TW202134261A - 使用il-33拮抗劑之方法 - Google Patents
使用il-33拮抗劑之方法 Download PDFInfo
- Publication number
- TW202134261A TW202134261A TW109138235A TW109138235A TW202134261A TW 202134261 A TW202134261 A TW 202134261A TW 109138235 A TW109138235 A TW 109138235A TW 109138235 A TW109138235 A TW 109138235A TW 202134261 A TW202134261 A TW 202134261A
- Authority
- TW
- Taiwan
- Prior art keywords
- antagonist
- egfr
- abnormal
- mucus
- epithelial
- Prior art date
Links
Images



































Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
- C07K16/24—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans against cytokines, lymphokines or interferons
- C07K16/244—Interleukins [IL]
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P11/00—Drugs for disorders of the respiratory system
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P11/00—Drugs for disorders of the respiratory system
- A61P11/06—Antiasthmatics
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/505—Medicinal preparations containing antigens or antibodies comprising antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/51—Complete heavy chain or Fd fragment, i.e. VH + CH1
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/515—Complete light chain, i.e. VL + CL
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/50—Immunoglobulins specific features characterized by immunoglobulin fragments
- C07K2317/56—Immunoglobulins specific features characterized by immunoglobulin fragments variable (Fv) region, i.e. VH and/or VL
- C07K2317/565—Complementarity determining region [CDR]
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2317/00—Immunoglobulins specific features
- C07K2317/70—Immunoglobulins specific features characterized by effect upon binding to a cell or to an antigen
- C07K2317/76—Antagonist effect on antigen, e.g. neutralization or inhibition of binding
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Medicinal Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- General Health & Medical Sciences (AREA)
- Pulmonology (AREA)
- Engineering & Computer Science (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Immunology (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Genetics & Genomics (AREA)
- Molecular Biology (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Peptides Or Proteins (AREA)
Abstract
本揭露關於一種用於預防或治療上皮生理學異常或EGFR介導的疾病的IL-33拮抗劑,以及相應的預防或治療方法,該等方法包括向有需要的患者投與IL-33拮抗劑。
Description
本揭露關於一種用於預防或治療上皮生理學異常或EGFR介導的疾病的IL-33拮抗劑,以及相應的預防或治療方法,該等方法包括向有需要的患者投與IL-33拮抗劑。
介白素33(IL-33),也稱為IL-1F11,係IL-1細胞介素家族之成員。IL-33係270個胺基酸的蛋白質,其由兩個結構域組成:同源結構域和細胞介素(IL-1樣)結構域。同源結構域含有核定位訊號(NLS)。已知IL-33以不同的形式存在:還原形式(redIL-33)和氧化形式(oxIL-33)。先前的研究已經顯示,還原形式在生理條件下被快速氧化以形成至少一個氧化形式之二硫鍵,並且該兩種形式可能具有不同的結合模式和作用。
先前發現,IL-33的還原形式與ST2結合,並且實際上係Th2細胞和肥大細胞表現的ST2受體的唯一已知配位基。還原型IL-33藉由結合ST2,隨後活化NFƙB和MAP激酶路徑來刺激靶細胞,從而產生細胞介素和趨化因子諸如IL-4、IL-5和IL-13來促進炎症。可溶性ST2(sST2)被認為係阻止還原型IL-33傳訊之誘餌受體。
最近發現,IL-33的氧化形式也具有生理作用。發現氧化型IL-33不結合ST2,而是與晚期糖基化終產物受體(RAGE)結合,並通過此替代路徑進行傳訊。
IL-33作為治療靶標引起了人們極大的興趣,這主要是由於現在已知之還原形式具有刺激ST2並產生強烈的炎症性作用之能力。但是,對氧化型IL-33路徑作為治療靶標的研究和興趣很少,這部分係由於其後來的發現,以及由於RAGE具有許多配位基且其下游相互作用尚未得到充分瞭解。
本文更詳細地描述了由氧化型IL-33刺激產生的該等下游RAGE相互作用中之至少一種。已經驚訝地發現,RAGE與上皮生長因子受體(EGFR)複合,作為氧化型IL-33路徑之一部分。還原型IL-33迅速轉化為氧化型IL-33,然後該氧化型IL-33與RAGE結合並與EGFR複合以刺激EGFR活性。EGFR參與的驚訝發現的重要之處在於EGFR係涉及上皮生理學異常方面的多種疾病之關鍵治療靶標。
由於此發現,據信可以與IL-33的任何形式結合的拮抗劑可以有效地阻止氧化型IL-33之傳訊。此可以是直接藉由與氧化型IL-33本身結合,或間接藉由抑制還原型IL-33向氧化型IL-33的轉化,此兩者又均阻止RAGE的刺激和EGFR的刺激。此EGFR刺激的減少將在任何EGFR介導的疾病中,但特別是在EGFR被過度刺激的病症中具有治療益處。
已知EGFR對上皮生理學具有多種穩態作用。EGFR刺激增加上皮細胞分化,增加上皮細胞遷移,並增加上皮黏膜產生。據信,抑制EGFR介導的傳訊將治療或預防存在上皮生理學異常之障礙,諸如氣道上皮組織重塑異常或黏液過度產生。
先前已將IL-33與氣道中的組織重塑相關聯(Li等人, JACI, 2014 134: 1422-32;Vannella等人, Sci Transl Med [科學轉化醫學], 337ra65;Allinne等人, JACI, 2019, 144: 1624-37)。然而,此被認為係藉由自保持擴增環(self-perpetuating amplification loop)間接發生的,藉由該自保持擴增環IL-33傳訊上調IL-33及其同源受體ST2兩者的表現,從而引起慢性ST2軸傳訊。先前尚未建立或提出IL-33本身直接影響氣道上皮生物學,因為藉由ST2產生的活性係由上面表現ST2的先天細胞,諸如巨噬細胞和2型先天淋巴樣細胞介導的。
