

Lobo dell'insula
| Lobo dell'insula | |
|---|---|
 alla rimozione degli operculi. | |
 rostrale al ponte. (L'insula è etichettata nella parte superiore destra.) | |
| Anatomia del Gray | (EN) Pagina 825 |
| Sistema | Sistema nervoso centrale |
| Arteria | Cerebrale media |
| Identificatori | |
| MeSH | A08.186.211.200.885.287.500.364 |
| TA | A14.1.09.149 e A12.2.07.053 |
| FMA | 67329 |
| NeuroNames | hier-93 |
| ID NeuroLex | birnlex_1117 |
In ogni emisfero del cervello dei mammiferi si trova una parte denominata il lobo dell'insula (spesso chiamata corteccia insulare o semplicemente insula), e nell'Homo sapiens è una porzione della corteccia cerebrale che si trova profondamente all'interno della scissura di Silvio; la si può osservare divaricando le due labbra della scissura di Silvio. L'area corticale che lo copre e lo separa dalla superficie laterale del cervello sono gli opercoli (che in latino significa "palpebre"). Gli opercoli sono formati da parti dei lobi frontale, temporale e parietale che costituiscono un coperchio per l'insula.
La corteccia insulare viene divisa in due parti: una anteriore (insula anteriore), costituita da circa tre circonvoluzioni brevi e una posteriore (insula posteriore), costituita da una o due circonvoluzioni lunghe. La porzione posteriore dell'insula risulta essere più piccola, e vi sono state identificate più di dodici campi citoarchitettonici
I limiti del lobo dell'insula, nei confronti degli altri lobi, sono rappresentati da tre solchi: una scissura circolare anteriore, posta al davanti della prima circonvoluzione breve e con decorso obliquo in alto e in avanti; una scissura circolare posteriore, posta al di dietro dell'ultima circonvoluzione lunga e con decorso obliquo in alto e all'indietro; e una scissura circolare superior', che abbraccia i margini di tutte le quattro-cinque circonvoluzioni dell'insula.
Il lobo dell'insula gioca un ruolo in diverse funzioni spesso legate all'emotività oppure alla regolazione dell'omeostasi corporea. Queste funzioni includono la percezione, il controllo motorio, l'auto-consapevolezza, le funzioni cognitive, e l'esperienza interpersonale. In relazione a queste alcuni ricercatori la sospettano coinvolta in aspetti anatomo-funzionali della psicopatologia. In letteratura sono state descritte anche delle implicazioni nella sensibilità gustativa e nelle funzioni affettrici viscerali.
L'insula venne descritta per la prima volta da Johann Christian Reil mentre scriveva sui nervi cranici, i nervi spinali e i loro plessi.[1] Questa attribuzione venne diffusa da Henry Gray nella sua raccolta di volumi di anatomia "Gray's Anatomy", da allora questa zona del cervello viene denominata Insula di Reil.[1]
Funzioni
[modifica | modifica wikitesto]Consapevolezza enterocettiva
[modifica | modifica wikitesto]La parte anteriore destra dell'insula interviene nella consapevolezza degli stati corporei fornita dagli enterocettori, come la capacità di percepire il proprio battito cardiaco. Inoltre, un maggiore volume della materia grigia insulare anteriore correla con un'aumentata accuratezza di questa sensibilità soggettiva dell'interno del corpo, e sarebbe anche in correlazione con esperienze emotive negative.[2] L'insula è coinvolta anche nel controllo della pressione arteriosa,[3] particolarmente durante e dopo l'esercizio fisico.[3] L'area del lobo limbico viene attivata anche quando il cervello percepisce un maggiore sforzo fisico.[4][5]
Dolore
[modifica | modifica wikitesto]Nella corteccia insulare viene quantificata e spesso localizzata la sensazione dolorosa.[6] Inoltre, l'insula è la zona corticale dove una persona immagina il dolore quando osserva immagini di eventi dolorosi mentre pensa che possano capitare al proprio corpo.[7] I pazienti affetti da sindrome del colon irritabile hanno un'abnorme elaborazione del dolore viscerale nella corteccia insulare, in correlazione all'inibizione disfunzionale e alla percezione del dolore all'interno del cervello.[8]
Temperatura
[modifica | modifica wikitesto]Un'altra percezione della corteccia insulare anteriore destra è quella dell'entità del calore non doloroso[9] oppure del freddo non doloroso[10] delle sensazioni ricevute dalla pelle. Altre sensazioni interne elaborate dall'insula includono la distensione dello stomaco che provoca sazietà.[11][12] Anche il riempimento della vescica urinaria provoca l'attivazione della corteccia insulare.[13]
Sistema vestibolare, equilibrio
[modifica | modifica wikitesto]La corteccia cerebrale che processa sensazioni vestibolari si estende nel lobo dell'insula[14] e piccole lesioni nella corteccia insulare anteriore possono causare la perdita dell'equilibrio e causare vertigini.[15]
Altre percezioni non enterocettive includono l'ascolto passivo della musica,[16] il riso e il pianto,[17], l'empatia e la compassione,[18] e alcuni aspetti del linguaggio[19]
Controllo motorio
