从所述发酵回收发酵产物。
[83]段80-82中任一项的方法,其中所述纤维素材料在糖化前经过预处理。
[84]段80-83中任一项的方法,其中所述酶组合物包含选自下组的一种或多种纤维素分解酶:内切葡聚糖酶、纤维二糖水解酶和β-葡糖苷酶。
[85]段80-84中任一项的方法,其中所述酶组合物进一步包含具有纤维素分解增强活性的多肽。
[86]段80-85中任一项的方法,其中所述酶组合物进一步包含选自下组的一种或多种酶:木聚糖酶、半纤维素酶、酯酶、蛋白酶、漆酶或过氧化物酶。
[88]段80-86中任一项的方法,其中发酵产物是醇、有机酸、酮、氨基酸,或气体。
本文描述和要求保护的本发明并不局限于本文公开的具体方面的范围内,因为这些方面旨在作为本发明几个方面的说明。旨在将任何等同的方面包含于本发明的范围内。实际上,从前面的说明中,除本文所显示和描述的之外,本发明的多种修改对于本领域的技术人员来说是显而易见的。这些修改也旨在落入所附的权利要求的范围内。在冲突的情况下,将以包括定义部分的本公开为准。


































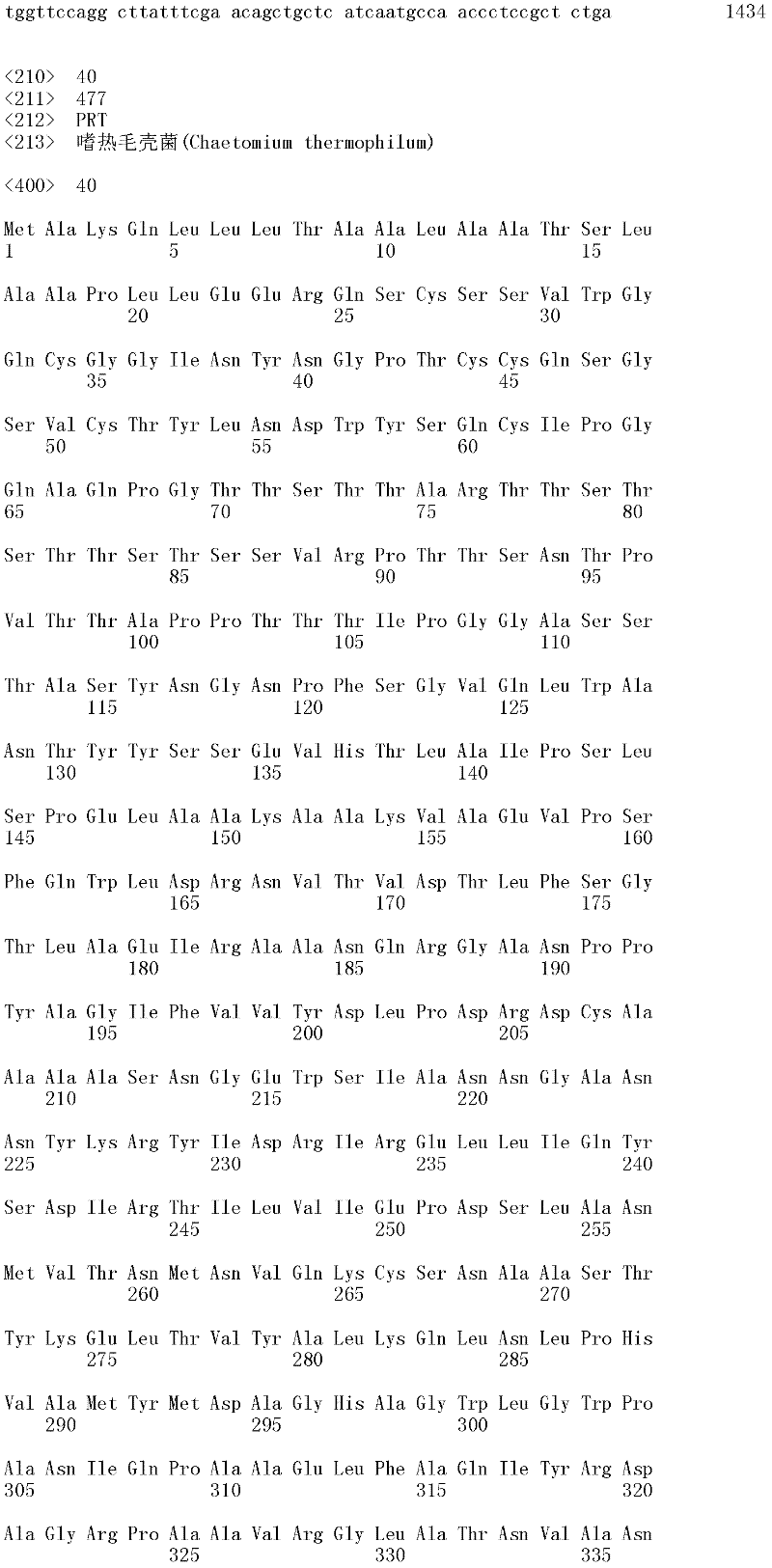

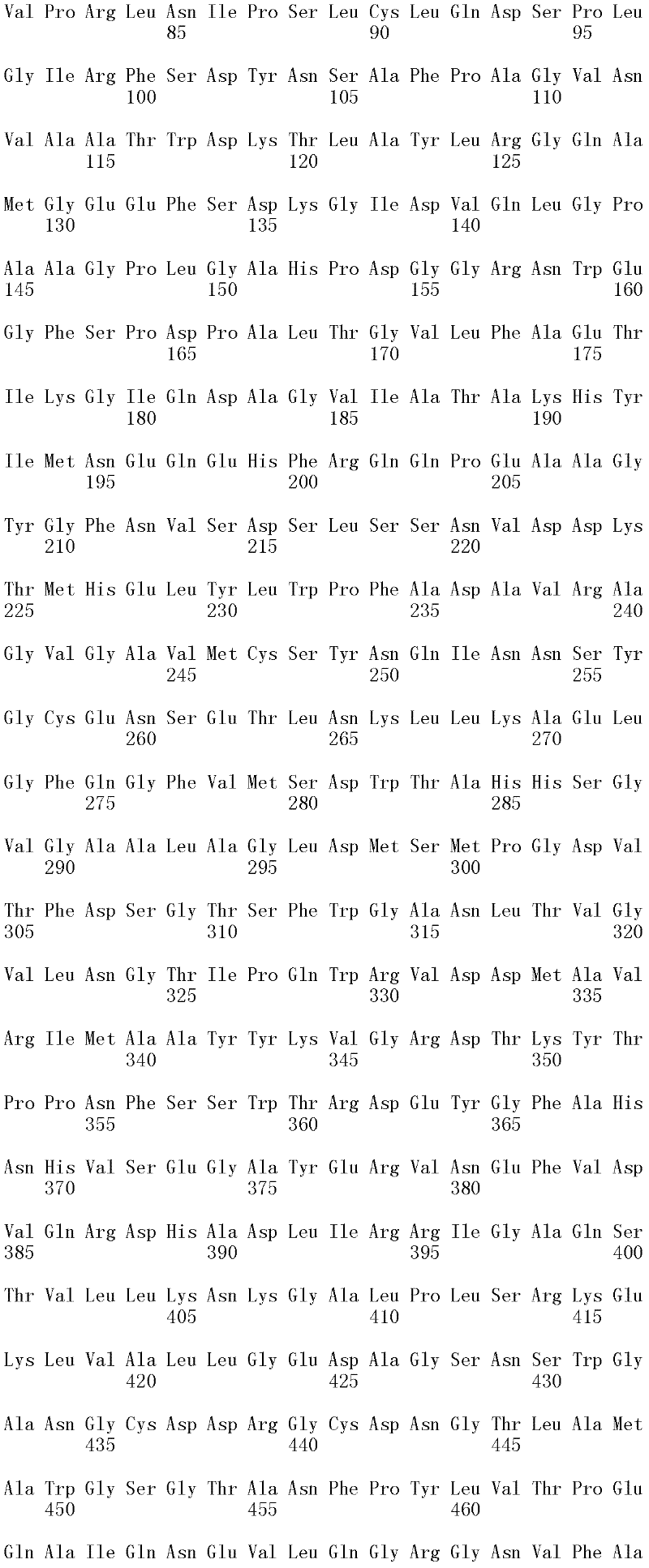







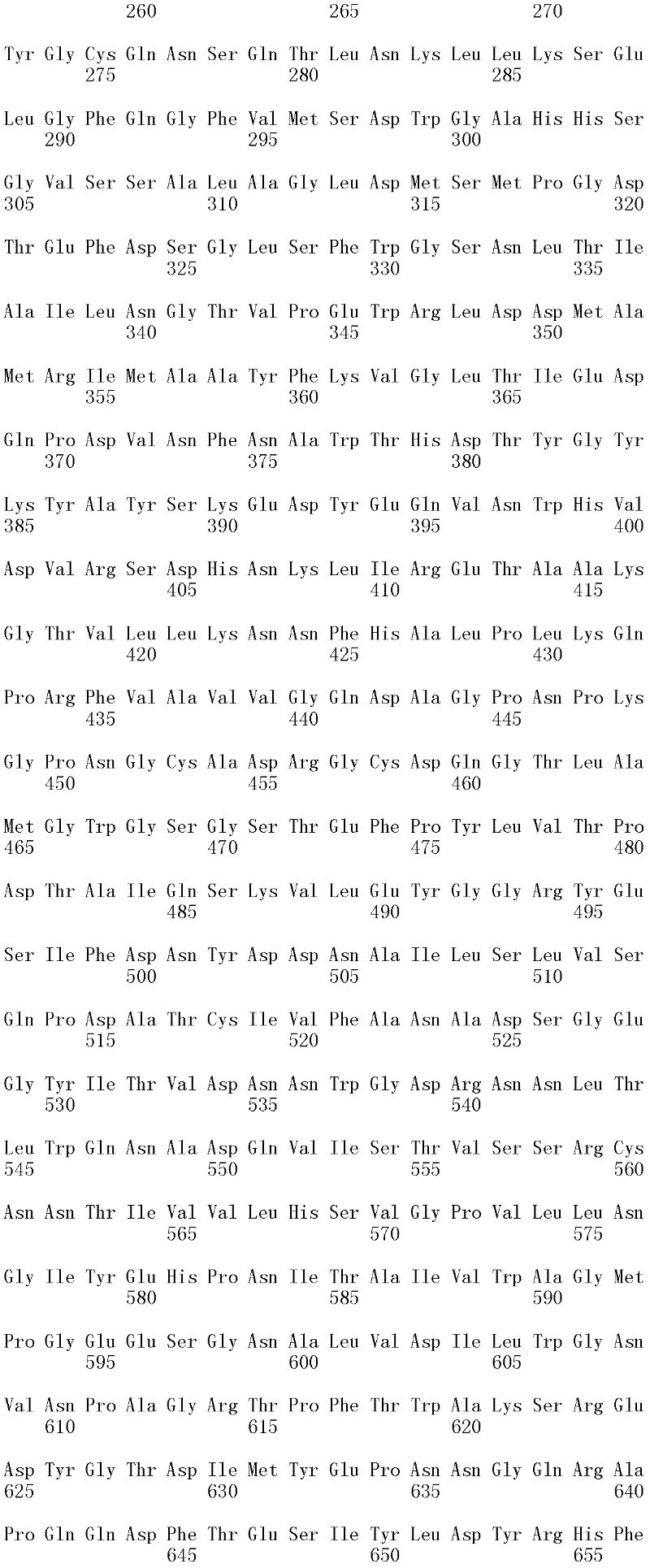

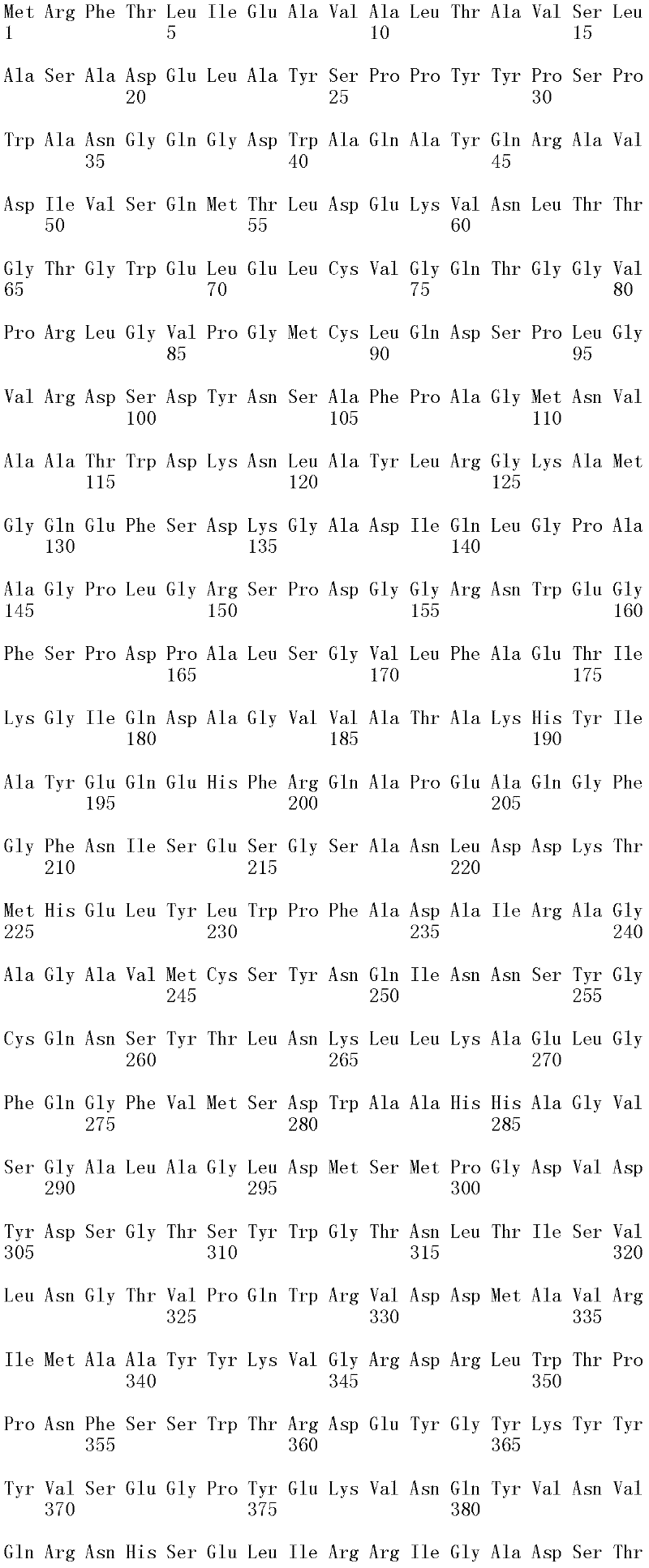








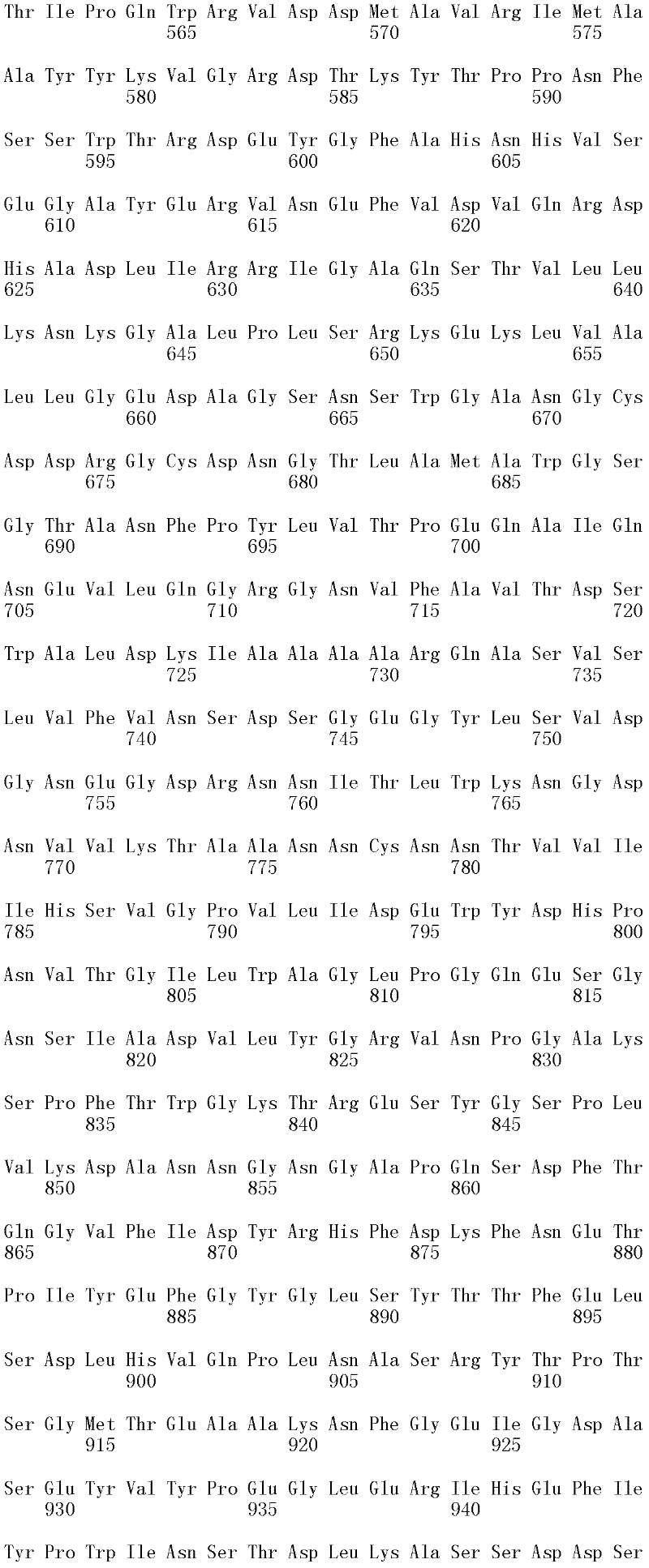

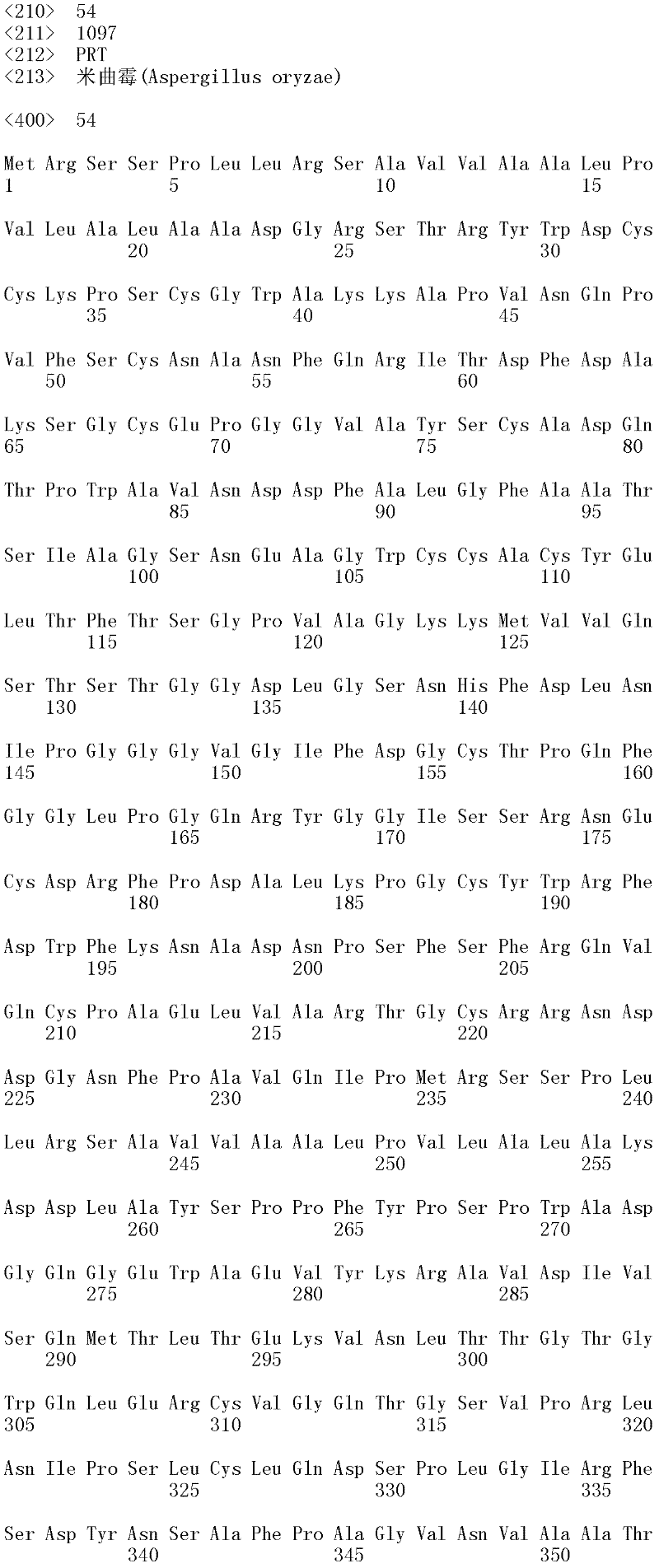



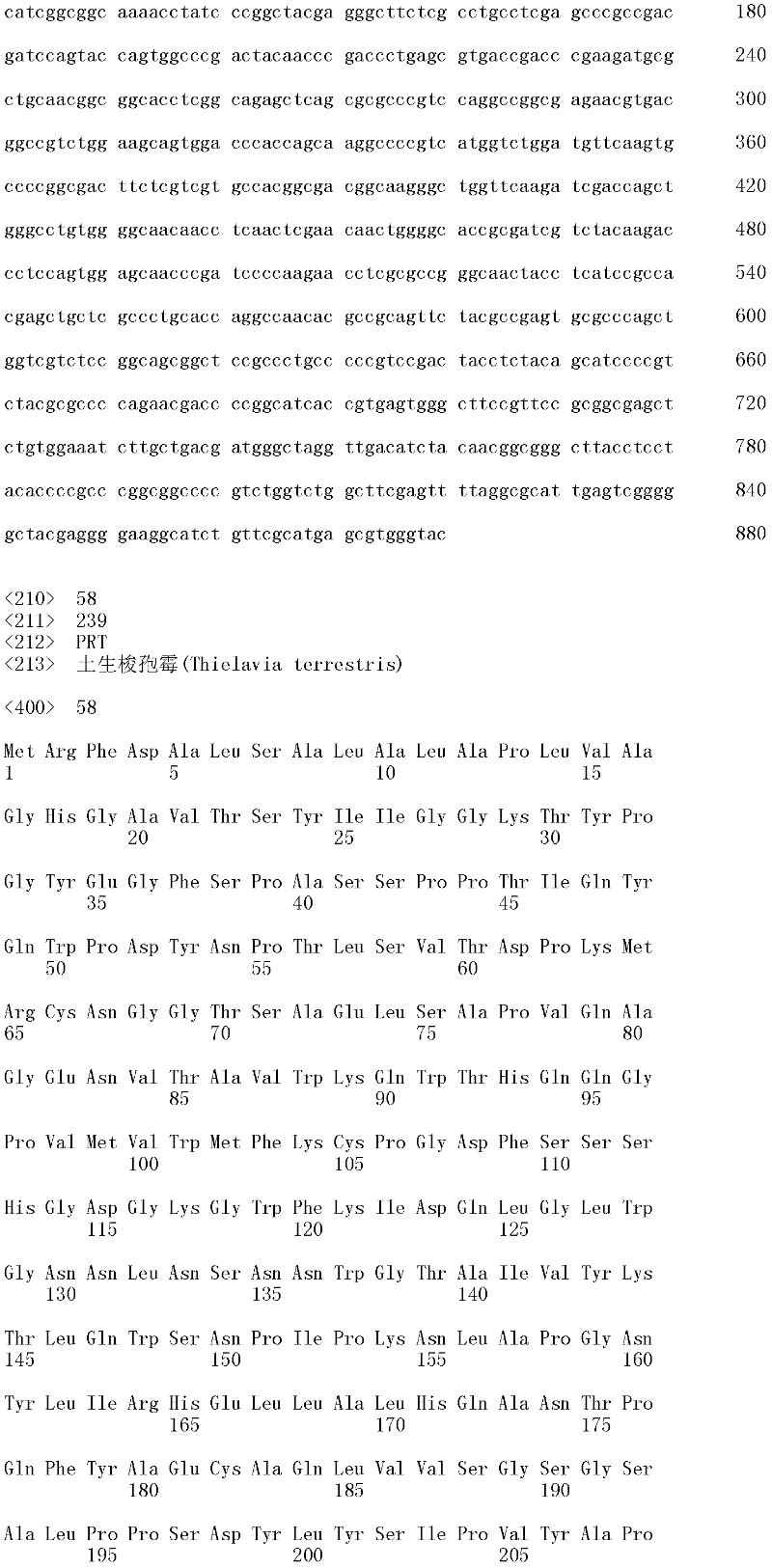










Claims (20)
1.具有β-葡糖苷酶活性的分离的多肽,其选自下组:
(a)包含氨基酸序列的多肽,所述氨基酸序列与SEQ ID NO:2的成熟多肽具有至少60%序列同一性;
(b)由多核苷酸编码的多肽,所述多核苷酸在至少中严格条件下与以下杂交:(i)SEQ ID NO:1的成熟多肽编码序列,(ii)包含SEQ ID NO:1的成熟多肽编码序列的基因组DNA序列,或(iii)(i)或(ii)的全长互补链;
(c)由包含核苷酸序列的多核苷酸编码的多肽,所述核苷酸序列与SEQID NO:1的成熟多肽编码序列具有至少60%序列同一性;和
(d)SEQ ID NO:2的成熟多肽的包含一个或多个(几个)氨基酸的取代、缺失和/或插入的变体。
2.权利要求1的多肽,其包含SEQ ID NO:2的氨基酸序列或其具有β-葡糖苷酶活性的片段,或由SEQ ID NO:2的氨基酸序列或其具有β-葡糖苷酶活性的片段组成。
3.权利要求1的多肽,所述多肽由下述质粒中所含的多核苷酸编码:包含在大肠杆菌NRRL B-50214中的质粒pAHYG-33。
4.分离的多核苷酸,其包含编码权利要求1-3中任一项的多肽的核苷酸序列。
5.用于产生权利要求1-3中任一项的多肽的方法,其包括:(a)在有助于所述多肽产生的条件下培养细胞,所述细胞以其野生型形式产生所述多肽;和(b)回收所述多肽。
6.用于产生权利要求1-3中任一项的多肽的方法,其包括:(a)在有助于所述多肽产生的条件下培养包含核酸构建体的宿主细胞,所述核酸构建体包含编码所述多肽的核苷酸序列;和(b)回收所述多肽。
7.用于产生亲本细胞的突变体的方法,所述方法包括破坏或缺失编码权利要求1-3中任一项的多肽或其部分的多核苷酸,其导致突变体与亲本细胞相比产生较少的所述多肽。
8.用于产生权利要求1-3中任一项的多肽的方法,其包括:(a)在有助于所述多肽产生的条件下培养转基因植物或植物细胞,所述转基因植物或植物细胞包含编码所述多肽的多核苷酸;和(b)回收所述多肽。
9.转基因植物、植物部分或植物细胞,其已经用编码权利要求1-3中任一项的多肽的多核苷酸转化。
10.包含权利要求4的多核苷酸的亚序列的双链抑制RNA(dsRNA)分子,其中任选地所述dsRNA是siRNA或miRNA分子。
11.用于抑制具有β-葡糖苷酶活性的多肽在细胞中表达的方法,其包括对所述细胞施用双链RNA(dsRNA)分子,或者在所述细胞中表达双链RNA(dsRNA)分子,其中所述dsRNA包含权利要求4的多核苷酸的亚序列。
12.分离的多核苷酸,其编码信号肽,所述信号肽包含SEQ ID NO:2的氨基酸1至19或由SEQ ID NO:2的氨基酸1至19组成。
13.用于产生蛋白质的方法,其包括:(a)在有助于所述蛋白质产生的条件下培养重组宿主细胞,所述宿主细胞包含编码蛋白质的基因,所述基因可操作地连接于权利要求12的多核苷酸,其中所述基因对于编码信号肽的多核苷酸是外源的;和(b)回收所述蛋白质。
14.组合物,其包含权利要求1-3中任一项的多肽。
15.用于降解或转化纤维素材料的方法,其包括:在权利要求1-3中任一项的具有β-葡糖苷酶活性的多肽存在下,用酶组合物处理纤维素材料。
16.权利要求15的方法,进一步包括回收已降解的纤维素材料。
17.产生发酵产物的方法,其包括:(a)在权利要求1-3中任一项的具有β-葡糖苷酶活性的多肽存在下,用酶组合物糖化纤维素材料;(b)用一种或多种发酵微生物发酵经糖化的纤维素材料以产生发酵产物;和(c)从所述发酵中回收所述发酵产物。
18.发酵纤维素材料的方法,其包括:用一种或多种发酵微生物发酵纤维素材料,其中在权利要求1-3中任一项的具有β-葡糖苷酶活性的多肽的存在下用酶组合物糖化所述纤维素材料。
19.权利要求18的方法,其中纤维素材料的发酵产生发酵产物。
20.权利要求19的方法,进一步包括从所述发酵回收发酵产物。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US14792309P | 2009-01-28 | 2009-01-28 | |
| US61/147,923 | 2009-01-28 | ||
| PCT/US2010/022405 WO2010088387A1 (en) | 2009-01-28 | 2010-01-28 | Polypeptides having beta-glucosidase activity and polynucleotides encoding same |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN102388134A true CN102388134A (zh) | 2012-03-21 |
Family
ID=42040464
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN2010800153740A Pending CN102388134A (zh) | 2009-01-28 | 2010-01-28 | 具有β-葡糖苷酶活性的多肽和编码该多肽的多核苷酸 |
Country Status (4)
| Country | Link |
|---|---|
| US (1) | US8604277B2 (zh) |
| EP (1) | EP2391715A1 (zh) |
| CN (1) | CN102388134A (zh) |
| WO (1) | WO2010088387A1 (zh) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN104822878A (zh) * | 2012-12-07 | 2015-08-05 | 诺维信公司 | 改进纸浆的排水 |
| CN107002056A (zh) * | 2014-09-05 | 2017-08-01 | 诺维信公司 | 碳水化合物结合模块变体以及编码它们的多核苷酸 |
Families Citing this family (84)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP2569426A4 (en) | 2010-05-14 | 2013-10-09 | Codexis Inc | ZELLBIOHYDROLASE VARIANTS |
| WO2012003379A1 (en) | 2010-06-30 | 2012-01-05 | Novozymes A/S | Polypeptides having beta-glucosidase activity and polynucleotides encoding same |
| EP3470514A1 (en) | 2010-08-30 | 2019-04-17 | Novozymes A/S | Polypeptides having cellulolytic enhancing activity and polynucleotides encoding same |
| US20130212746A1 (en) | 2010-08-30 | 2013-08-15 | Novoyzmes A/S | Polypeptides Having Hemicellulolytic Activity And Polynucleotides Encoding Same |