如上所述,本揭露基於以下發現:IL-33也藉由不同的機制:RAGE-EGFR路徑直接起作用以直接影響上皮生理學。此新理解很重要,因為其可以用於擴展IL-33拮抗劑的治療應用以治療更多疾病、更多疾病症狀和更多患者。藉由靶向IL-33直接控制和抑制IL-33介導的EGFR介導的傳訊的治療機會先前尚未實現。
本申請的揭露內容首次顯示,使用IL-33拮抗劑可以藉由直接抑制RAGE/EGFR介導的oxIL-33活性來直接影響受損的上皮修復反應,減少上皮杯狀細胞的分化和增殖,減少黏液產生,並改進患有上皮生理學異常的患者(諸如患有COPD或支氣管炎的患者)的黏膜纖毛(mucociliary)運動。因此,本文提出的研究支持IL-33拮抗劑在直接預防或治療典型地由EGFR介導的作用引起,因此存在於EGFR介導的疾病中的上皮生理學異常中之治療用途。
根據第一方面,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於藉由調節或抑制RAGE-EGFR介導的作用來預防或治療上皮生理學異常。
根據替代的第一方面,提供了一種預防或治療患者的上皮生理學異常之方法,該方法包括:向有需要的患者投與有效量的IL-33拮抗劑以調節或抑制RAGE-EGFR介導之作用。
根據替代的第一方面,提供了IL-33拮抗劑在製備用於預防或治療上皮生理學異常的藥物中之用途。
根據第二方面,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於預防或治療EGFR介導的疾病。
根據替代的第二方面,提供了一種預防或治療患者的EGFR介導的疾病之方法,該方法包括:向有需要的患者投與有效量的IL-33拮抗劑。
根據替代的第二方面,提供了IL-33拮抗劑在製備用於預防或治療EGFR介導的疾病的藥物中之用途。
根據第三方面,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於藉由改進上皮生理學來預防或治療疾病。
根據替代的第三方面,提供了一種藉由改進患者的上皮生理學來預防或治療呼吸疾病之方法,該方法包括:向有需要的患者投與有效量的IL-33拮抗劑。
根據替代的第三方面,提供了IL-33拮抗劑在製備用於藉由改進上皮生理學來預防或治療呼吸疾病的藥物中之用途。
根據第四方面,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於藉由抑制EGFR介導的作用來預防或治療疾病。
根據替代的第四方面,提供了一種藉由抑制患者的EGFR介導的作用來預防或治療呼吸疾病之方法,該方法包括:向有需要的患者投與有效量的IL-33拮抗劑。
根據替代的第四方面,提供了IL-33拮抗劑在製備用於藉由抑制EGFR介導的作用來預防或治療呼吸疾病的藥物中之用途。
根據另一方面,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於藉由抑制IL-33介導的EGFR傳訊來預防或治療疾病。
根據替代的另一方面,提供了一種藉由抑制患者的IL-33介導的EGFR傳訊來預防或治療疾病之方法,該方法包括:向有需要的患者投與有效量的IL-33拮抗劑。
根據替代的另一方面,提供了IL-33拮抗劑在製備用於藉由抑制IL-33介導的EGFR傳訊來預防或治療疾病的藥物中之用途。
上文定義的方面的其他特徵和實施方式在下文的帶標題部分中進行描述。每個部分可以以任何可相容組合與任何上述方面組合。
定義
如本文所用的「IL-33」蛋白係指介白素33,特別是哺乳動物介白素33蛋白,例如以UniProt編號095760保藏的人蛋白。然而,顯然此實體不為單一種類,而是以還原和氧化形式存在。鑒於還原形式在體內例如在5分鐘至40分鐘時段內和在體外快速氧化,先前技術提及IL-33實際上可能是提及氧化形式。此外,商業測定可能無法有效地區分還原形式與氧化形式。術語「IL-33」和「IL-33多肽」可互換使用。在某些實施方式中,IL-33係全長。在另一實施方式中,IL-33係成熟的截短IL-33(胺基酸112-270)。最近研究表明全長IL-33係有活性的(Cayrol和Girard, Proc Natl Acad Sci USA [美國國家科學院院刊] 106 (22): 9021-6 (2009);Hayakawa等人, Biochem Biophys Res Commun. [生物化學與生物物理學研究通訊] 387 (1): 218-22 (2009);Talabot-Ayer等人, J Biol Chem. [生物化學雜誌] 284 (29): 19420-6 (2009))。然而,N末端處理的或截短的IL-33(包括但不限於aa 72-270、79-270、95-270、99-270、107-270、109-270、111-270、112-270)可具有增強的活性(Lefrancais 2012, 2014)。在另一實施方式中,IL-33可以包括全長IL-33、其片段或IL-33突變體或變體多肽,其中IL-33片段或IL-33變體多肽保留活性IL-33的一些或所有功能特性。
如本文所用的「氧化型IL-33」或「oxIL-33」係指與RAGE結合,且觸發RAGE-EGFR介導的傳訊的IL-33形式。氧化型IL-33係指作為獨特帶可見的蛋白質,例如藉由非還原條件下的西方墨點法分析,特別是質量比相應還原形式小4 Da的蛋白質。特別地,它係指在獨立地選自半胱胺酸208、227、232和259的半胱胺酸之間具有一個或兩個二硫鍵的蛋白質。在一個實施方式中,氧化型IL-33顯示不與ST2結合。
如本文所用的「還原型IL-33」或「redIL-33」係指與ST2結合且觸發ST2介導的傳訊的IL-33形式。特別地,還原形式的半胱胺酸208、227、232和259不與二硫鍵鍵合。在一個實施方式中,還原型IL-33顯示不與RAGE結合。
應理解,提及「WT IL-33」或「IL-33」可以指還原形式或氧化形式,或兩者,除非從使用該等形式的上下文中清楚地看出係指該等形式之一。
如本文所用的「IL-33的抗原性不同形式」係指可以充當抗原並被抗體或其結合片段結合的IL-33的任何形式,典型地,在本揭露的上下文中,此係指氧化型IL-33、還原型IL-33和還原型IL-33/sST2複合物。
如本文所用的「ST2介導的傳訊/作用」係指如下IL-33/ST2系統,其中由ST2識別還原型IL-33促進了細胞表面上與IL-1RAcP的二聚,並且在細胞內促進了受體複合組分MyD88、TRAF6和IRAK1-4向胞內TIR結構域的募集。因此,ST2依賴性傳訊/作用可以藉由擾動IL-33與ST2的相互作用或可替代地藉由中斷與IL-1RAcP的相互作用而被中斷和減弱。
如本文所用的「RAGE-EGFR介導的傳訊/作用」係指氧化型IL-33/RAGE-EGFR系統,其中由RAGE識別氧化型IL-33促進了細胞膜內與EGFR的複合。因此,RAGE-EGFR介導的傳訊/作用可以藉由擾動氧化型IL-33與RAGE的相互作用,或藉由中斷還原型IL-33向氧化型IL-33的轉化而被中斷和減弱。
如本文所用的「減弱……的活性」係指減少或抑制相關活性或停止相關活性。通常,減弱和抑制在本文中可互換使用。
應注意,術語「一個/種(a或an)」實體係指一個/種或多個/種該實體;例如,「抗IL-33抗體」應理解為表示一種或多種抗IL-33抗體。因此,術語「一個/種(a或an)」、「一個/種或多個/種」和「至少一個/種」在本文中可互換使用。
如本文中所用,術語「治療(treat或treatment)」係指治療性處理和防治性或預防性措施,其中目標係預防或減緩(減輕)不希望的生理變化或障礙。有益的或所希望的臨床結果包括但不限於症狀緩解、疾病程度減輕、疾病狀態穩定化(即,未惡化)、疾病進展延遲或減緩、疾病狀態改善或緩和以及減輕(無論是部分減輕還是全部減輕),無論是可檢測的還是不可檢測的。「治療」還可以意指如與未接受治療時的預期存活相比,延長存活。需要治療的那些包括已患有病症或障礙的那些以及易於患上病症或障礙的那些或打算預防病症或障礙的那些。
「受試者」或「個體」或「動物」或「患者」或「哺乳動物」意指需要診斷、預後或治療的任何受試者,特別是哺乳動物受試者,除非受試者被定義為「健康受試者」的情況。哺乳動物受試者包括人;家畜;農場動物;諸如狗、貓、豚鼠、兔子、大鼠、小鼠、馬、牛、母牛等。 IL-33 拮抗劑
本揭露關於IL-33拮抗劑的醫學用途,特別是藉由抑制IL-33介導的EGFR傳訊來預防或治療疾病的醫學用途。在特定的情況下,本揭露關於IL-33拮抗劑用於預防或治療可在EGFR介導的疾病中發現的上皮生理學異常之用途。
如本文所用的「IL-33拮抗劑」係指減弱IL-33活性,例如還原型IL-33活性、氧化型IL-33活性或兩者的活性的任何試劑。適合地,IL-33拮抗劑對還原型和/或氧化型IL-33具有特異性。適合地,該減弱藉由結合還原或氧化形式的IL-33進行。適合地,在拮抗劑減弱還原型IL-33活性和氧化型IL-33活性的情況下,該減弱藉由與還原形式的IL-33結合(即,藉由與還原型IL-33結合)進行。
適合地,IL-33拮抗劑係結合分子或其片段。