[modifica | modifica wikitesto]Nel controllo motorio ha un ruolo minore nei movimenti involontari delle mani e degli occhi, correlato all'apparato vestibolare[20][21] nella deglutizione,[22] nella motilità gastrica,[23] e nell'articolazione del linguaggio.[24][25] Lo si è identificato come un centro di "perfezionamento della funzione" dei nuclei automatici (simpatici e parasimpatici) che si trovano nel midollo allungato e che controllano l'aumento della frequenza cardiaca e della pressione arteriosa quando si compie uno sforzo fisico.[26] La ricerca sul linguaggio e la conversazione lo collega alla capacità di avviare le giuste pause e respiri che precedono e permettono lunghe e complesse frasi parlate.[27]
Si ritiene anche che l'insula sia coinvolta nell'apprendimento di atti motori[28] e gli si attribuisce un ruolo nel recupero motorio dall'ictus.[29]
Omeostasi
[modifica | modifica wikitesto]Nell'omeostasi, l'insula controlla le funzioni autonome attraverso una fine regolazione dei sistemi simpatico e parasimpatico.[30][31] Ha un ruolo non ben definito nella regolazione del sistema immune (forse attraverso la risposta allo stress mediata dal cortisolo).[32][33][34]
Coscienza del sé corporeo
[modifica | modifica wikitesto]Il lobo dell'insula è stato identificato come avente un ruolo nella coscienza del sé corporeo,[35][36] il senso di auto-attribuzione di un rapporto causa-effetto (agency in inglese)[37] e la proprietà del proprio corpo.[38]
Emotività sociale
[modifica | modifica wikitesto]La parte anteriore dell'insula elabora il senso di disgusto provato da una persona sia agli odori[39] ed alla vista di una qualsiasi contaminazione (escrementi, sporcizia, ecc.) e mutilazione[40] — attivandosi anche quando si immagina soltanto quest'esperienza.[41] Questo si associa con un sistema di neuroni specchio, simile ad un collegamento tra l'esperienza esterna e interna.
Nell'esperienza sociale l'insula viene coinvolta nel processamento della violazione alle norme (delitti e convenzioni sociali),[42] nel processo emozionale,[43] nell'empatia,[44] e nell'orgasmo.[45]
Emozioni
[modifica | modifica wikitesto]La corteccia insulare, in particolare la sua porzione più anteriore, viene considerata un'area associata alla corteccia limbica. L'insula viene sempre più studiata per il suo ruolo nella rappresentazione corporea e nell'esperienza emozionale soggettiva. In particolare, Antonio Damasio ha proposto che questa regione giochi un ruolo nel mappare stati viscerali che sono associati con l'esperienza emozionale, producendo così sentimenti consci. Questa in essenza è una formulazione in chiave neurobiologica delle idee di William James, che per primo propose che le esperienze soggettive emozionali (ad.es. i sentimenti) emergono da come il cervello interpreta stati corporei che vengono suscitati da eventi emotivi. Questo è un esempio di "filosofia del corpo".
Dal punto di vista funzionale, si pensa che l'insula processi informazioni convergenti per produrre un contesto emozionale rilevante in risposta all'esperienza sensoriale. Più specificamente, l'insula anteriore correla di più alle funzioni olfattive, gustative, viscerali-autonome, e alla funzione limbica, mentre l'insula posteriore correla di più alla funzione uditiva-somato-estesica-scheletrica-motoria. Alcuni esperimenti di risonanza magnetica funzionale hanno rivelato che l'insula ha un ruolo importante nel regolare l'esperienza dolorosa e l'esperienza di un certo numero di emozioni di base, includendo l'ira, la paura, l'ansia, il disgusto, la felicità e la tristezza.
Studi di imaging funzionale hanno dimostrato un coinvolgimento dell'insula nei desideri consci, come lo smodato desiderio per il cibo e la droga. Il denominatore comune di questi stati emotivi è che essi modificano l'organismo in qualche modo e sono associati a qualità soggettive molto rilevanti. L'insula si trova in una posizione molto adatta per poter integrare l'informazione relativa agli stati corporei in modo da eseguire processi cognitivi ed emozionali di elevato processo. L'insula riceve informazioni dalle vie sensoriali "afferenti omeostatiche" attraverso il talamo e manda segnali ad un certo numero di altre strutture correlate al lobo limbico, come l'amigdala, lo striato ventrale e la corteccia orbitofrontale, come anche alle cortecce motorie.[46]
Uno studio che si è avvalso della risonanza magnetica ha scoperto che l'insula anteriore destra era significativamente più spessa nelle persone che meditano.[47]
Un altro studio utilizzante la morfometria basata sui voxel e la MRI, che studiò degli esperti in meditazione nelle tecniche Vipassana, venne eseguito per approfondire le scoperte del gruppo di Lazar. Questo studio rivelò un aumento della concentrazione della materia grigia in questa e in altre aree del cervello nei meditatori esperti.[48]
Rilevanza