| EP2611901B1 (en) | 2010-08-30 | 2016-05-11 | Novozymes A/S | Polypeptides having beta-glucosidase activity, beta-xylosidase activity, or beta-glucosidase and beta-xylosidase activity and polynucleotides encoding same |
| US8624082B2 (en) | 2010-08-30 | 2014-01-07 | Novozymes A/S | Polypeptides having xylanase activity and polynucleotides encoding same |
| US9267126B2 (en) | 2010-08-30 | 2016-02-23 | Novozymes, Inc. | Polypeptides having endoglucanase activity and polynucleotides encoding same |
| US9187742B2 (en) | 2010-08-30 | 2015-11-17 | Novozymes, Inc. | Polypeptides having cellobiohydrolase activity and polynucleotides encoding same |
| AU2011302536A1 (en) | 2010-09-15 | 2013-03-14 | The Regents Of The University Of California | Thermophilic mutants of Trichoderma reesei endoglucanase I |
| WO2012059053A1 (en) | 2010-11-04 | 2012-05-10 | Novozymes A/S | Polypeptides having cellobiohydrolase activity and polynucleotides encoding same |
| BR112013009026A2 (pt) | 2010-11-12 | 2016-07-12 | Novozymes Inc | polipeptídeo isolado, célula hospedeira recombinante, métodos para produzir um polipeptídeo, um mutante de uma célula precursora, uma proteína, e um produto de fermentação, para inibir a expressão de um polipeptídeo, para degradar ou converter um material celulósico, e método para fermentar um material celulósico, planta transgênica, parte de planta ou célula de planta, molécula de rna inibidora de filamento duplo, polinucleotídeo isolado, e, formulação de caldo integral ou composição de cultura de célula |
| CN103534348B (zh) | 2011-01-26 | 2018-04-10 | 诺维信公司 | 具有纤维二糖水解酶活性的多肽和编码该多肽的多核苷酸 |
| US9080161B2 (en) | 2011-01-26 | 2015-07-14 | Novozymes, Inc. | Polypeptides having cellobiohydrolase activity and polynucleotides encoding same |
| MX2013007720A (es) | 2011-01-26 | 2013-08-09 | Novozymes As | Polipeptidos que tienen actividad celobiohidrolasa y polinucleotidos que codifican para los mismos. |
| US8759023B2 (en) | 2011-01-26 | 2014-06-24 | Novozymes Inc. | Polypeptides having cellobiohydrolase activity and polynucleotides encoding same |
| BR112013016830A2 (pt) | 2011-02-23 | 2017-03-01 | Novozymes Inc | polipeptídeo isolado, polinucleotídeo isolado, método de produzir o polipeptídeo, de produzir um mutante de uma célula parental, de inibir a expressão de um polipeptídeo, de produzir uma proteína, de degradar ou converter um material celulósico, de produzir um produto de fermentação e de fermentar um material celulósico, planta transgênica, parte da planta ou célula de planta transformada com um polinucleotídeo, molécula de rna de fita dupla, composição, e, formulação de caldo completo ou composição de cultura de células |
| EP2683830A1 (en) | 2011-03-09 | 2014-01-15 | Novozymes A/S | Methods of increasing the cellulolytic enhancing activity of a polypeptide |
| WO2012122477A1 (en) | 2011-03-10 | 2012-09-13 | Novozymes A/S | Polypeptides having cellulolytic enhancing activity and polynucleotides encoding same |
| US20140080182A1 (en) | 2011-03-31 | 2014-03-20 | Novozymes, Inc. | Cellulose Binding Domain Variants and Polynucleotides Encoding Same |
| US9410136B2 (en) | 2011-03-31 | 2016-08-09 | Novozymes, Inc. | Methods for enhancing the degradation or conversion of cellulosic material |
| US20120276594A1 (en) * | 2011-04-27 | 2012-11-01 | Codexis, Inc. | Cellobiohydrolase variants |
| CA2834513A1 (en) | 2011-04-28 | 2012-11-01 | Novozymes, Inc. | Polypeptides having endoglucanase activity and polynucleotides encoding same |
| EP2702162B1 (en) | 2011-04-29 | 2020-02-26 | Novozymes, Inc. | Methods for enhancing the degradation or conversion of cellulosic material |
| CN109022518A (zh) | 2011-07-22 | 2018-12-18 | 诺维信北美公司 | 用于预处理纤维素材料和改进其水解的方法 |
| DK3091073T4 (da) | 2011-08-04 | 2023-06-06 | Novozymes Inc | Polypeptider med xylanaseaktivitet og polynukleotider, der koder for dem |