本揭露的「結合分子」或「抗原結合分子」在其最廣泛意義上係指特異性結合抗原決定簇的分子。適合地,結合分子與IL-33,特別是還原型IL-33或氧化型IL-33特異性結合。
適合地,結合分子可以選自:抗體、其抗原結合片段、適體、參考抗體分子的至少一個重鏈或輕鏈CDR以及來自一個或多個參考抗體分子的至少六個CDR。
適合地,IL-33拮抗劑係抗體或其結合片段。適合地,IL-33拮抗劑係抗IL-33抗體或其結合片段。適合地,抗IL-33抗體或其結合片段與IL-33,特別是還原型IL-33或氧化型IL-33特異性結合。
如本文所用的「抗體」係指如下文更詳細論述的免疫球蛋白分子,特別是全長抗體或包含全長抗體的分子,例如DVD-Ig分子等。
「其結合片段」與「其抗原結合片段」可互換,且指抗體片段的表位/抗原結合片段,其例如包含結合區,特別是包含6個CDR,諸如重鏈可變區中的3個CDR和輕鏈可變區中的3個CDR。
適合地,抗體或其結合片段選自:天然、多株、單株、多特異性、小鼠、人、人源化、靈長類動物化或嵌合抗體或其結合片段。適合地,抗體或其結合片段可以是表位結合片段,例如Fab'和F(ab')2、Fd、Fv、單鏈Fv(scFv)、二硫鍵連接的Fv(sdFv)、包含VL或VH結構域的片段或Fab表現文庫產生的片段。適合地,抗體或其結合片段可以是微抗體、雙抗體、三抗體、四抗體或單鏈抗體。適合地,抗體或其結合片段係單株抗體。scFv分子在本領域係已知的,並且描述於例如美國專利案號5,892,019中。
本揭露的免疫球蛋白或抗體分子可以是任何類型(例如,IgG、IgE、IgM、IgD、IgA和IgY)、類別(例如,IgG1、IgG2、IgG3、IgG4、IgA1和IgA2等)或子類別的免疫球蛋白分子。
適合地,IL-33拮抗劑藉由抑制氧化型IL-33的形成適合地抑制氧化型IL-33的活性。適合地,IL-33拮抗劑抑制還原型IL-33轉化為氧化型IL-33。
適合地,IL-33拮抗劑係還原型IL-33拮抗劑。換句話說,IL-33拮抗劑減弱了還原型IL-33的活性。適合地,該減弱藉由與還原型IL-33結合來進行。適合地,藉由與還原型IL-33結合,所述拮抗劑還因阻止該還原型IL-33轉化為氧化型IL-33形式而抑制/減弱了氧化型IL-33的活性
適合地,抑制氧化型IL-33的活性下調或關閉了RAGE依賴性傳訊和/或RAGE介導的作用。適合地,該抑制下調或關閉了RAGE-EGFR依賴性傳訊和/或RAGE-EGFR介導的作用。適合地,該抑制下調或關閉了EGFR依賴性傳訊。適合地,該抑制下調或關閉了EGFR介導的作用。特別地,已經顯示與還原型IL-33結合的IL33拮抗劑可以阻止氧化型IL-33與RAGE結合,從而抑制RAGE-EGFR傳訊。
適合地,抑制氧化型IL-33的活性下調或阻止了RAGE-EGFR複合。適合地,該抑制下調或阻止了EGFR活化,適合地,RAGE介導的EGFR活化。
適合地,IL-33拮抗劑具有上述所有抑制作用。適合地,還原型IL-33拮抗劑具有上述所有抑制作用。
適合地,IL-33拮抗劑係還原型IL-33結合分子或其片段。適合地,IL-33拮抗劑係還原型IL-33抗體或其結合片段,適合地,抗還原型IL33抗體或其結合片段。
適合地,該結合分子或其片段以小於以下的結合親和力(Kd)與redIL-33結合:小於5 x 10-2
M、10-2
M、5 x 10-3
M、10-3
M、5 x 10-4
M、10-4
M、5 x 10-5
M、10-5
M、5 x 10-6
M、10-6
M、5 x 10-7
M、10-7
M、5 x 10-8
M、10-8
M、5 x 10-9
M、10-9
M、5 x 10-10
M、10-10
M、5 x 10-11
M、10-11
M、5 x 10-12
M、10-12
M、5 x 10-13
M、10-13
M、5 x 10-14
M、10-14
M、5 x 10-15
M或10-15
M。適合地,與redIL-33的結合親和力小於5 x 10-14
M(即,0.05 pM)。適合地,使用動力學排除測定(KinExA)或BIACORE™,適合地使用KinExA,使用諸如WO 2016/156440(參見例如,實例11)中所述之方案測量結合親和力,該文獻在此藉由全文引用併入。以此結合親和力與redIL-33結合的結合分子似乎與redIL-33足夠緊密地結合,以阻止結合分子/redIL-33複合物在生物學相關的時間範圍內解離。不希望被理論束縛,認為此結合強度阻止了在抗體/抗原複合物在體內降解之前釋放抗原,使得redIL-33不被釋放並且不能進行從redIL-33向oxIL-33的轉化。因此,結合分子當以此結合親和力與redIL-33結合時,可以藉由阻止oxIL-33的形成來抑制或減弱oxIL-33的活性,從而抑制RAGE傳訊。
適合地,結合分子或其片段可以以大於或等於103
M-1
sec-1
、5 X 103
M-1
sec-1
、104
M-1
sec-1
或5 X 104
M-1
sec-1
的結合速率(k(on))與redIL-33特異性結合。例如,本揭露的結合分子可以以大於或等於105
M-1
sec-1
、5 X 105
M-1
sec-1
、106
M-1
sec-1
或5 X 106
M-1
sec-1
或107
M-1
sec-1
的結合速率(k(on))與redIL-33或其片段或變體結合。適合地,k(on)速率大於或等於107
M-1
sec-1
。
適合地,結合分子或其片段可以以小於或等於5 X 10-1
sec-1
、10-1
sec-1
、5 X 10-2
sec-1
、10-2
sec-1
、5 X l0-3
sec-1
或10-3
sec-1
的解離速率(k(off))與redIL-33特異性結合。例如,可以稱本揭露的結合分子以小於或等於5 X 10-4
sec-1
、10-4
sec-1
、5 X 10-5
sec-1
、10-5
sec-1
、5 X 10-6
sec-1
、10-6
sec-1
、5 X 10-7
sec-1
或10-7
sec-1
的解離速率(k(off))與redIL-33或其片段或變體結合。適合地,k(off)速率小於或等於10-3
sec-1
。IL-33係響應於炎症刺激迅速且以高濃度釋放的警報蛋白細胞介素。釋放至胞外環境中後約5-45分鐘,redIL-33被轉化為氧化型。因此,為了阻止redIL-33轉化為oxIL-33,本文所述之結合分子可以以該等k(on)和/或k(off)速率與redIL-33結合。不希望受理論束縛,認為該等k(on)/k(off)速率確保結合分子可以在redIL-33轉化為oxIL-33之前與redIL-33快速結合,從而減少了oxIL-33的形成,從而減弱了RAGE傳訊,適合地減弱了RAGE/EGFR傳訊,從而減弱了RAGE/EGFR介導的作用。
適合地,IL-33結合分子可以競爭性地抑制IL-33與結合分子33_640087-7B的結合(如WO 2016/156440中所述)。適合地,WO 2016/156440描述了33_640087-7B以特別高的親和力與redIL-33結合並且減弱了ST-2和RAGE依賴性IL-33傳訊。因此,競爭性抑制IL-33與結合分子33_640087-7B的結合的結合分子極有可能抑制redIL-33和oxIL-33傳訊兩者,因此特別適用於本文所述之方法。
如果結合分子或其片段以在某種程度上阻斷參考抗體與給定表位結合的程度與該表位結合,則稱該結合分子或其片段競爭性抑制參考抗體與該表位的結合。競爭性抑制可藉由本領域已知的任何方法確定,例如固相測定(諸如競爭ELISA測定)、解離增強鑭系螢光免疫測定(DELFIA®
,珀金埃爾默公司(Perkin Elmer))和放射性配位基結合測定。例如,技術人員可以藉由使用體外競爭結合測定,諸如WO 2016/156440實例1中所述之HTRF測定的衍生測定來判定結合分子或其片段是否競爭與redIL-33結合,該文獻在此藉由引用併入。例如,技術人員可以用供體螢光團標記表1的重組抗體,並將多種濃度與固定濃度的受體螢光團標記的redIL-33樣本混合。隨後,可以測量每個樣本內供體與受體螢光團之間的螢光共振能量轉移,以確定結合特徵。為了闡明競爭性結合分子,技術人員可以首先將各種濃度的測試結合分子與固定濃度的表1的標記抗體混合。當將混合物與標記的IL-33一起孵育時,與僅標記抗體的陽性對照相比,FRET訊號的減少指示競爭性結合IL-33。可以稱結合分子或其片段將參考抗體與給定表位的結合競爭性抑制至少90%、至少80%、至少70%、至少60%或至少50%。
在一些實施方式中,結合分子選自以下抗IL-33抗體中的任一種:33_640087-7B(如WO 2016/156440中所述)、稱為埃托克單抗(Etokimab)的ANB020(如WO 2015/106080中所述)、9675P(如US 2014/0271658中所述)、A25-3H04(如US 2017/0283494中所述)、Ab43(如WO 2018/081075中所述)、IL33-158(如US 2018/0037644中所述)、10C12.38.H6. 87Y.581 lgG4(如WO 2016/077381中所述)或其結合片段,該等文獻中的每一個均藉由引用併入本文中。該等抗體均如表1中所提及。