[modifica | modifica wikitesto]La ricerca di imaging funzionale suggerisce che l'insula sia coinvolta in due tipi di rilevanza ("salience" in inglese). L'elaborazione di informazioni enterocettive che collegano gli enterocettori (ricettori non dolorifici del sistema delle fibre sensitive dorsali: movimento degli arti, pressione, stiramento tendini, vibrazione, ecc.) con la rilevanza emozionale per generare una rappresentazione soggettiva della posizione del corpo. Questo processo coinvolge la corteccia insulare anteriore con la corteccia cingolare anteriore pre-genuale (area di Brodmann 33) e le cortecce medio-cingolate anteriore e posteriore. Secondo, un sistema generale di salience, che si occupa del monitoraggio dell'ambiente circostante, della selezione di risposte e comportamenti adeguati, e dell'orientamento muscolo-scheletrico del corpo che coinvolge tutta la corteccia insulare e la corteccia medio-cingolare.[49]
Una proposta alternativa o forse complementare è che la corteccia anteriore insulare destra regola l'interazione tra la salience dell'attenzione selettiva che viene creata dalla coscienza per espletare un compito (il sistema dorsale dell'attenzione) e la rilevanza del livello di vigilanza creato per mantenersi focalizzati sopra la parte rilevante dell'ambiente (sistema dell'attenzione ventrale).[50] Questa regolazione dell'attenzione potrebbe essere particolarmente importante durante compiti impegnativi dove progressivamente l'attenzione può andare incontro a fatica e dunque causare errori per negligenza (anche pericolosi, ad esempio nell'ambito della sicurezza sul lavoro) ma se vi è troppa vigilanza si rischia di incorrere in una diminuzione della performance per l'eccessiva ansietà.[50]
Neuroanatomia
[modifica | modifica wikitesto]Connessioni
[modifica | modifica wikitesto]L'insula anteriore riceve proiezioni dirette dalla parte basale del nucleo ventro-mediale (VMb) del talamo e un particolarmente grosso input dal nucleo centrale della amigdala. Inoltre l'insula anteriore invia proiezioni all'amigdala. L'insula posteriore si connette reciprocamente con la corteccia sensoriale secondaria (S2) e riceve afferenze dal nucleo ventro-postero-inferiore del talamo (VPI) (spinotalamicamente attivato). Alcune ricerche più recenti eseguite da Bud Craig e dai suoi colleghi mostrano che questa regione riceve afferenze dal nucleo ventro-mediale (parte posteriore) del talamo, che sono altamente specializzate per convogliare informazione emozionali/omeostatiche come il dolore, la temperatura, il prurito, la condizione locale di saturazione dell'ossigeno e il toccamento sensuale.
Citoarchitettonica
[modifica | modifica wikitesto]La corteccia insulare ha regioni di struttura cellulare variabile (citoarchitettonica), che variano da corteccia cerebrale granulare nella porzione posteriore fino ad a-granulare nella porzione anteriore. Il lobo dell'insula riceve anche afferenze differenziali corticali e talamiche per tutta la sua lunghezza.
Origine filogenetica
[modifica | modifica wikitesto]Da alcune autorità in anatomia, la corteccia insulare viene considerata un lobo separato del telencefalo.[51] Altre fonti vedono l'insula di Reil come parte del lobo temporale.[52] A volte viene raggruppato con altre strutture limbiche profonde del cervello per costituire (funzionalmente più che anatomicamente) un lobo limbico.
Classificabile come corteccia paralimbica, la corteccia insulare è dal punto di vista filogenetico una struttura molto antica. Gioca un ruolo una grande varietà di importanti funzioni altamente conservate (dall'evoluzione) che sono correlate alle necessità basiche della sopravvivenza, come il gusto, le sensazioni viscerali e il controllo nervoso-autonomo (le cosiddette funzioni omeostatiche). Esiste evidenza che in aggiunta a le funzioni più conservate, l'insula può giocare un ruolo in alcune funzioni "superiori" che si vedono soltanto nell'essere umano e in altri grandi primati. John Allman e i suoi colleghi hanno dimostrato che la corteccia insulare anteriore contiene una popolazione di neuroni, chiamati neuroni a fuso, che sono specifici dei grossi primati. Questi neuroni vengono trovati anche nella corteccia cingolata anteriore, che è un'altra regione che ha raggiunto un alto livello di specializzazione nei grossi primati.
I neuroni fusiformi ("spindle neurons") vengono trovati con maggiore densità nella corteccia insulare destra. È stato proposto che questi neuroni siano coinvolti in processi cognitivo-emozionali che sono specidici delle "grandi scimmie", come l'empatia ed i sentimenti emozionali di auto-coscienza. Questo viene supportato da studi di imaging funzionale che mostrano che la struttura e la funzione dell'insula anteriore destra sono correlate con la capacità di sentire il proprio battito cardiaco, o di provare empatia rispetto al dolore altrui. Si pensa che queste funzioni non siano distinte dalle funzioni "inferiori" dell'insula, ma che piuttosto emergano come una conseguenza del ruolo dell'insula nel convogliare informazioni omeostatiche alla coscienza.