| EP2739728B1 (en) | 2011-08-04 | 2017-07-12 | Novozymes A/S | Polypeptides having endoglucanase activity and polynucleotides encoding same |
| BR112014003740A2 (pt) | 2011-08-23 | 2018-08-14 | Codexis Inc | variantes da celobiohidrolase |
| AU2012298713B2 (en) | 2011-08-24 | 2017-11-23 | Novozymes, Inc. | Methods for obtaining positive transformants of a filamentous fungal host cell |
| IN2014CN02136A (zh) | 2011-08-24 | 2015-05-29 | Novozymes Inc | |
| BR112014005290B1 (pt) * | 2011-09-09 | 2021-11-09 | Novozymes A/S | Método para melhoramento da resistência do papel |
| US20140308705A1 (en) | 2011-09-20 | 2014-10-16 | Novozymes A/S | Polypeptides Having Cellulolytic Enhancing Activity And Polynucleotides Encoding Same |
| US20140329284A1 (en) | 2011-09-30 | 2014-11-06 | Novozymes, Inc. | Chimeric Polypeptides Having Beta-Glucosidase Activity and Polynucleotides Encoding Same |
| EP2773656B1 (en) | 2011-10-31 | 2019-06-19 | Novozymes, Inc. | Polypeptides having cellulolytic enhancing activity and polynucleotides encoding same |
| CN102399769B (zh) * | 2011-11-16 | 2013-05-01 | 广东溢多利生物科技股份有限公司 | 一种优化改良的中性纤维素酶meg1及其基因和应用 |
| EP3409769A1 (en) | 2011-11-18 | 2018-12-05 | Novozymes A/S | Polypeptides having beta-glucosidase activity, beta-xylosidase activity, or beta-glucosidase and beta-xylosidase activity and polynucleotides encoding same |
| EP3219794B1 (en) | 2011-11-21 | 2019-09-11 | Novozymes A/S | Gh61 polypeptide variants and polynucleotides encoding same |
| EP2782998B1 (en) | 2011-11-22 | 2018-01-10 | Novozymes Inc. | Polypeptides having beta-xylosidase activity and polynucleotides encoding same |
| DK2785732T3 (en) | 2011-12-01 | 2017-06-19 | Novozymes Inc | POLYPEPTIDES WITH BETA-XYLOSIDASE ACTIVITY AND POLYNUCLEOTIDES CODING THEM |
| DK2791330T3 (da) | 2011-12-16 | 2017-11-06 | Novozymes Inc | Polypeptider med laccaseaktivitet og polynukleotider, som koder for dem |
| BR112014014697A2 (pt) | 2011-12-19 | 2020-10-27 | Novozymes, Inc. | polipeptídeo isolado, composição, polinucleotídeo isolado, construto de ácido nucleico ou vetor de expressão, célula hospedeira recombinante, métodos para produzir um polipeptídeo e uma proteína, para gerar oxigênio molecular, e para remover peróxido de hidrogênio do tecido, processos para degradar ou converter um material celulósico, e para produzir um produto de fermentação, e, formulação de caldo integral ou composição de cultura de células |
| CA2859796A1 (en) | 2011-12-19 | 2013-06-27 | Novozymes A/S | Processes and compositions for increasing the digestibility of cellulosic materials |
| DK2794869T3 (en) | 2011-12-20 | 2018-01-02 | Novozymes Inc | CELLOBIO HYDROLASE VARIABLES AND POLYNUCLEOTIDES CODING THEM |
| EP2794899A1 (en) | 2011-12-21 | 2014-10-29 | Novozymes, Inc. | Methods for determining the degradation of a biomass material |
| CN104245929A (zh) * | 2012-04-23 | 2014-12-24 | 诺维信公司 | 具有α-葡糖醛酸糖苷酶活性的多肽以及编码其的多核苷酸 |
| CA2868308A1 (en) | 2012-04-27 | 2013-10-31 | Novozymes, Inc. | Gh61 polypeptide variants and polynucleotides encoding same |
| CN104364371B (zh) | 2012-05-31 | 2018-10-23 | 诺维信公司 | 具有有机磷水解酶活性的多肽 |
| BR112015005884A2 (pt) | 2012-09-19 | 2017-08-08 | Novozymes Inc E Novozymes As | processos para degradação de um material celulósico, para produção de um produto da fermentação, e de fermentação de um material celulósico, composição, e, formulação de caldo inteiro ou composição de cultura de células |
| CN105861469B (zh) | 2012-10-08 | 2020-04-14 | 诺维信公司 | 具有纤维素分解增强活性的多肽以及编码它们的多核苷酸 |
| WO2014066141A2 (en) | 2012-10-24 | 2014-05-01 | Novozymes A/S | Polypeptides having cellulolytic enhancing activity and polynucleotides encoding same |
| WO2014093275A1 (en) | 2012-12-12 | 2014-06-19 | Danisco Us Inc. | Variants of cellobiohydrolases |
| CN104870639A (zh) | 2012-12-14 | 2015-08-26 | 诺维信公司 | 具有纤维素分解增强活性的多肽以及编码它们的多核苷酸 |
| WO2014099798A1 (en) | 2012-12-19 | 2014-06-26 | Novozymes A/S | Polypeptides having cellulolytic enhancinc activity and polynucleotides encoding same |