適合地,IL-33拮抗劑係包含選自表1的可變重鏈結構域(VH)和可變輕鏈結構域(VL)對的互補決定區(CDR)的抗體或抗原結合片段。對1對應於WO 2016/156440中所述之33_640087-7B的VH和VL結構域序列。對2-7對應於US 2014/0271658中所述之抗體的VH和VL結構域序列。對8-12對應於US 2017/0283494中所述之抗體的VH和VL結構域序列。對13對應於WO 2015/106080中所述之ANB020的VH和VL結構域序列。對14-16對應於WO 2018/081075中所述之抗體的VH和VL結構域序列。對17對應於US 2018/0037644中所述之IL33-158的VH和VL結構域序列。對18對應於WO 2016/077381中所述之10C12.38.H6. 87Y.581 lgG4的VH和VL結構域序列。
[表1]:示例性抗IL-33抗體VH和VL對
| 對 | SEQ ID NO: | HCVR 胺基酸序列 | SEQ ID NO: | LCVR 胺基酸序列 |
| 1 | SEQ ID NO: 1 | EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSGISAIDQSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARQKFMQLWGGGLRYPFGYWGQGTMVTVSS | SEQ ID NO: 19 | SYVLTQPPSVSVSPGQTASITCSGEGMGDKYAAWYQQKPGQSPVLVIYRDTKRPSGIPERFSGSNSGNTATLTISGTQAMDEADYYCGVIQDNTGVFGGGTKLTVL |
| 2 | SEQ ID NO: 2 | EVQLVESGGGLVQPGGSLRLSCAASGFTFRSFAMSWVRQAPGKGLELVSDLRTSGGSTYYADSVKGRLTISRDNSKNTLYLQMNSLRAEDTAVYYCAKSHYSTSWFGGFDYWGQGTLVTVSS | SEQ ID NO: 20 | DIQMTQSPSSVSASVGDRVTITCRASQGFSSWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTITNLQPEDFATYYCQQANSFPLTFGGGTKVEIK |
| 3 | SEQ ID NO: 3 | QVQLQESGPGLVKPSETLSLTCTVSGGSISSYYWSWIRQPPGKGLELIGYIYYSGSTNYNPSLKSRVTISVDTSKNHFSLKLSSVTAADTAVYYCARSQYTSSWYGSFDIWGQGTMVTVSS | SEQ ID NO: 21 | DIQMTQSPSSVSASVGDRVTITCRASQGISTWLAWFQQKPGKAPKLLIYAASTLQGGVPSRFSGSGSGPEFTLTISSLQPEDFATYYCQQANSFPWTFGQGTKVEIK |
| 4 | SEQ ID NO: 4 | QVQLVQSGAEVKKPGASVKVSCKASGYTFNSYGISWVRQAPGQGLEWMGWISSHNGNSHYVQKFQGRVSMTTDTSTSTAYMELRSLRSDDTAVYYCARHSYTTSWYGGFDYWGQGTLVTVSS | SEQ ID NO: 22 | DIQMTQSPSSVSASVGDRVTITCRASQGFSSWLAWYQQKPGKAPQLLIYAASSLQSGVPSRFSGSGSGSDFTLTISSLQPEDFATYYCQQANSFPLTFGGGTKVEIK |
| 5 | SEQ ID NO: 5 | EVQLVESGGGLVQPGGSLRLSCAASGFTFSSYALTWVRQAPGKGLEWVSFISGSGGRPFYADSVKGRFTISRDNSKNMLYLQMNSLRAEDTAIYYCAKSLYTTSWYGGFDSWGQGTLVTVSS | SEQ ID NO: 23 | DIQMTQSPSSVSASVGDRVTITCRASQGVVSWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSNSFPFTLGPGTKVDIK |
| 6 | SEQ ID NO: 6 | EVQLVESGGGLVQPGGSLRLSCAASGFTFSNYAMTWVRQAPGKGLEWVSTISGSGDNTYYADSVQGRFTISRGHSKNTLYLQMNSLRAEDTAVYYCAKPTYSRSWYGAFDFWGQGTMVTVSS | SEQ ID NO: 24 | DIQMTQSPSSVSASVGDRVTITCRASQGISSWLAWYQQKPGKAPQLLIYAASRLQSGVPSRFWGSGSGTDFTLTISSLQPEDFATYYCQQANNFPFTFGPGTKVDIK |
| 7 | SEQ ID NO: 7 | EVQLVESGGNLEQPGGSLRLSCTASGFTFSRSAMNWVRRAPGKGLEWVSGISGSGGRTYYADSVKGRFTISRDNSKNTLYLQMNSLSAEDTAAYYCAKDSYTTSWYGGMDVWGHGTTVTV SS | SEQ ID NO: 25 | DIQMTQSPSSVSASVGDRVTITCRASQGIFSWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFAIYYCQQANSVPITFGQGTRLEIK |
| 8 | SEQ ID NO: 8 | EVQLLESGGGLVQPGGSLRLSCAASGFTFSDYYMNWVRQAPGKGLEWVSSISRYSSYIYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDIGGMDVWGQGTLVTVSS | SEQ ID NO: 26 | QSVLTQPPSASGTPGQRVTISCTGSSSNIGAVYDVHWYQQLPGTAPKLLIYRNNQRPSGVPDRFSGSKSGTSASLAISGLRSEDEADYYCQTYDSSRWVFGGGTKLTVL |
| 9 | SEQ ID NO: 9 | EVQLLESGGGLVQPGGSLRLSCAASGFTFSNYYMHWVRQAPGKGLEWVSSISARSRYHYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLATRHNAFDIWGQGTLVTVSS | SEQ ID NO: 27 | QSVLTQPPSASGTPGQRVTISCSGSSSNIGNNAVSWYQQLPGTAPKLLIYASNMRVIGVPDRFSGSKSGTSASLAISGLRSEDEADYYCGAWDDSQKALVFGGGTKLTVL |
| 10 | SEQ ID NO: 10 | EVQLLESGGGLVQPGGSLRLSCAASGFTFSNYYMHWVRQAPGKGLEWVSSISARSSYIYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLATRNNAFDIWGQGTLVTVSS | SEQ ID NO: 28 | QSVLTQPPSASGTPGQRVTISCSGSSSNIGRNAVNWYQQLPGTAPKLLIYASNMRVSGVPDRFSGSKSGTSASLAISGLRSEDEADYYCWAWDDSQKVGVFGGGTKLTVL |
| 11 | SEQ ID NO: 11 | EVQLLESGGGLVQPGGSLRLSCAASGFTFSRYYMHWVRQAPGKGLEWVSSISAQSSHIYYADSVEGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLATRQNAFDIWGQGTLVTVSS | SEQ ID NO: 29 | QSVLTQPPSASGTPGQRVTISCSGSSSNIGRNAVNWYQQLPGTAPKLLIYASNMRRSGVPDRFSGSKSGTSASLAISGLRSEDEADYYCSAWDDSQKVVVFGGGTKLTVL |
| 12 | SEQ ID NO: 12 | EVQLLESGGGLVQPGGSLRLSCAASGFTFSNYYMHWVRQAPGKGLEWVSSISARSSYLYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLATRHVAFDIWGQGTLVTVSS | SEQ ID NO: 30 | QSVLTQPPSASGTPGQRVTISCSGSSSNIGNNAVNWYQQLPGTAPKLLIYASNMRRPGVPDRFSGSKSGTSASLAISGLRSEDEADYYCEAWDDSQKAVVFGGGTKLTVL |