Clinica
[modifica | modifica wikitesto]Afasia progressiva non-fluente
[modifica | modifica wikitesto]L'afasia progressiva non-fluente consiste nel deterioramento delle normali funzioni del linguaggio, che provoca la perdita dell'abilità di comunicare fluentemente, pur mantenendo la capacità di comprendere le singole parole udite e la capacità di comprendere altre percezioni non linguistiche come il tono emotivo del discorso. Si trova in un buon numero di condizioni neurologiche degenerative, che includono la Malattia di Pick, vari tipi di malattia del motoneurone, la degenerazione corticobasale, la demenza frontotemporale e la malattia di Alzheimer. Si associa spesso all'ipometabolismo dello stesso lobo[53] e all'atrofia della corteccia insulare anteriore sinistra.[54]
Comportamento additivo e di dipendenza
[modifica | modifica wikitesto]"L'insula si incarica anche di interpretare stati corporei come la fame e le voglie particolari ed aiuta a spingere le persone ad attivarsi per raggiungere il prossimo panino, sigaretta o striscia di cocaina."[55] Un certo numero di studi di risonanza magnetica funzionale ha mostrato che la corteccia insulare viene attivata quando tossicodipendenti e altre persone con vari tipi di additività vengono esposti a stimoli o indizi ambientali che scatenano le voglie. Questo è stato mostrato per una varietà di droghe di abuso, che includono la cocaina, l'alcool, gli oppiacei e la nicotina. Nonostante queste rilevanti scoperte, il lobo dell'insula è stato trascurato nella letteratura frutto di studio dell'additività alle droghe, forse perché non è conosciuto come diretto bersaglio del sistema della dopamina meso-telencefalica che attualmente si considera preponderante per spiegare le teorie della ricompensa che generano addittività. Recenti ricerche[56] hanno mostrato che i fumatori di sigaretta che subiscono un danno nella corteccia insulare, ad esempio da un ictus, mostrano la pratica eliminazione della loro dipendenza dalle sigarette. Comunque, lo studio venne condotto in media dopo circa otto anni dopo lo stroke (colpo apoplettico), e gli stessi autori ammettono che il recall bias potrebbe avere alterato i risultati[57]
Si scopri che questi individui erano 136 volte più sensibili ad una sospensione dell'addittività al fumo rispetto ai fumatori con danni in altre aree. La riduzione della dipendenza veniva messa in evidenza dai cambiamenti auto-riferiti nel comportamento come l'astensione dal fumo meno di un giorno dopo il danno cerebrale, a smettere di fumare con grande facilità, al non riprendere a fumare dopo aver smesso, e al non avere alcun bisogno di riprendere a fumare. Questo suggerisce un ruolo significativo per la corteccia insulare nei meccanismi neurologici sottostanti all'addittività alla nicotina e ad altre droghe, e potrebbe rendere quest'area del cervello un bersaglio possibile per nuovi farmaci contro l'abuso di sostanze. Nelle dipendenze da sostanze, queste scoperte suggeriscono che le funzioni mediate dall'insula, specialmente i sentimenti consci, possono essere particolarmente importanti nel mantenimento della dipendenza, anche se questo punto di vista non viene sostenuto da alcuna moderna ricerca o da riesami di tutta la letteratura riguardanti la materia.[58]
Un recente studio nei topi eseguito da Contreras et al.[59] ha corroborato queste scoperte mostrando che la inattivazione reversibile dell'insula danneggia la conditioned place preference per l'anfetamina, un modello animale di dipendenza dalla droga indotto dall'esperienza associata a simboli ("cue-induced"). In questo studio, l'inattivazione dell'insula inibiva le risposte "con malessere" ("malaise") all'iniezione di cloruro di litio, suggerendo che la rappresentazione degli statieenterocettivi negativi eseguita dall'insula giuoca un ruolo nelle dipendenze da sostanze.
Comunque, in questo stesso studio, la preferenza condizionata per un luogo si verificò immediatamente dopo l'iniezione di anfetamina, facendo pensare che fossero gli effetti enterocettivi immediati e piacevoli della somministrazione di anfetamina, piuttosto che gli effetti tardivi e avversi dell'astinenza da anfetamina che sono rappresentati all'interno dell'insula.
Un modello proposto da Naqvi et al. (vedi sopra) è che l'insula immagazzini una rappresentazione degli effetti enterocettivi dell'utilizzo di droghe (ad.es. gli effetti sensoriali sulla via aerea della nicotina, gli effetti cardiovascolari dell'anfetamina), e che questa rappresentazione sia attivata dall'esposizione a simboli e indizi che sono stati previamente associati con l'assunzione di droghe. Un numero di studi di imaging funzionale hanno mostrato che l'insula si attiva durante la somministrazione di droghe che possono indurre l'abuso. Alcuni studi di imaging funzionale hanno anche mostrato che l'insula si attiva quando i tossicodipendenti vengono esposti a simboli e indizi della droga, e che questa attività si correla con le urgenze soggettive. Negli studi da esposizione a indizi, l'attività dell'insula viene avviata anche se non vi è alcuna variazione dei livelli di droga nell'organismo.
Secondo questo studio invece che rappresentare gli effetti enterocettivi dell consumo di droga come avviene, l'insula potrebbe giocare un ruolo nella memoria per gli effetti enterocettivi piacevoli provocati dall'assunzione in passato, anticipando le sensazioni piacevoli che si avranno in futuro, o entrambi. Questa rappresentazione potrebbe dare adito a urgenze consce che sono percepite come se emergessero dall'interno del corpo. Questo potrebbe fare che il tossicodipendente senta la necessità da parte del corpo di usare una droga, e può essere questa la causa per la quale persone con lesioni nell'insula riferiscano che i loro corpi hanno "dimenticato" la necessità urgente dell'abuso di droga.