| CN105164254B (zh) | 2013-03-08 | 2019-06-28 | 诺维信公司 | 纤维二糖水解酶变体和编码它们的多核苷酸 |
| EP2994529B1 (en) | 2013-05-10 | 2018-11-21 | Novozymes A/S | Polypeptides having xylanase activity and polynucleotides encoding same |
| CN105492601A (zh) | 2013-09-04 | 2016-04-13 | 诺维信公司 | 增加对纤维素材料的酶法水解的方法 |
| FR3013731B1 (fr) * | 2013-11-22 | 2017-09-01 | Ifp Energies Now | Variants d'endoglucanases a activite amelioree et leurs utilisations |
| MY186318A (en) | 2014-01-07 | 2021-07-08 | Novozymes As | Process for degrading mannan-containing cellulosic materials |
| EP3186372B1 (en) | 2014-08-28 | 2020-10-21 | Novozymes A/S | Solubilization of msw with blend enzymes |
| WO2016045569A1 (en) | 2014-09-23 | 2016-03-31 | Novozymes A/S | Processes for producing ethanol and fermenting organisms |
| WO2016138167A2 (en) | 2015-02-24 | 2016-09-01 | Novozymes A/S | Cellobiohydrolase variants and polynucleotides encoding same |
| WO2016145350A1 (en) | 2015-03-12 | 2016-09-15 | Novozymes A/S | Multi-stage enzymatic hydrolysis of lignocellulosic biomass |
| US10513721B2 (en) | 2015-03-12 | 2019-12-24 | Novozymes A/S | Two-stage process for enzymatic hydrolysis of lignocellulosic biomass |
| BR112017019331A2 (pt) | 2015-03-12 | 2018-06-05 | Beta Renewables Spa | processos para sacarificação de múltiplos estágios de um material lignocelulósico e para melhorar um rendimento de glicose ou xilose de sacarificação de um material lignocelulósico em um reator de tanque continuamente agitado |
| EP3067428A1 (en) | 2015-03-12 | 2016-09-14 | BETA RENEWABLES S.p.A. | A process for producing a hydrolyzed mixture from a pre-treated ligno-cellulosic slurry comprising a slurry liquid and slurry solids |
| EP3303578B1 (en) | 2015-05-27 | 2020-07-08 | Novozymes A/S | Polypeptides having cellobiohydrolase activity and polynucleotides encoding same |
| WO2017019491A1 (en) | 2015-07-24 | 2017-02-02 | Novozymes Inc. | Polypeptides having beta-xylosidase activity and polynucleotides encoding same |
| EP3325616A1 (en) | 2015-07-24 | 2018-05-30 | Novozymes, Inc. | Polypeptides having arabinofuranosidase activity and polynucleotides encoding same |
| WO2017040907A1 (en) | 2015-09-04 | 2017-03-09 | Novozymes A/S | Methods of inhibiting aa9 lytic polysaccharide monooxygenase catalyzed inactivation of enzyme compositions |
| DK3353195T3 (da) | 2015-09-22 | 2021-11-22 | Novozymes As | Polypeptider med cellobiohydrolaseaktivitet og polynukleotider, der koder for dem |
| WO2017070219A1 (en) | 2015-10-20 | 2017-04-27 | Novozymes A/S | Lytic polysaccharide monooxygenase (lpmo) variants and polynucleotides encoding same |
| WO2017076421A1 (en) | 2015-11-02 | 2017-05-11 | Renescience A/S | Solubilization of msw with blend enzymes |
| DK3393983T3 (da) | 2015-12-23 | 2023-12-18 | Envirozyme Llc | Fremgangsmåder til forøgelse af slams afvandingsevne med enzymbehandling |
| EP3416740B1 (en) | 2016-02-19 | 2021-01-06 | Intercontinental Great Brands LLC | Processes to create multiple value streams from biomass sources |
| DK3423577T3 (da) | 2016-03-02 | 2024-07-08 | Novozymes As | Cellobiohydrolasevarianter og polynukleotider til kodning deraf |
| WO2017165760A1 (en) | 2016-03-24 | 2017-09-28 | Novozymes A/S | Cellobiohydrolase variants and polynucleotides encoding same |
| WO2017205535A1 (en) | 2016-05-27 | 2017-11-30 | Novozymes, Inc. | Polypeptides having endoglucanase activity and polynucleotides encoding same |
| WO2018026868A1 (en) | 2016-08-01 | 2018-02-08 | Novozymes, Inc. | Polypeptides having endoglucanase activity and polynucleotides encoding same |
| WO2018085370A1 (en) | 2016-11-02 | 2018-05-11 | Novozymes A/S | Processes for reducing production of primeverose during enzymatic saccharification of lignocellulosic material |
| WO2019074828A1 (en) | 2017-10-09 | 2019-04-18 | Danisco Us Inc | CELLOBIOSE DEHYDROGENASE VARIANTS AND METHODS OF USE |
| EP3781697A1 (en) | 2018-04-20 | 2021-02-24 | Renescience A/S | Method for determining chemical compounds in waste |