| 13 | SEQ ID NO: 13 | MRAWIFFLLCLAGRALAQVQLMQSGAEVKKPGASVKVSCKASGYTFTSYWMHWVRQAPGQGLEWMGTIYPRNSNTDYNQKFKARVTMTRDTSTSTVYMELSSLRSEDTAVYYCARPLYYYLTSPPTLFWGQGTLVTVSS | SEQ ID NO: 31 | MRAWIFFLLCLAGRALADIQLTQSPSFLSASVGDRVTITCKASQDVGTAVAWYQQKPGKAPKLLIYWASTRHTGVPSRFSGSGSGTEFTLTISSLQPEDFATYYCQQAKTYPFTFGSGTKLEIKR |
| 14 | SEQ ID NO: 14 | EVQLVETGGGLIQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARTLHGIRAAYDAFIIWGQGTLVTVSS | SEQ ID NO: 32 | EIVLTQSPGTLSLSPGERATLSCRASQSVGINLSWYQQKPGQAPRLLIYGASHRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCHQYSQSPPFTFGGGTKVEIK |
| 15 | SEQ ID NO: 15 | EVQLVETGGGLIQPGGSLRLSCAASGFTFSFYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARTLHGIRAAYDAFIIWGQGTLVTVSS | SEQ ID NO: 33 | EIVLTQSPGTLSLSPGERATLSCRASQSVGINLSWYQQKPGQAPRLLIYGASHRLTGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCHQYSQPPPFTFGGGTKVEIK |
| 16 | SEQ ID NO: 16 | EVQLVETGGGLIQPGGSLRLSCAASGFTFSFYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARTIHGIRAAYDAFIIWGQGTLVTVSS | SEQ ID NO: 34 | EIVLTQSPGTLSLSPGERATLSCRASQSVGINLSWYQQKPGQAPRLLIYGASHRLTGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCHQYSQPPPFTFGGGTKVEIK |
| 17 | SEQ ID NO: 17 | EVQLVESGGGLVQPGGSLRLSCAASGFTFSSYWMYWVRQAPGKGLEWVAAITPNAGEDYYPESVKGRFTISRDNAKNSLYLQMNSLRAEDTAVYYCARGHYYYTSYSLGYWGQGTLVTVSS | SEQ ID NO: 35 | DIQMTQSPSSLSASVGDRVTITCKASQNINKHLDWYQQKPGKAPKLLIYFTNNLQTGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCFQYNQGWTFGGGTKVEIK |
| 18 | SEQ ID NO: 18 | EVQLVESGGGLVQPGGSLRLSCAASGFTFSSFSMSWVRQAPGKGLEWVATISGGKTFTDYVDSVKGRFTISRDDSKNTLYLQMNSLRAEDTAVYYCTRANYGNWFFEVWGQGTLVTVSS | SEQ ID NO: 36 | EIVLTQSPATLSLSPGERATLSCRASESVAKYGLSLLNWFQQKPGQPPRLLIFAASNRGSGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQSKEVPFTFGQGTKVEIK |
適合地,IL-33拮抗劑係包含以下的抗體或抗原結合片段:包含SEQ ID NO: 1的序列的重鏈可變區(HCVR)的互補決定區(CDR),和包含SEQ ID NO: 19的序列的輕鏈可變區(LCVR)的互補決定區(CDR)。該等CDR對應於源自33_640087-7B的CDR(如WO 2016/156440中所述),該等CDR結合還原型IL-33並抑制其轉化為氧化型IL-33。33_640087-7B在WO 2016/156440中進行了完整描述,該文獻以引用的方式併入本文中。
適合地,IL-33拮抗劑係包含以下的抗體或抗原結合片段:包含SEQ ID NO: 7的序列的重鏈可變區(HCVR)的互補決定區(CDR),和包含SEQ ID NO: 25的序列的輕鏈可變區(LCVR)的互補決定區(CDR)。該等CDR對應於源自抗體9675P的CDR。9675P在US 2014/0271658中進行了完整描述,該文獻以引用的方式併入本文中。
適合地,IL-33拮抗劑係包含以下的抗體或抗原結合片段:包含SEQ ID NO: 11的序列的重鏈可變區(HCVR)的互補決定區(CDR),和包含SEQ ID NO: 29的序列的輕鏈可變區(LCVR)的互補決定區(CDR)。該等CDR對應於源自抗體A25-3H04的CDR。A25-3H04在US 2017/0283494中進行了完整描述,該文獻以引用的方式併入本文中。
適合地,IL-33拮抗劑係包含以下的抗體或抗原結合片段:包含SEQ ID NO: 13的序列的重鏈可變區(HCVR)的互補決定區(CDR),和包含SEQ ID NO: 31的序列的輕鏈可變區(LCVR)的互補決定區(CDR)。該等CDR對應於源自抗體ANB020的CDR。ANB020在WO 2015/106080中進行了完整描述,該文獻以引用的方式併入本文中。
適合地,IL-33拮抗劑係包含以下的抗體或抗原結合片段:包含SEQ ID NO: 16的序列的重鏈可變區(HCVR)的互補決定區(CDR),和包含SEQ ID NO: 34的序列的輕鏈可變區(LCVR)的互補決定區(CDR)。該等CDR對應於源自抗體Ab43的CDR。Ab43在WO 2018/081075中進行了完整描述,該文獻以引用的方式併入本文中。
適合地,IL-33拮抗劑係包含以下的抗體或抗原結合片段:包含SEQ ID NO: 17的序列的重鏈可變區(HCVR)的互補決定區(CDR),和包含SEQ ID NO: 35的序列的輕鏈可變區(LCVR)的互補決定區(CDR)。該等CDR對應於源自抗體IL33-158的CDR。IL33-158在US 2018/0037644中進行了完整描述,該文獻以引用的方式併入本文中。
適合地,IL-33結合分子係包含以下的抗體或抗原結合片段:包含SEQ ID NO: 18的序列的重鏈可變區(HCVR)的互補決定區(CDR),和包含SEQ ID NO: 36的序列的輕鏈可變區(LCVR)的互補決定區(CDR)。該等CDR對應於源自抗體10C12.38.H6. 87Y.581 lgG4的CDR。10C12.38.H6. 87Y.581 lgG4在WO 2016/077381中進行了完整描述,該文獻以引用的方式併入本文中。
適合地,技術人員已知本領域中可用於鑒定抗體或其抗原結合片段的重鏈和輕鏈可變區內的CDR之方法。適合地,技術人員可以例如進行基於序列的注釋。CDR之間的區域通常高度保守,因此,可以使用邏輯規則來確定CDR位置。技術人員可以使用用於常規抗體的一組基於序列的規則(Pantazes和Maranas, Protein Engineering, Design and Selection [蛋白質工程化、設計和選擇], 2010),可替代地或另外,他還可以基於多序列比對來完善規則。可替代地,技術人員可以使用BLAST+的BLASTP指令將抗體序列與根據Kabat、Chothia或IMGT方法操作的公眾可獲得的數據庫進行比較,以鑒定最類似的注釋序列。該等方法中的每一種均設計了獨特的殘基編號方案,根據該編號方案對高變區殘基進行編號,然後根據某些關鍵位置確定六個CDR中的每一個的起始和結束。在例如與最類似的注釋序列比對後,可以將CDR從注釋序列外推至非注釋序列,從而鑒定CDR。適合的工具/數據庫係:例如Kabat數據庫、Kabatman、Scalinger、IMGT、Abnum。
適合地,IL-33拮抗劑係包含選自表1的可變重鏈結構域(VH)和可變輕鏈結構域(VL)對的抗體或抗原結合片段。
適合地,IL33抗體或其抗原結合片段包含SEQ ID NO: 1的序列的VH結構域和SEQ ID NO: 19的序列的VL結構域。
適合地,IL33抗體或其抗原結合片段包含SEQ ID NO: 7的序列的VH結構域和SEQ ID NO: 25的序列的VL結構域。
適合地,IL33抗體或其抗原結合片段包含SEQ ID NO: 11的序列的VH結構域和SEQ ID NO: 29的序列的VL結構域。
適合地,IL33抗體或其抗原結合片段包含SEQ ID NO: 13的序列的VH結構域和SEQ ID NO: 31的序列的VL結構域。