Altre condizioni cliniche
[modifica | modifica wikitesto]Alcuni ricercatori hanno proposto che la corteccia insulare abbia un ruolo nei disordini d'ansia,[60] e nella difettosa regolazione delle emozioni.[61]
Galleria d'immagini
[modifica | modifica wikitesto]-
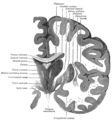 Coronal section of brain through intermediate mass of third ventricle.
Coronal section of brain through intermediate mass of third ventricle. -
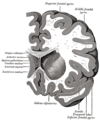 Coronal section through anterior cornua of lateral ventricles.
Coronal section through anterior cornua of lateral ventricles. -
 Coronal section of brain through anterior commissure.
Coronal section of brain through anterior commissure. -
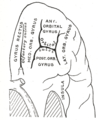 Orbital surface of left frontal lobe.
Orbital surface of left frontal lobe. -
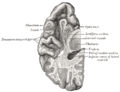 Section of brain showing upper surface of temporal lobe.
Section of brain showing upper surface of temporal lobe. -
 Horizontal section of left cerebral hemisphere.
Horizontal section of left cerebral hemisphere. -
 Human brain frontal (coronal) section
Human brain frontal (coronal) section -
 Human brain view on transverse temporal and insular gyri
Human brain view on transverse temporal and insular gyri








Note
[modifica | modifica wikitesto]- ^ a b Binder DK, Schaller K, Clusmann H, The seminal contributions of Johann-Christian Reil to anatomy, physiology, and psychiatry, in Neurosurgery, vol. 61, n. 5, novembre 2007, pp. 1091–6; discussion 1096, DOI:10.1227/01.neu.0000303205.15489.23, PMID 18091285.
- ^ Critchley HD, Wiens S, Rotshtein P, Ohman A, Dolan RJ, Neural systems supporting interoceptive awareness, in Nat. Neurosci., vol. 7, n. 2, febbraio 2004, pp. 189–95, DOI:10.1038/nn1176, PMID 14730305.
- ^ a b Lamb K, Gallagher K, McColl R, Mathews D, Querry R, Williamson JW, Exercise-induced decrease in insular cortex rCBF during postexercise hypotension, in Med Sci Sports Exerc, vol. 39, n. 4, aprile 2007, pp. 672–9, DOI:10.1249/mss.0b013e31802f04e0, PMID 17414805. URL consultato l'8 settembre 2010 (archiviato dall'url originale l'11 maggio 2012).
- ^ Williamson JW, McColl R, Mathews D, Mitchell JH, Raven PB, Morgan WP, Hypnotic manipulation of effort sense during dynamic exercise: cardiovascular responses and brain activation, in J. Appl. Physiol., vol. 90, n. 4, aprile 2001, pp. 1392–9, PMID 11247939. URL consultato l'8 settembre 2010 (archiviato dall'url originale l'8 ottobre 2009).
- ^ Williamson JW, McColl R, Mathews D, Ginsburg M, Mitchell JH, Activation of the insular cortex is affected by the intensity of exercise, in J. Appl. Physiol., vol. 87, n. 3, settembre 1999, pp. 1213–9, PMID 10484598. URL consultato l'8 settembre 2010 (archiviato dall'url originale l'8 ottobre 2009).
- ^ Baliki MN, Geha PY, Apkarian AV, Parsing pain perception between nociceptive representation and magnitude estimation [collegamento interrotto], in J. Neurophysiol., vol. 101, n. 2, febbraio 2009, pp. 875–87, DOI:10.1152/jn.91100.2008, PMID 19073802.
- ^ Ogino Y, Nemoto H, Inui K, Saito S, Kakigi R, Goto F, Inner experience of pain: imagination of pain while viewing images showing painful events forms subjective pain representation in human brain, in Cereb. Cortex, vol. 17, n. 5, maggio 2007, pp. 1139–46, DOI:10.1093/cercor/bhl023, PMID 16855007.
- ^ Song GH, Venkatraman V, Ho KY, Chee MW, Yeoh KG, Wilder-Smith CH, Cortical effects of anticipation and endogenous modulation of visceral pain assessed by functional brain MRI in irritable bowel syndrome patients and healthy controls, in Pain, vol. 126, n. 1-3, dicembre 2006, pp. 79–90, DOI:10.1016/j.pain.2006.06.017, PMID 16846694.
- ^ Olausson H, Charron J, Marchand S, Villemure C, Strigo IA, Bushnell MC, Feelings of warmth correlate with neural activity in right anterior insular cortex, in Neurosci. Lett., vol. 389, n. 1, novembre 2005, pp. 1–5, DOI:10.1016/j.neulet.2005.06.065, PMID 16051437.
- ^ Craig AD, Chen K, Bandy D, Reiman EM, Thermosensory activation of insular cortex, in Nat. Neurosci., vol. 3, n. 2, febbraio 2000, pp. 184–90, DOI:10.1038/72131, PMID 10649575.
- ^ Ladabaum U, Minoshima S, Hasler WL, Cross D, Chey WD, Owyang C, Gastric distention correlates with activation of multiple cortical and subcortical regions, in Gastroenterology, vol. 120, n. 2, febbraio 2001, pp. 369–76, DOI:10.1053/gast.2001.21201, PMID 11159877.