| CN113302293A (zh) | 2018-12-12 | 2021-08-24 | 诺维信公司 | 具有木聚糖酶活性的多肽以及编码这些多肽的多核苷酸 |
| AU2020326511A1 (en) | 2019-08-06 | 2022-01-06 | Novozymes A/S | Fusion proteins for improved enzyme expression |
| JP2023547177A (ja) | 2020-11-04 | 2023-11-09 | レネサイエンス エー/エス | プロセス水の再循環を含む廃棄物の酵素処理及び/又は微生物処理のための方法 |
| EP4240542A1 (en) | 2020-11-04 | 2023-09-13 | Renescience A/S | Method for sanitizing waste |
Family Cites Families (93)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| DK187280A (da) | 1980-04-30 | 1981-10-31 | Novo Industri As | Ruhedsreducerende middel til et fuldvaskemiddel fuldvaskemiddel og fuldvaskemetode |
| DK122686D0 (da) | 1986-03-17 | 1986-03-17 | Novo Industri As | Fremstilling af proteiner |
| EP0406314B1 (en) | 1988-03-24 | 1993-12-01 | Novo Nordisk A/S | A cellulase preparation |
| US5776757A (en) | 1988-03-24 | 1998-07-07 | Novo Nordisk A/S | Fungal cellulase composition containing alkaline CMC-endoglucanase and essentially no cellobiohydrolase and method of making thereof |
| US5223409A (en) | 1988-09-02 | 1993-06-29 | Protein Engineering Corp. | Directed evolution of novel binding proteins |
| US5110735A (en) | 1989-09-26 | 1992-05-05 | Midwest Research Institute | Thermostable purified endoglucanase from thermophilic bacterium acidothermus cellulolyticus |
| US5536655A (en) | 1989-09-26 | 1996-07-16 | Midwest Research Institute | Gene coding for the E1 endoglucanase |
| US5275944A (en) | 1989-09-26 | 1994-01-04 | Midwest Research Institute | Thermostable purified endoglucanas from acidothermus cellulolyticus ATCC 43068 |
| NZ237549A (en) | 1990-03-23 | 1993-06-25 | Gist Brocades Nv | Production of enhanced levels of enzymes in the seeds of transgenic plants and the use of these seeds |
| AU639570B2 (en) | 1990-05-09 | 1993-07-29 | Novozymes A/S | A cellulase preparation comprising an endoglucanase enzyme |
| DK115890D0 (da) | 1990-05-09 | 1990-05-09 | Novo Nordisk As | Enzym |
| US6395966B1 (en) | 1990-08-09 | 2002-05-28 | Dekalb Genetics Corp. | Fertile transgenic maize plants containing a gene encoding the pat protein |
| IL99552A0 (en) | 1990-09-28 | 1992-08-18 | Ixsys Inc | Compositions containing procaryotic cells,a kit for the preparation of vectors useful for the coexpression of two or more dna sequences and methods for the use thereof |
| SG52693A1 (en) | 1991-01-16 | 1998-09-28 | Procter & Gamble | Detergent compositions with high activity cellulase and softening clays |
| EP1431389A3 (en) | 1992-10-06 | 2004-06-30 | Novozymes A/S | Cellulase variants |
| ATE412048T1 (de) | 1993-03-10 | 2008-11-15 | Novozymes As | Enzyme mit xylanaseaktivität aus aspergillus aculeatus |
| DE4343591A1 (de) | 1993-12-21 | 1995-06-22 | Evotec Biosystems Gmbh | Verfahren zum evolutiven Design und Synthese funktionaler Polymere auf der Basis von Formenelementen und Formencodes |
| US5605793A (en) | 1994-02-17 | 1997-02-25 | Affymax Technologies N.V. | Methods for in vitro recombination |
| DE69536145D1 (de) | 1994-03-08 | 2011-04-07 | Novozymes As | Neuartige alkalische Zellulasen |
| DK0765394T3 (da) | 1994-06-03 | 2001-12-10 | Novozymes As | Oprensede Myceliopthora-laccaser og nukleinsyrer der koder for disse |
| DE69534185T2 (de) | 1994-06-30 | 2006-02-23 | Novozymes Biotech, Inc., Davis | Nicht-toxisches, nicht-toxigenes, nicht-pathogenes fusarium expressionssystem und darin zu verwendende promotoren und terminatoren |
| EP1995303A3 (en) | 1994-10-06 | 2008-12-31 | Novozymes A/S | Enzyme preparation with endoglucanase activity |
| CN1182451A (zh) | 1995-03-17 | 1998-05-20 | 诺沃挪第克公司 | 新的内切葡聚糖酶 |
| US6313081B1 (en) | 1995-04-28 | 2001-11-06 | Henkel Kommanditgesellschaft Auf Aktien (Kgaa) | Detergents comprising cellulases |
| US20030044956A1 (en) | 1995-08-23 | 2003-03-06 | Short Jay M. | Enzymes having carboxymethyl cellulase activity and methods of use thereof |
| CA2232245C (en) | 1995-10-17 | 2011-06-07 | Rohm Enzyme Finland Oy | Cellulases, the genes encoding them and uses thereof |