適合地,IL33抗體或其抗原結合片段包含SEQ ID NO: 16的序列的VH結構域和SEQ ID NO: 34的序列的VL結構域。
適合地,IL33抗體或其抗原結合片段包含SEQ ID NO: 17的序列的VH結構域和SEQ ID NO: 35的序列的VL結構域。
因此,適合地,IL-33拮抗劑係如下結合分子,該結合分子可以包含例如獨立地選自SEQ ID NO: 1、7、11、13、16、17和18的重鏈可變區中的3個CDR。
適合地,IL-33拮抗劑係包含根據SEQ ID NO: 1的重鏈可變區中的3個CDR的結合分子。
適合地,IL-33拮抗劑係如下結合分子,該結合分子可以包含獨立地選自SEQ ID NO: 19、25、29、31、34、35和36的輕鏈可變區中的3個CDR。
適合地,IL-33拮抗劑係包含根據SEQ ID NO: 19的輕鏈可變區中的3個CDR的結合分子。
因此,適合地,IL-33拮抗劑係如下結合分子,該結合分子可以包含例如獨立地選自SEQ ID NO: 1、7、11、13、16、17和18的重鏈可變區中的3個CDR,以及例如獨立地選自SEQ ID NO: 19、25、29、31、34、35和36的輕鏈可變區中的3個CDR。
因此,適合地,IL-33拮抗劑係如下結合分子,該結合分子包含根據SEQ ID NO: 1的重鏈可變區中的3個CDR,以及根據SEQ ID NO: 19的輕鏈可變區中的3個CDR。
因此,適合地,IL-33拮抗劑係可以包含可變重鏈結構域(VH)和可變輕鏈結構域(VL)的結合分子,該結合分子具有分別具有序列SEQ ID NO: 37、38和39的VH CDR 1-3,其中一個或多個VHCDR具有3個或更少的單個胺基酸取代、插入和/或缺失。
因此,適合地,IL-33拮抗劑係包含VH結構域的結合分子,該VH結構域包含分別具有SEQ ID NO: 37、SEQ ID NO: 38和SEQ ID NO: 39的VHCDR 1-3。
因此,適合地,IL-33拮抗劑係包含VH結構域的結合分子,該VH結構域包含分別由SEQ ID NO: 37、SEQ ID NO: 38和SEQ ID NO: 39組成的VHCDR 1-3。
因此,適合地,IL-33拮抗劑係可以包含可變重鏈結構域(VH)和可變輕鏈結構域(VL)的結合分子,該結合分子具有分別具有SEQ ID NO: 40、41和42的序列的VL CDR 1-3,其中一個或多個VLCDR具有3個或更少的單個胺基酸取代、插入和/或缺失。
因此,適合地,IL-33拮抗劑係包含VL結構域的結合分子,該VL結構域包含分別具有SEQ ID NO: 40、SEQ ID NO: 41和SEQ ID NO: 42的VLCDR 1-3。
因此,適合地,IL-33拮抗劑係包含VL結構域的結合分子,該VL結構域包含分別由SEQ ID NO: 40、SEQ ID NO: 41和SEQ ID NO: 42組成的VLCDR 1-3。
因此,適合地,IL-33拮抗劑係如下結合分子,該結合分子可以包含具有SEQ ID NO: 37的序列的VHCDR1、具有SEQ ID NO: 38的序列的VHCDR2、具有SEQ ID NO: 39的序列的VHCDR3、具有SEQ ID NO: 40的序列的VLCDR1、具有SEQ ID NO: 41的序列的VLCDR2和具有SEQ ID NO: 42的序列的VLCDR3。
因此,適合地,IL-33拮抗劑係包含VH和VL的抗體或其結合片段,其中VH具有與根據SEQ ID NO: 1、7、11、13、16、17和18的VH至少90%,例如91%、92%、93%、94%、95%、96%、97%、98%、99%或100%一致的胺基酸序列。
因此,適合地,IL-33拮抗劑係包含VH和VL的抗體或其結合片段,其中VH具有與根據SEQ ID NO: 1的VH至少90%,例如91%、92%、93%、94%、95%、96%、97%、98%、99%或100%一致的胺基酸序列。
因此,適合地,IL-33拮抗劑係包含VH和VL的抗體或其結合片段,其中上文揭露的VH具有如下序列,其中框架中的1、2、3或4個胺基酸缺失、用不同胺基酸插入和/或獨立地替換。
因此,適合地,IL-33拮抗劑係包含VH和VL的抗體或其結合片段,其中VL具有與根據SEQ ID NO: 19、25、29、31、34、35和36的VL至少90%,例如91%、92%、93%、94%、95%、96%、97%、98%、99%或100%一致的胺基酸序列。
因此,適合地,IL-33拮抗劑係包含VH和VL的抗體或其結合片段,其中VL具有與根據SEQ ID NO: 19的VL至少90%,例如91%、92%、93%、94%、95%、96%、97%、98%、99%或100%一致的胺基酸序列。
因此,適合地,IL-33拮抗劑係包含VH和VL的抗體或其結合片段,其中上文揭露的VL具有如下序列,其中框架中的1、2、3或4個胺基酸獨立地缺失、用不同胺基酸插入和/或替換。
因此,適合地,IL-33拮抗劑係包含VH和VL的抗體或其結合片段,其中VH具有與根據SEQ ID NO: 1、7、11、13、16、17和18的VH至少90%,例如91%、92%、93%、94%、95%、96%、97%、98%、99%或100%一致的胺基酸序列,並且VL具有與根據SEQ ID NO: 19、25、29、31、34、35和36的VL至少90%,例如91%、92%、93%、94%、95%、96%、97%、98%、99%或100%一致的胺基酸序列。
因此,適合地,IL-33拮抗劑係包含VH和VL的抗體或其結合片段,其中VH具有由SEQ ID NO: 1、7、11、13、16、17和18組成的胺基酸序列,並且VL具有由SEQ ID NO: 19、25、29、31、34、35和36組成的胺基酸序列。
因此,適合地,IL-33拮抗劑係包含VH和VL的抗體或其結合片段,其中VH具有由SEQ ID NO: 1組成的胺基酸序列,並且VL具有由SEQ ID NO: 19組成的胺基酸序列。 組成物和投與
本文所述之醫學用途和方法中的IL-33拮抗劑可以以藥物組成物的形式投與至患者。
適合地,本文中對「IL-33拮抗劑」的任何提及也可以指包含IL-33拮抗劑的藥物組成物。適合地,藥物組成物可以包含一種或多種IL-33拮抗劑。
適合地,IL-33拮抗劑可以以藥物有效量投與以用於體內治療如本文中治療方面的醫學用途和方法中所定義的上皮生理學異常、或EGFR介導的疾病、或呼吸疾病。
適合地,應將IL-33拮抗劑的「藥物有效量」或「治療有效量」理解為係指足以實現與IL-33有效結合並實現益處,例如改善如本文中醫學用途/方法中所述之疾病或病症的症狀的量。
適合地,IL-33拮抗劑或其藥物組成物可以根據前述治療方法/醫學用途以足以產生治療效果的量投與至人或其他動物。
適合地,IL-33拮抗劑或其藥物組成物可以按常規劑型投與至此種人或其他動物,該劑型藉由根據已知技術將IL-33拮抗劑與常規藥學上可接受的載體或稀釋劑組合來製備。
熟悉該項技術者應認識到,藥學上可接受的載體或稀釋劑的形式和特徵藉由與其組合的活性成分的量、投與途徑以及其他熟知變數來確定。熟悉該項技術者將進一步瞭解,包含一種或多種IL-33拮抗劑的混合物可以被證明係特別有效的。
可以與載體材料組合以產生單一劑型的IL-33拮抗劑的量將根據所治療的受試者和特定的投與方式而變化。適合地,藥物組成物可以以單次劑量、多次劑量或經確定時間段以輸注形式投與。適合地,也可以調整劑量方案以提供最佳期望反應(例如,治療性或防治性反應)。
適合地,IL-33拮抗劑經配製以便促進IL-33拮抗劑的投與並提升其穩定性。
適合地,藥物組成物經配製以包含藥學上可接受的無毒性無菌載體,諸如生理鹽水、無毒性緩衝劑、防腐劑等。
適合地,藥物組成物可以包含藥學上可接受的載體,包括例如水;離子交換劑;氧化鋁;硬脂酸鋁;卵磷脂;血清蛋白,諸如人血清白蛋白;緩衝物質,諸如磷酸鹽;甘胺酸;山梨酸;山梨酸鉀;飽和植物脂肪酸的偏甘油酯混合物;水、鹽或電解質,諸如硫酸魚精蛋白、磷酸氫二鈉、磷酸氫鉀、氯化鈉、鋅鹽;膠態二氧化矽;三矽酸鎂;聚乙烯吡咯啶酮;纖維素基物質;聚乙二醇;羧甲基纖維素鈉;聚丙烯酸酯;蠟;聚乙烯-聚氧丙烯嵌段聚合物;聚乙二醇和羊毛脂。
適合地,藥物組成物可以包括無菌水性或非水性溶液、懸浮液和乳液。非水性溶劑的實例係丙二醇、聚乙二醇、植物油諸如橄欖油和可注射有機酯諸如油酸乙酯。水性載體包括例如水、醇性/水性溶液、乳液或懸浮液,包括鹽水和緩衝介質。
適合地,藥學上可接受的載體包括但不限於0.01-0.1 M且較佳的是0.05 M磷酸鹽緩衝液或0.8%鹽水。其他常見的胃腸外載體包括磷酸鈉溶液、林格氏右旋糖(Ringer's dextrose)、右旋糖和氯化鈉、乳酸林格氏液(lactated Ringer's)或不揮發性油。靜脈內載體包括流體和營養補充劑、電解質補充劑,諸如基於林格氏右旋糖的那些等。還可以存在防腐劑以及其他添加劑,諸如像抗微生物劑、抗氧化劑、螯合劑和惰性氣體等。