- ^ Hamaguchi T, Kano M, Rikimaru H, et al., Brain activity during distention of the descending colon in humans [collegamento interrotto], in Neurogastroenterol. Motil., vol. 16, n. 3, giugno 2004, pp. 299–309, DOI:10.1111/j.1365-2982.2004.00498.x, PMID 15198652.
- ^ Matsuura S, Kakizaki H, Mitsui T, Shiga T, Tamaki N, Koyanagi T, Human brain region response to distention or cold stimulation of the bladder: a positron emission tomography study, in J. Urol., vol. 168, n. 5, novembre 2002, pp. 2035–9, PMID 12394703.
- ^ Kikuchi M, Naito Y et al., Cortical activation during optokinetic stimulation — an fMRI study, in Acta Otolaryngol., vol. 129, n. 4, aprile 2009, pp. 440–3, DOI:10.1080/00016480802610226, PMID 19116795.
- ^ Papathanasiou ES, Papacostas SS, Charalambous M, Eracleous E, Thodi C, Pantzaris M, Vertigo and imbalance caused by a small lesion in the anterior insula, in Electromyogr Clin Neurophysiol, vol. 46, n. 3, 2006, pp. 185–92, PMID 16918202.
- ^ Brown S, Martinez MJ, Parsons LM, Passive music listening spontaneously engages limbic and paralimbic systems, in Neuroreport, vol. 15, n. 13, settembre 2004, pp. 2033–7, DOI:10.1097/00001756-200409150-00008, PMID 15486477. URL consultato l'8 settembre 2010 (archiviato dall'url originale l'11 maggio 2012).
- ^ Sander K, Scheich H, Left auditory cortex and amygdala, but right insula dominance for human laughing and crying, in J Cogn Neurosci, vol. 17, n. 10, ottobre 2005, pp. 1519–31, DOI:10.1162/089892905774597227, PMID 16269094.
- ^ Zimbardo begins Heroic Imagination Project | The Center for Compassion and Altruism Research and Education Archiviato il 14 luglio 2010 in Internet Archive.
- ^ Bamiou DE, Musiek FE, Luxon LM, The insula (Island of Reil) and its role in auditory processing. Literature review, in Brain Res. Brain Res. Rev., vol. 42, n. 2, maggio 2003, pp. 143–54, DOI:10.1016/S0165-0173(03)00172-3, PMID 12738055.
- ^ Anderson TJ, Jenkins IH, Brooks DJ, Hawken MB, Frackowiak RS, Kennard C, Cortical control of saccades and fixation in man. A PET study, in Brain, vol. 117, Pt 5, ottobre 1994, pp. 1073–84, DOI:10.1093/brain/117.5.1073, PMID 7953589.
- ^ Fink GR, Frackowiak RS, Pietrzyk U, Passingham RE, Multiple nonprimary motor areas in the human cortex, in J. Neurophysiol., vol. 77, n. 4, aprile 1997, pp. 2164–74, PMID 9114263. URL consultato l'8 settembre 2010 (archiviato dall'url originale l'8 ottobre 2009).
- ^ Sörös P, Inamoto Y, Martin RE, Functional brain imaging of swallowing: an activation likelihood estimation meta-analysis, in Hum Brain Mapp, vol. 30, n. 8, agosto 2009, pp. 2426–39, PMID 19107749.
- ^ Penfield W, Faulk ME, The insula; further observations on its function, in Brain, vol. 78, n. 4, 1955, pp. 445–70, PMID 13293263.
- ^ Dronkers NF, A new brain region for coordinating speech articulation, in Nature, vol. 384, n. 6605, novembre 1996, pp. 159–61, DOI:10.1038/384159a0, PMID 8906789.
- ^ Ackermann H, Riecker A, The contribution of the insula to motor aspects of speech production: a review and a hypothesis, in Brain Lang, vol. 89, n. 2, maggio 2004, pp. 320–8, DOI:10.1016/S0093-934X(03)00347-X, PMID 15068914.
- ^ Nowak M, Holm S, Biering-Sørensen F, Secher NH, Friberg L, "Central command" and insular activation during attempted foot lifting in paraplegic humans, in Hum Brain Mapp, vol. 25, n. 2, giugno 2005, pp. 259–65, DOI:10.1002/hbm.20097, PMID 15849712.
- ^ Borovsky A, Saygin AP, Bates E, Dronkers N, Lesion correlates of conversational speech production deficits, in Neuropsychologia, vol. 45, n. 11, giugno 2007, pp. 2525–33, DOI:10.1016/j.neuropsychologia.2007.03.023, PMID 17499317.
- ^ Mutschler I, Schulze-Bonhage A, Glauche V, Demandt E, Speck O, Ball T, A rapid sound-action association effect in human insular cortex, in PLoS ONE, vol. 2, n. 2, 2007, pp. e259, DOI:10.1371/journal.pone.0000259, PMC 1800344, PMID 17327919.
- ^ Weiller C, Ramsay SC, Wise RJ, Friston KJ, Frackowiak RS, Individual patterns of functional reorganization in the human cerebral cortex after capsular infarction, in Ann. Neurol., vol. 33, n. 2, febbraio 1993, pp. 181–9, DOI:10.1002/ana.410330208, PMID 8434880.