| WO1998008940A1 (en) | 1996-08-26 | 1998-03-05 | Novo Nordisk A/S | A novel endoglucanase |
| DE69735767T2 (de) | 1996-09-17 | 2007-04-05 | Novozymes A/S | Cellulasevarianten |
| US6451063B1 (en) | 1996-09-25 | 2002-09-17 | Genencor International, Inc. | Cellulase for use in industrial processes |
| US6017870A (en) | 1996-10-09 | 2000-01-25 | Genencor International, Inc. | Purified cellulase and method of producing |
| US5811381A (en) | 1996-10-10 | 1998-09-22 | Mark A. Emalfarb | Cellulase compositions and methods of use |
| US7883872B2 (en) | 1996-10-10 | 2011-02-08 | Dyadic International (Usa), Inc. | Construction of highly efficient cellulase compositions for enzymatic hydrolysis of cellulose |
| US5989899A (en) | 1996-12-23 | 1999-11-23 | Genencor International, Inc. | Oversized cellulase compositions for use in detergent compositions and in the treatment of textiles |
| WO1999006574A1 (en) | 1997-07-31 | 1999-02-11 | Dsm N.V. | Cellulose degrading enzymes of aspergillus |
| US5871550A (en) | 1997-08-26 | 1999-02-16 | Genencor International, Inc. | Mutant Thermonospora spp. cellulase |
| EP1034280B1 (en) | 1997-11-19 | 2006-05-31 | Genencor International, Inc. | Cellulase produced by actinomycetes and method of producing same |
| JP3661995B2 (ja) | 1997-11-19 | 2005-06-22 | ジェネンコア インターナショナル インコーポレーテッド | 放線菌類により産生されるセルラーゼとその産生方法 |
| US6562612B2 (en) | 1997-11-19 | 2003-05-13 | Genencor International, Inc. | Cellulase producing actinomycetes, cellulase produced therefrom and method of producing same |
| EP1038008B1 (en) | 1997-12-16 | 2006-10-18 | Genencor International, Inc. | Methods for producing egiii-like enzymes |
| US6506559B1 (en) | 1997-12-23 | 2003-01-14 | Carnegie Institute Of Washington | Genetic inhibition by double-stranded RNA |
| EP1080210A2 (en) | 1998-05-27 | 2001-03-07 | Novo Nordisk Biotech, Inc. | Methods for producing a polypeptide by modifying the copy number of a gene |
| EP1088080B1 (en) | 1998-06-24 | 2006-10-11 | Genencor International, Inc. | Recombinant production of cellulase from actinomycetes |
| EP1124949B1 (en) | 1998-10-26 | 2006-07-12 | Novozymes A/S | Constructing and screening a dna library of interest in filamentous fungal cells |
| US6511824B1 (en) | 1999-03-17 | 2003-01-28 | Exelixis, Inc. | Nucleic acids and polypeptides of invertebrate TWIK channels and methods of use |
| CN1940067A (zh) | 1999-03-22 | 2007-04-04 | 诺沃奇梅兹有限公司 | 用于在真菌细胞中表达基因的启动子 |
| CA2372594A1 (en) | 1999-05-19 | 2000-11-23 | Midwest Research Institute | E1 endoglucanase variants y245g, y82r and w42r |
| US6531644B1 (en) | 2000-01-14 | 2003-03-11 | Exelixis, Inc. | Methods for identifying anti-cancer drug targets |
| ES2166316B1 (es) | 2000-02-24 | 2003-02-16 | Ct Investig Energeticas Ciemat | Procedimiento de produccion de etanol a partir de biomasa lignocelulosica utilizando una nueva levadura termotolerante. |
| DE60134400D1 (de) | 2000-04-07 | 2008-07-24 | Novozymes As | Einfangen von signalsequenzen |
| ES2423898T3 (es) | 2000-07-21 | 2013-09-25 | The Government Of The United States Of America, As Represented By The Secretary, Department Of Health And Human Services | Citoblastos de pulpa dental de seres humanos adultos in vitro e in vivo |
| AU2001292292A1 (en) * | 2000-09-29 | 2002-04-08 | Meiji Seika Kaisha Ltd. | Novel enzyme having beta-glucosidase activity and use thereof |
| US6515109B1 (en) | 2000-10-12 | 2003-02-04 | Exelixis, Inc. | Human ECT2 polypeptide |
| US7151204B2 (en) | 2001-01-09 | 2006-12-19 | Monsanto Technology Llc | Maize chloroplast aldolase promoter compositions and methods for use thereof |
| AU2002316785A1 (en) | 2001-05-18 | 2002-12-03 | Novozymes A/S | Polypeptides having cellobiase activity and polynucleotides encoding same |
| US6982159B2 (en) | 2001-09-21 | 2006-01-03 | Genencor International, Inc. | Trichoderma β-glucosidase |
| DK1448595T3 (da) | 2001-11-20 | 2007-02-19 | Novozymes As | Antimikrobielle polypeptider |