適合地,用於可注射用途的藥物組成物可以包括無菌水溶液(在可溶於水時)或分散液和用於臨時製備無菌可注射溶液或分散液的無菌粉末。在此類情況下,組成物必須是無菌的並且必須具有達到容易注射的程度的流動性。在製造和存儲條件下它應當係穩定的,並且將被保存以防止微生物(諸如細菌和真菌)的污染作用。適合地,載體可以是含有以下的溶劑或分散介質,例如:水、乙醇、多元醇(例如,甘油、丙二醇和液體聚乙二醇等)及其適合的混合物。可以例如藉由使用塗層(諸如卵磷脂)、藉由在分散液的情況下維持所需顆粒大小以及藉由使用表面活性劑來維持適當的流動性。
用於本文揭露的治療方法的適合配製物描述於Remington's Pharmaceutical Sciences [雷明頓氏藥物科學](麥克出版公司(Mack Publishing Co.))第16版(1980)。
適合地,阻止微生物的作用可以藉由各種抗細菌劑和抗真菌劑,例如對羥苯甲酸酯、三氯三級丁醇、苯酚、抗壞血酸、硫柳汞等來實現。在許多情況下,在藥物組成物中適合地包括等滲劑(例如,糖)、多元醇(諸如甘露醇、山梨醇)或氯化鈉。可以藉由在組成物中包括延遲吸收的試劑(例如,單硬脂酸鋁和明膠)來實現可注射組成物的延長吸收。
適合地,無菌可注射溶液可以藉由在具有本文所列舉的成分之一或組合的適當溶劑中摻入所需量的活性化合物(例如,IL-33拮抗劑,單獨地或與其他活性劑組合),視需要隨後過濾滅菌來製備。通常,藉由將活性化合物併入無菌媒劑來製備分散液,該無菌媒劑含有基礎分散介質以及來自以上列舉的所需其他成分。在用於製備無菌可注射溶液的無菌粉末的情況下,製備方法可以是真空乾燥和冷凍乾燥,該等方法產生活性成分的粉末以及來自其以前的無菌過濾溶液的任何其他期望成分。
將IL-33拮抗劑或其藥物組成物投與至有需要的受試者之方法係熟悉該項技術者所熟知的或容易由熟悉該項技術者來確定的。
適合地,IL-33拮抗劑或其藥物組成物的投與途徑可以是例如口服、胃腸外、吸入或局部。適合地,如本文使用的術語胃腸外包括例如靜脈內、動脈內、腹膜內、肌內、皮下、經直腸或經陰道投與。
適合地,IL-33拮抗劑或其藥物組成物可以按可接受的劑型(包括例如,膠囊、片劑、水性懸浮液或溶液)口服投與。
適合地,IL-33拮抗劑或其藥物組成物可以藉由經鼻氣霧劑或吸入投與。使用苄醇或其他適合的防腐劑、提高生體可用率的吸收促進劑和/或其他常規增溶劑或分散劑,可以將此類組成物製成鹽水中的溶液。
適合地,胃腸外配製物可以是單次推注劑量、輸注或載入推注劑量,隨後是維持劑量。該等組成物可以按特定的固定或可變的間隔投與,例如每天一次,或基於「根據需要」投與。
適合地,將IL-33拮抗劑或其藥物組成物直接遞送至疾病或病症例如上皮生理學異常的部位,從而增加患病組織對治療劑的暴露。適合地,將IL-33拮抗劑或其藥物組成物直接投與至疾病或病症的部位。因此,適合地,將IL-33拮抗劑或其藥物組成物投與至上皮生理學異常、EGFR介導的疾病或呼吸疾病的部位。
在一個實施方式中,將IL-33拮抗劑或其藥物組成物投與至呼吸道。適合地藉由鼻內投與。適合地藉由鼻內吸入。適合地,IL-33拮抗劑或其藥物組成物可以在吸入器裝置中提供。適合的吸入器裝置係本領域熟知的。
在一個實施方式中,提供了一種包含IL-33拮抗劑或其藥物組成物的吸入器,該吸入器用於預防或治療如本文中所定義的病症或疾病。
因此,適合地,將IL-33拮抗劑或其藥物組成物配製成液體組成物。適合地,其呈可以氣霧化的液體組成物的形式。
在一個實施方式中,IL-33拮抗劑或其藥物組成物以氣霧劑形式提供。
適合地,可以將上文所述之用於製備本文所述之藥物組成物的組分以套組(kit)的形式包裝和出售。此種套組適合地具有標籤或包裝說明書以指示相關藥物組成物可用於治療患有或傾向於患有疾病或障礙的受試者。
適合地,將液體配製物用組分處理,填充至容器諸如安瓿、袋子、瓶子、注射器或小瓶中,並根據本領域中已知之方法密封在無菌條件下。適合地,容器可以被加壓,適合地,其可以是氣霧劑容器。該等容器可以被包括在上述套組中。適合地,套組可以進一步包含吸入器裝置。適合地,吸入器裝置包含本文所述之IL-33拮抗劑或藥物組成物,或可操作以包含上述可包含本文所述之IL-33拮抗劑或藥物組成物的容器。 上皮生理學異常
本揭露關於IL-33拮抗劑用於預防或治療上皮生理學異常中的醫學用途。
如本文所用的「上皮生理學異常」係指人體上皮功能的任何異常。人體上皮功能包括:充當保護下方組織的屏障;調節和交換組織與腔之間的化學實體;將化學物質分泌到腔中;以及感覺。該等功能中任一種的異常均可能具有破壞性的生理作用。上皮存在於身體的多種組織中,包括皮膚、呼吸道、胃腸道、生殖道、泌尿道、外分泌腺和內分泌腺,因此,上皮內的異常可能涉及多種疾病或病症。適合地,上皮係氣道上皮,並且上皮生理學異常是氣道上皮生理學異常。
如本文所用的「異常」係指與健康受試者中的功能相比所述功能的差異,典型地與健康受試者中的功能相比,所述功能增加或減少。
適合地,上皮選自:鱗狀、立方、柱狀和假複層上皮。適合地,上皮係柱狀上皮。
適合地,上皮係纖毛上皮。適合地,上皮係纖毛柱狀上皮。適合地,上皮生理學異常是纖毛柱狀上皮生理學異常。
適合地,上皮生理學異常包括上皮細胞遷移異常。適合地,上皮生理學異常可以包括上皮細胞遷移減少。適合地,上皮生理學異常可以包括上皮細胞增殖異常。適合地,上皮生理學異常可以包括上皮細胞增殖減少。
適合地,上皮細胞遷移的減少導致上皮修復劃痕的能力受損。適合地,上皮生理學異常包括劃痕修復受損。劃痕修復受損可能包括劃痕閉合受損和劃痕細胞密度降低。
適合地,上皮生理學異常的治療可以包括增加或改進上皮細胞遷移。適合地,上皮生理學異常的治療可以包括增加或改進上皮劃痕修復。適合地,上皮生理學異常的治療可以包括增加或改進劃痕閉合。適合地,上皮生理學異常的治療可以包括增加或改進劃痕細胞密度。
適合地,上皮生理學異常是異常黏膜纖毛生理學。
如本文所用的「黏膜纖毛生理學異常」係指上皮的特定黏膜纖毛作用的功能異常。上皮的黏膜纖毛作用的功能異常可能是由於對黏膜纖毛功能關鍵的纖毛柱狀細胞和/或杯狀細胞的功能異常。適合地,黏膜纖毛生理學異常是由於杯狀細胞的功能異常。
如本文所用的「黏膜纖毛」係指上皮內的纖毛柱狀細胞和杯狀柱狀細胞分泌和移動黏液的功能。上皮的黏膜纖毛作用可以包括:杯狀細胞增殖;杯狀細胞分化;黏液分泌;黏液成分調節;和/或黏液的運動或清除。
在一個實施方式中,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於預防或治療黏膜纖毛生理學異常,諸如上皮的黏膜纖毛生理學異常。
在一個實施方式中,提供了一種預防或治療患者的黏膜纖毛生理學異常,諸如上皮的黏膜纖毛生理學異常之方法,該方法包括:向有需要的患者投與有效量的IL-33拮抗劑。
黏膜纖毛生理學異常可以包括上皮的纖毛柱狀細胞或杯狀細胞的任何異常功能。適合地,黏膜纖毛生理學異常包括:黏液產生異常;杯狀細胞分化異常;杯狀細胞增殖異常;上皮厚度異常;黏液清除異常;和/或黏液組成異常。
適合地,黏液產生異常包括MUC5AC產生異常。適合地,杯狀細胞分化異常包括MUC5AC+杯狀細胞分化異常。適合地,杯狀細胞增殖異常包括MUC5AC+杯狀細胞增殖異常。適合地,上皮厚度異常包括上皮的總組織區域中MUC5AC+
杯狀細胞的量異常。
適合地,黏膜纖毛生理學異常包括:杯狀細胞數量增加;黏液產生增加;杯狀細胞分化增加;上皮厚度增加;和/或黏液清除減少。
適合地,黏液產生增加包括MUC5AC產生增加。適合地,杯狀細胞分化增加包括MUC5AC+杯狀細胞分化增加。適合地,杯狀細胞增殖增加包括MUC5AC+杯狀細胞增殖增加。適合地,上皮厚度增加包括上皮的總組織區域中MUC5AC+
杯狀細胞的量增加。
適合地,MUC5AC產生增加由MUC5AC
基因表現增加引起。適合地,黏膜纖毛生理學異常包括上皮細胞中MUC5AC
基因表現的增加。適合地,黏膜纖毛生理學異常包括上皮的杯狀細胞中MUC5AC
的表現增加。
適合地,黏膜纖毛生理學異常包括黏液組成的變化。此種變化可以包括黏液中含有的不同黏液化合物的比率增加或減少、一種或多種特定黏液化合物增加或減少或者黏液的濃度或厚度增加或減少。
黏液組成的變化可以包含不同黏蛋白的比率增加或減少,諸如黏蛋白MUC5AC和MUC5B的比率增加或減少。
黏液組成的變化可以包含黏蛋白的濃度增加或減少。適合地,黏液組成的變化包含黏蛋白5AC的濃度減少。適合地,黏液組成的變化包含具有上調的MUC5AC表現的杯狀細胞的數量減少。
黏液中含有的黏蛋白的此類變化可以如WO 2018/204598中所述進行測量和計算,該文獻藉由引用併入本文中。
適合地,黏液組成異常包括MUC5AC : MUC5B的比率增加。適合地,黏液組成異常包括黏液中含有的MUC5AC增加。適合地,黏液組成異常包括黏液厚度增加。
黏膜纖毛生理學異常可以包含以上症狀中的任何一種或多種的組合。
適合地,上皮生理學異常包括組織重塑異常,諸如上皮重塑異常。適合地,上皮生理學異常包括組織重塑增加。適合地,上皮生理學異常包括上皮重塑增加。
上皮生理學異常可以包含以上症狀中的任何一種或多種的組合。
治療或預防上皮生理學異常,或治療或預防黏膜纖毛生理學異常可以包含:
改進或增加黏膜纖毛清除;
減少或抑制黏液產生;
抑制黏液組成異常;
減少或抑制上皮重塑;和/或
減少或抑制杯狀細胞的分化和/或增殖。
適合地,減少或抑制黏液產生包含減少或抑制MUC5AC產生。因此,適合地,該治療或預防減少或抑制了MUC5AC產生。