- ^ Oppenheimer SM, Gelb A, Girvin JP, Hachinski VC, Cardiovascular effects of human insular cortex stimulation, in Neurology, vol. 42, n. 9, settembre 1992, pp. 1727–32, PMID 1513461.
- ^ Critchley HD, Neural mechanisms of autonomic, affective, and cognitive integration, in J. Comp. Neurol., vol. 493, n. 1, dicembre 2005, pp. 154–66, DOI:10.1002/cne.20749, PMID 16254997.
- ^ Pacheco-López G, Niemi MB, Kou W, Härting M, Fandrey J, Schedlowski M, Neural substrates for behaviorally conditioned immunosuppression in the rat, in J. Neurosci., vol. 25, n. 9, marzo 2005, pp. 2330–7, DOI:10.1523/JNEUROSCI.4230-04.2005, PMID 15745959.
- ^ Ramírez-Amaya V, Alvarez-Borda B, Ormsby CE, Martínez RD, Pérez-Montfort R, Bermúdez-Rattoni F, Insular cortex lesions impair the acquisition of conditioned immunosuppression, in Brain Behav. Immun., vol. 10, n. 2, giugno 1996, pp. 103–14, DOI:10.1006/brbi.1996.0011, PMID 8811934.
- ^ Ramírez-Amaya V, Bermúdez-Rattoni F, Conditioned enhancement of antibody production is disrupted by insular cortex and amygdala but not hippocampal lesions, in Brain Behav. Immun., vol. 13, n. 1, marzo 1999, pp. 46–60, DOI:10.1006/brbi.1998.0547, PMID 10371677.
- ^ Karnath HO, Baier B, Nägele T, Awareness of the functioning of one's own limbs mediated by the insular cortex?, in J. Neurosci., vol. 25, n. 31, agosto 2005, pp. 7134–8, DOI:10.1523/JNEUROSCI.1590-05.2005, PMID 16079395.
- ^ Craig AD, How do you feel—now? The anterior insula and human awareness, in Nat. Rev. Neurosci., vol. 10, n. 1, gennaio 2009, pp. 59–70, DOI:10.1038/nrn2555, PMID 19096369.
- ^ Farrer C, Frith CD, Experiencing oneself vs another person as being the cause of an action: the neural correlates of the experience of agency, in Neuroimage, vol. 15, n. 3, marzo 2002, pp. 596–603, DOI:10.1006/nimg.2001.1009, PMID 11848702.
- ^ Tsakiris M, Hesse MD, Boy C, Haggard P, Fink GR, Neural signatures of body ownership: a sensory network for bodily self-consciousness, in Cereb. Cortex, vol. 17, n. 10, ottobre 2007, pp. 2235–44, DOI:10.1093/cercor/bhl131, PMID 17138596.
- ^ Wicker B, Keysers C, Plailly J, Royet JP, Gallese V, Rizzolatti G, Both of us disgusted in My insula: the common neural basis of seeing and feeling disgust, in Neuron, vol. 40, n. 3, ottobre 2003, pp. 655–64, DOI:10.1016/S0896-6273(03)00679-2, PMID 14642287.
- ^ Wright P, He G, Shapira NA, Goodman WK, Liu Y, Disgust and the insula: fMRI responses to pictures of mutilation and contamination, in Neuroreport, vol. 15, n. 15, ottobre 2004, pp. 2347–51, DOI:10.1097/00001756-200410250-00009, PMID 15640753. URL consultato l'8 settembre 2010 (archiviato dall'url originale l'11 maggio 2012).
- ^ Jabbi M, Bastiaansen J, Keysers C, A common anterior insula representation of disgust observation, experience and imagination shows divergent functional connectivity pathways, in PLoS ONE, vol. 3, n. 8, 2008, pp. e2939, DOI:10.1371/journal.pone.0002939, PMC 2491556, PMID 18698355.
- ^ Sanfey AG, Rilling JK, Aronson JA, Nystrom LE, Cohen JD, The neural basis of economic decision-making in the Ultimatum Game, in Science, vol. 300, n. 5626, giugno 2003, pp. 1755–8, DOI:10.1126/science.1082976, PMID 12805551.
- ^ Phan KL, Wager T, Taylor SF, Liberzon I, Functional neuroanatomy of emotion: a meta-analysis of emotion activation studies in PET and fMRI, in Neuroimage, vol. 16, n. 2, giugno 2002, pp. 331–48, DOI:10.1006/nimg.2002.1087, PMID 12030820.
- ^ Singer T, The neuronal basis and ontogeny of empathy and mind reading: review of literature and implications for future research, in Neurosci Biobehav Rev, vol. 30, n. 6, 2006, pp. 855–63, DOI:10.1016/j.neubiorev.2006.06.011, PMID 16904182.
- ^ Ortigue S, Grafton ST, Bianchi-Demicheli F, Correlation between insula activation and self-reported quality of orgasm in women, in Neuroimage, vol. 37, n. 2, agosto 2007, pp. 551–60, DOI:10.1016/j.neuroimage.2007.05.026, PMID 17601749.
- ^ A. D. (Bud) Craig, A new view of pain as a homeostatic emotion, in Trends in Neuroscience, vol. 26, n. 6, 2002, pp. 303–307.