| US7045332B2 (en) | 2001-12-18 | 2006-05-16 | Genencor International, Inc. | BGL4 β-glucosidase and nucleic acids encoding the same |
| US7045331B2 (en) | 2001-12-18 | 2006-05-16 | Genencor International, Inc. | EGVII endoglucanase and nucleic acids encoding the same |
| US7049125B2 (en) | 2001-12-18 | 2006-05-23 | Genencor International, Inc. | EGVIII endoglucanase and nucleic acids encoding the same |
| US7005289B2 (en) | 2001-12-18 | 2006-02-28 | Genencor International, Inc. | BGL5 β-glucosidase and nucleic acids encoding the same |
| US7056721B2 (en) | 2001-12-18 | 2006-06-06 | Genencor International, Inc. | EGVI endoglucanase and nucleic acids encoding the same |
| BR0306740A (pt) | 2002-01-23 | 2004-12-28 | Royal Nedalco B V | Célula hospedeira transformada com um construto de ácido nucléico, molécula de ácido nucleico isolada, e, processos para a produção de etanol, e de um produto de fermentação |
| DK2322606T3 (en) | 2002-08-16 | 2015-12-14 | Danisco Us Inc | NOVEL HYPROCREA JECORINA CBH1 cellulase WITH INCREASED THERMAL STABILITY COMPREHENSIVE SUBSTITUTION OR DELETION FOR POSITION T332 |
| EP1556512B1 (en) | 2002-11-07 | 2016-06-15 | Danisco US Inc. | Bgl6 beta-glucosidase and nucleic acids encoding the same |
| US7407788B2 (en) | 2002-11-21 | 2008-08-05 | Danisco A/S, Genencor Division | BGL7 beta-glucosidase and nucleic acids encoding the same |
| CN1898381B (zh) | 2003-03-21 | 2013-07-17 | 金克克国际有限公司 | Cbh1同源物和cbh1纤维素酶变体 |
| CA2520636C (en) | 2003-04-01 | 2012-05-08 | Genencor International, Inc. | Variant humicola grisea cbh1.1 |
| DK1862626T3 (da) | 2003-05-29 | 2011-12-12 | Genencor Int | Nye Trichoderma-gener |
| WO2005047499A1 (en) | 2003-10-28 | 2005-05-26 | Novozymes Inc. | Polypeptides having beta-glucosidase activity and polynucleotides encoding same |
| WO2006011899A1 (en) | 2003-11-25 | 2006-02-02 | L-3 Communications Security and Detection Systems Corporation | Security system for detecting nuclear masses |
| WO2005074647A2 (en) | 2004-01-30 | 2005-08-18 | Novozymes Inc. | Polypeptides having cellulolytic enhancing activity and polynucleotides encoding same |
| CN1965078B (zh) | 2004-02-06 | 2013-09-18 | 诺维信股份有限公司 | 具有增强分解纤维素活性的多肽和编码所述多肽的多核苷酸 |
| CN101389645B (zh) | 2004-02-12 | 2016-08-03 | 诺维信股份有限公司 | 具有木聚糖酶活性的多肽和编码它的多核苷酸 |
| US8097445B2 (en) | 2004-03-25 | 2012-01-17 | Danisco Us Inc. | Exo-endo cellulase fusion protein |
| CA2560588A1 (en) | 2004-03-25 | 2005-10-06 | Genencor International, Inc. | Cellulase fusion protein and heterologous cellulase fusion construct encoding the same |
| US20060004699A1 (en) | 2004-06-30 | 2006-01-05 | Nokia Corporation | Method and system for managing metadata |
| DK1838849T3 (en) | 2004-12-30 | 2015-11-30 | Danisco Us Inc | CBH2 CELLULASE VARIETIES OF HYPOCREA JECORINA |
| CN101160405B (zh) | 2005-04-12 | 2014-01-01 | 纳幕尔杜邦公司 | 处理生物质以获得目标化学物质 |
| BRPI0609906A8 (pt) | 2005-04-29 | 2017-11-21 | Ab Enzymes Oy | proteína de fusão de celulase, seu processo de produção e composição detergente contendo a mesma, vetor de expressão, célula hospedeira, preparação de enzima, processos para desbotamento e para bioacabamento e métodos de tratamento de fibra celulósica contendo material têxtil, para tratar fibra ou polpa derivada de madeira e para melhorar a qualidade de ração animal |
| JP2007028912A (ja) | 2005-07-22 | 2007-02-08 | Gekkeikan Sake Co Ltd | 新規β−グルコシダーゼ、及びその利用 |
| WO2007019442A2 (en) | 2005-08-04 | 2007-02-15 | Novozymes, Inc. | Polypeptides having beta-glucosidase activity and polynucleotides encoding same |
| JP5129142B2 (ja) | 2005-09-30 | 2013-01-23 | ノボザイムス,インコーポレイティド | セルロース材料の分解又は転換を増強するための方法 |
| EP1969123B1 (en) | 2005-12-22 | 2017-07-12 | AB Enzymes Oy | Novel enzymes |
| FI120045B (fi) | 2005-12-22 | 2009-06-15 | Roal Oy | Selluloosamateriaalin käsittely ja siinä käyttökelpoiset entsyymit |
| US8304212B2 (en) | 2006-07-10 | 2012-11-06 | Dyadic International, Inc. | Methods and compositions for degradation of lignocellulosic material |
| MX2009012845A (es) * | 2007-05-31 | 2009-12-15 | Novozymes Inc | Metodos para aumentar la actividad celulosica mejorada de un polipeptido. |
| WO2008148131A1 (en) | 2007-05-31 | 2008-12-04 | Novozymes, Inc. | Polypeptides having cellulolytic enhancing activity and polynucleotides encoding same |
| WO2009042846A1 (en) | 2007-09-28 | 2009-04-02 | Novozymes A/S | Polypeptides having acetylxylan esterase activity and polynucleotides encoding same |
| CN101978049B (zh) | 2007-11-30 | 2013-11-06 | 诺维信公司 | 具有阿拉伯呋喃糖苷酶活性的多肽及其编码多核苷酸 |
| CN101932704A (zh) | 2007-12-05 | 2010-12-29 | 诺维信公司 | 具有木聚糖酶活性的多肽和编码该多肽的多核苷酸 |
| EP2224822B1 (en) | 2007-12-06 | 2014-05-21 | Novozymes A/S | Polypeptides having acetylxylan esterase activity and polynucleotides encoding same |
| BRPI0821088A2 (pt) | 2007-12-07 | 2015-09-15 | Novozymes As | construção de ácido nucléico, célula hospedeira microbiana transgênica, e, métodos para produzir um polipeptídeo tendo atividade de feruloil esterase, para produzir um mutante de uma célula-mãe, para inibir a expressão de um polipeptídeo tendo atividade de feruloil esterase em uma célula, para produzir uma proteína, e, para degradar um material contendo xilano. |
| WO2009085864A2 (en) * | 2007-12-19 | 2009-07-09 | Novozymes A/S | Polypeptides having cellulolytic enhancing activity and polynucleotides encoding same |
-
2010
- 2010-01-28 CN CN2010800153740A patent/CN102388134A/zh active Pending
- 2010-01-28 EP EP10702001A patent/EP2391715A1/en not_active Withdrawn
- 2010-01-28 US US13/144,810 patent/US8604277B2/en not_active Expired - Fee Related
- 2010-01-28 WO PCT/US2010/022405 patent/WO2010088387A1/en not_active Ceased
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN104822878A (zh) * | 2012-12-07 | 2015-08-05 | 诺维信公司 | 改进纸浆的排水 |
| CN104822878B (zh) * | 2012-12-07 | 2017-12-22 | 诺维信公司 | 改进纸浆的排水 |
| CN107002056A (zh) * | 2014-09-05 | 2017-08-01 | 诺维信公司 | 碳水化合物结合模块变体以及编码它们的多核苷酸 |
Also Published As
| Publication number | Publication date |
|---|---|
| WO2010088387A1 (en) | 2010-08-05 |
| EP2391715A1 (en) | 2011-12-07 |
| US8604277B2 (en) | 2013-12-10 |
| US20120096597A1 (en) | 2012-04-19 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US8604277B2 (en) | Polypeptides having beta-glucosidase activity and polynucleotides encoding same | |
| CN102112604B (zh) | 具有乙酰木聚糖酯酶活性的多肽和编码该多肽的多核苷酸 | |
| CN102482652B (zh) | 具有纤维二糖水解酶活性的多肽和编码该多肽的多核苷酸 | |
| CN102482680B (zh) | 具有木聚糖酶活性的多肽和编码该多肽的多核苷酸 | |
| CN102639697B (zh) | 具有木聚糖酶活性的多肽和编码该多肽的多核苷酸 | |
| CN102597243B (zh) | 具有纤维素分解增强活性的多肽和编码该多肽的多核苷酸 | |
| US8299322B2 (en) | Polypeptides having xylanase activity and polynucleotides encoding same | |
| US8735128B2 (en) | Polypeptides having alpha-glucuronidase activity and polynucleotides encoding same | |
| US8058513B2 (en) | Polypeptides having feruloyl esterase activity and polynucleotides encoding same | |
| CN102712916A (zh) | 具有β-葡糖苷酶活性的多肽和编码该多肽的多核苷酸 | |
| CN102918151B (zh) | 纤维二糖水解酶变体及编码其的多核苷酸 | |
| CN102666847A (zh) | 具有纤维二糖水解酶活性的多肽和编码该多肽的多核苷酸 | |
| CN102300986A (zh) | 具有纤维素分解增强活性的多肽和编码该多肽的多核苷酸 | |
| US8629324B2 (en) | Polypeptides having expansin activity and polynucleotides encoding same | |
| CA2745608A1 (en) | Polypeptides having feruloyl esterase activity and polynucleotides encoding same | |
| CN102597228A (zh) | 纤维二糖水解酶变体及编码其的多核苷酸 | |
| CN102695719A (zh) | 具有纤维素分解增强活性的多肽和编码该多肽的多核苷酸 | |
| CN103068976A (zh) | 具有纤维二糖水解酶活性的多肽和编码该多肽的多核苷酸 | |
| CN104245930A (zh) | 具有葡萄糖醛酸酯酶活性的多肽以及编码其的多核苷酸 | |
| CN104245929A (zh) | 具有α-葡糖醛酸糖苷酶活性的多肽以及编码其的多核苷酸 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| C06 | Publication | ||
| PB01 | Publication | ||
| C10 | Entry into substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| C02 | Deemed withdrawal of patent application after publication (patent law 2001) | ||
| WD01 | Invention patent application deemed withdrawn after publication |
Application publication date: 20120321 |