- ^ Sara W. Lazar, Catherine E. Kerr, Rachel H. Wasserman, Jeremy R. Gray, Douglas N. Greve, Michael T. Treadway, Metta McGarvey, Brian T. Quinn, Jeffery A. Dusek, Herbert Benson, Scott L. Rauch, Christopher I. Moore, and Bruce Fischl, Meditation experience is associated with increased cortical thickness, in NeuroReport, vol. 16, n. 17, 2005, pp. 1893–7, DOI:10.1097/01.wnr.0000186598.66243.19, PMC 1361002, PMID 16272874.
- ^ Investigation of mindfulness meditation practitioners with voxel-based morphometry
- ^ Taylor KS, Seminowicz DA, Davis KD, Two systems of resting state connectivity between the insula and cingulate cortex, in Hum Brain Mapp, vol. 30, n. 9, settembre 2009, pp. 2731–45, DOI:10.1002/hbm.20705, PMID 19072897.
- ^ a b Eckert MA, Menon V, Walczak A, Ahlstrom J, Denslow S, Horwitz A, Dubno JR. (2009). At the heart of the ventral attention system: the right anterior insula. Hum Brain Mapp. 30(8):2530-41. DOI: 10.1002/hbm.20688 PMID 19072895
- ^ Brain Archiviato il 28 ottobre 2009 in Internet Archive., MSN Encarta. Archived Archiviato il 22 ottobre 2021 in Internet Archive. 2009-10-31.
- ^ Kolb, Bryan; Whishaw, Ian Q., Fundamentals of human neuropsychology, 5th, [New York], Worth, 2003, ISBN 0-7167-5300-6.
- ^ Nestor PJ, Graham NL, Fryer TD, Williams GB, Patterson K, Hodges JR, Progressive non-fluent aphasia is associated with hypometabolism centred on the left anterior insula, in Brain, vol. 126, Pt 11, novembre 2003, pp. 2406–18, DOI:10.1093/brain/awg240, PMID 12902311.
- ^ Gorno-Tempini ML, Dronkers NF, Rankin KP, et al., Cognition and anatomy in three variants of primary progressive aphasia, in Ann. Neurol., vol. 55, n. 3, marzo 2004, pp. 335–46, DOI:10.1002/ana.10825, PMC 2362399, PMID 14991811.
- ^ Sandra Blakeslee, A Small Part of the Brain, and Its Profound Effects, The New York Times, 6 febbraio 2007.
- ^ Nasir H. Naqvi, David Rudrauf, Hanna Damasio, Antoine Bechara., Damage to the Insula Disrupts Addiction to Cigarette Smoking (abstract), in Science, vol. 315, n. 5811, gennaio 2007, pp. 531–4, DOI:10.1126/science.1135926, PMID 17255515.
- ^ Vorel SR, Bisaga A, McKhann G, Kleber HD, Insula damage and quitting smoking, in Science, vol. 317, n. 5836, luglio 2007, pp. 318–9; author reply 318–9, DOI:10.1126/science.317.5836.318c, PMID 17641181.
- ^ Steven E. Hyman, Addiction: A Disease of Learning and Memory (abstract), in Am J Psychiatry, vol. 162, n. 8, 1º agosto 2005, pp. 1414–22, DOI:10.1176/appi.ajp.162.8.1414, PMID 16055762. URL consultato l'8 settembre 2010 (archiviato dall'url originale il 12 giugno 2011).
- ^ Marco Contreras, Francisco Ceric, Fernando Torrealba, Inactivation of the Interoceptive Insula Disrupts Drug Craving and Malaise Induced by Lithium (abstract), in Science, vol. 318, n. 5850, gennaio 2007, pp. 655–8, DOI:10.1126/science.1145590, PMID 17962567.
- ^ Paulus MP, Stein MB, An insular view of anxiety, in Biol. Psychiatry, vol. 60, n. 4, agosto 2006, pp. 383–7, DOI:10.1016/j.biopsych.2006.03.042, PMID 16780813.
- ^ Thayer JF, Lane RD, A model of neurovisceral integration in emotion regulation and dysregulation, in J Affect Disord, vol. 61, n. 3, dicembre 2000, pp. 201–16, DOI:10.1016/S0165-0327(00)00338-4, PMID 11163422.
Bibliografia
[modifica | modifica wikitesto]- Thomas P. Naidicha, et al., The Insula: Anatomic Study and MR Imaging Display at 1.5 T, in American Journal of Neuroradiology, vol. 25, n. 2, 1º febbraio 2004, pp. 222–32, PMID 14970021.
Altri progetti
[modifica | modifica wikitesto] Wikimedia Commons contiene immagini o altri file su Lobo dell'insula
Wikimedia Commons contiene immagini o altri file su Lobo dell'insula
Collegamenti esterni
[modifica | modifica wikitesto]- (EN) Edward G. Jones et al., insular cortex, in BrainMaps.org, UC Regents Davis campus.
- NIF Search - Insula via the Neuroscience Information Framework
- Kakigia R, Nakataa H, Inuia K, Hiroea N, et al., Intracerebral pain processing in a Yoga Master who claims not to feel pain during meditation, in Eur J Pain., vol. 9, n. 5, ottobre 2005, pp. 581–9, DOI:10.1016/j.ejpain.2004.12.006, PMID 16139187.«As for fMRI recording, there were remarkable changes in levels of activity in the...SII-insula (mainly the insula)»