適合地,抑制黏液組成異常可以包含恢復正常的黏液組成。適合地,此可以包含減少MUC5AC : MUC5B的比率。因此,適合地,該治療或預防減少了MUC5AC : MUC5B的比率。適合地,該預防或治療抑制或減少了黏液中的MUC5AC。適合地,該預防或治療減少了黏液的厚度。
適合地,減少或抑制杯狀細胞的分化和/或增殖包含減少或抑制MUC5AC+
杯狀細胞的分化或增殖。因此,適合地,該治療或預防減少或抑制了MUC5AC+
杯狀細胞的分化或增殖。
適合地,減少或抑制上皮重塑包含減少呼吸上皮的厚度。因此,適合地,該治療或預防減少了呼吸上皮的厚度。
適合地,減少或抑制上皮重塑包含減少了上皮的總組織區域中MUC5AC+
杯狀細胞的量。因此,適合地,該治療或預防減少或抑制了上皮的總組織區域中MUC5AC+
杯狀細胞的數量。
改進或增加黏膜纖毛清除包含改進或增加黏膜纖毛運動。因此,適合地,該治療或預防改進或增加了黏膜纖毛運動。
適合地,上皮係呼吸上皮。適合地,上皮生理學異常是呼吸上皮中的上皮生理學異常。
在一個實施方式中,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於治療呼吸疾病中的上皮生理學異常。
在一個實施方式中,提供了一種預防或治療患有呼吸疾病的患者的上皮生理學異常之方法,該方法包括:向有需要的患者投與有效量的IL-33拮抗劑。
適合的呼吸疾病如本文別處所定義。
適合地,上皮生理學異常是呼吸上皮中的黏膜纖毛生理學異常。
在一個實施方式中,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於預防或治療呼吸上皮中的黏膜纖毛生理學異常。
在一個實施方式中,提供了一種預防或治療患者的呼吸上皮的黏膜纖毛生理學異常之方法,該方法包括:向有需要的患者投與有效量的IL-33拮抗劑。
適合地,上皮生理學異常是呼吸疾病中的黏膜纖毛生理學異常。
在一個實施方式中,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於預防或治療呼吸疾病中的黏膜纖毛生理學異常。
在一個實施方式中,提供了一種預防或治療患有呼吸疾病的患者的黏膜纖毛生理學異常之方法,該方法包括:向有需要的患者投與有效量的IL-33拮抗劑。
適合地,上皮生理學異常存在於呼吸道中。適合地,上皮生理學異常是呼吸道的上皮生理學異常。適合地,上皮生理學異常是呼吸道的黏膜纖毛生理學異常。
呼吸道包括上呼吸道和下呼吸道。典型地,上呼吸道包括鼻道、鼻旁竇、咽和喉。典型地,下呼吸道包括氣管、支氣管、細支氣管、肺泡管和肺泡。
適合地,上皮生理學異常是下呼吸道,諸如支氣管的上皮生理學異常。
適合地,上皮生理學異常是下呼吸道的上皮生理學異常。適合地,上皮生理學異常是支氣管的上皮生理學異常。適合地,下呼吸道上皮生理學異常是下呼吸道的黏膜纖毛生理學異常。適合地,下呼吸道的黏膜纖毛生理學異常是支氣管的黏膜纖毛生理學異常。
在一個實施方式中,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於預防或治療下呼吸道的黏膜纖毛生理學異常。
在一個實施方式中,提供了一種預防或治療患者的下呼吸道的黏膜纖毛生理學異常之方法,該方法包括:向有需要的患者投與有效量的IL-33拮抗劑。
在一個實施方式中,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於預防或治療支氣管的黏膜纖毛生理學異常。
在一個實施方式中,提供了一種預防或治療患者的支氣管的黏膜纖毛生理學異常之方法,該方法包括:向有需要的患者投與有效量的IL-33拮抗劑。 EGFR 傳訊
本揭露基於如下發現:氧化型IL-33與RAGE結合,從而又與EGFR複合並起破壞上皮穩態的作用。使用IL-33拮抗劑可以抑制氧化型IL-33的傳訊,從而抑制RAGE的活化並抑制RAGE-EGFR複合。本文揭露的數據證明,阻止RAGE-EGFR複合物的形成阻止了IL-33介導的EGFR傳訊,並恢復了正常的上皮生理學。
適合地,IL-33拮抗劑抑制氧化型IL-33傳訊。
適合地,IL-33拮抗劑抑制氧化型IL-33與RAGE的結合。
適合地,IL-33拮抗劑抑制RAGE-EGFR複合物的形成。適合地,IL-33拮抗劑抑制氧化型IL33-RAGE-EGFR複合物的形成。
適合地,IL-33拮抗劑抑制EGFR的聚類。適合地,IL-33拮抗劑抑制EGFR在細胞膜中的聚類。適合地,IL-33拮抗劑抑制EGFR的內在化。適合地,IL-33拮抗劑抑制RAGE和EGFR在細胞膜內的共定位。適合地,IL-33拮抗劑抑制RAGE-EGFR複合物的內在化。
適合地,IL-33拮抗劑抑制EGFR的活化。適合地,IL-33拮抗劑抑制EGFR的磷酸化。
適合地,IL-33拮抗劑抑制RAGE-EGFR介導的作用。適合地,IL-33拮抗劑抑制由RAGE-EGFR複合物介導的作用。適合地,IL-33拮抗劑抑制由氧化型IL33-RAGE-EGFR複合物介導的作用。
適合地,IL-33拮抗劑抑制EGFR傳訊。適合地,IL-33拮抗劑抑制RAGE-EGFR傳訊。適合地,IL-33拮抗劑抑制氧化型IL33-RAGE-EGFR傳訊。
適合地,IL-33拮抗劑抑制氧化型IL-33與RAGE的結合,從而抑制RAGE-EGFR複合,從而抑制RAGE-EGFR介導的作用,諸如下游傳訊。
適合地,IL-33拮抗劑抑制IL-33介導的EGFR作用。適合地,IL-33拮抗劑抑制IL-33介導的EGFR傳訊。適合地,IL-33拮抗劑抑制氧化型IL-33介導的EGFR作用。適合地,IL-33拮抗劑抑制氧化型IL-33介導的EGFR傳訊。適合地,IL-33拮抗劑抑制氧化型IL-33介導的RAGE-EGFR作用。適合地,IL-33拮抗劑抑制氧化型IL-33介導的RAGE-EGFR傳訊。
適合地,RAGE-EGFR介導的作用由RAGE-EGFR複合物引起,適合地由氧化型IL-33-RAGE-EGFR複合物引起。
適合地,此類作用可以典型地包括下游傳訊,該下游傳訊在本文中可以被稱為EGFR傳訊或RAGE-EGFR傳訊。適合地,此種EGFR傳訊可以包括磷酸化和/或趨化因子釋放。
適合地,此種EGFR傳訊包括EGFR的磷酸化和隨後EGFR路徑中的組分諸如EGFR、PLC、JNK、MAPK/ERK 1/2、p38和STAT5的磷酸化。適合地,EGFR傳訊包括酪胺酸激酶諸如JNK、MAPK/ERK、p38的磷酸化。
適合地,EGFR傳訊包括趨化因子諸如IL-8的釋放增加。
因此,適合地,IL-33拮抗劑抑制EGFR介導的磷酸化和/或趨化因子釋放。
因此,適合地,IL-33拮抗劑抑制EGFR路徑中組分的磷酸化。適合地,IL-33拮抗劑抑制以下中的任一種的磷酸化:EGFR、PLC、JNK、MAPK/ERK 1/2、p38和STAT5。適合地,IL-33拮抗劑抑制EGFR介導的以下中的任一種的磷酸化:EGFR、PLC、JNK、MAPK/ERK 1/2、p38和STAT5。適合地,IL-33拮抗劑抑制酪胺酸激酶的磷酸化。適合地,IL-33拮抗劑抑制酪胺酸激酶的磷酸化,該等酪胺酸激酶選自:JNK、MAPK/ERK、p38。適合地,IL-33拮抗劑抑制EGFR介導的酪胺酸激酶的磷酸化,該等酪胺酸激酶選自:JNK、MAPK/ERK和p38。
因此,適合地,IL-33拮抗劑抑制趨化因子的釋放。適合地,IL-33拮抗劑抑制IL-8的釋放。適合地,IL-33拮抗劑抑制EGFR介導的趨化因子的釋放。適合地,IL-33拮抗劑抑制EGFR介導的IL-8的釋放。
在一個實施方式中,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於預防或治療EGFR介導的疾病。
在另一個實施方式中,IL-33拮抗劑可以用於藉由抑制EGFR介導的作用來預防或治療呼吸疾病。
此外,IL-33拮抗劑可以用於預防或治療EGFR介導的疾病中的上皮生理學異常。
在一個實施方式中,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於藉由改進上皮生理學異常來預防或治療EGFR介導的疾病。
適合地,EGFR介導的疾病係RAGE-EGFR介導的疾病。
適合地,EGFR介導的作用係RAGE-EGFR介導的作用。
適合地,EGFR介導的作用係RAGE-EGFR介導的傳訊。
適合地,IL-33拮抗劑抑制EGFR介導的作用。適合地,IL-33拮抗劑藉由抑制EGFR介導的作用來治療或預防疾病或病症。
適合地,IL-33拮抗劑抑制RAGE-EGFR介導的作用。適合地,IL-33拮抗劑藉由抑制RAGE-EGFR介導的作用來治療或預防疾病或病症。
如本文所述之「RAGE-EGFR介導的作用」係指由RAGE與EGFR在細胞膜中複合和所導致的異常EGFR活性引起的任何生理學作用。RAGE-EGFR介導的作用還可以包括和/或在本文中稱為「RAGE-EGFR傳訊」,視需要「RAGE-EGFR介導的傳訊」。此類RAGE-EGFR介導的作用典型地在上皮中可見,並以上皮生理學異常的形式存在。上皮生理學異常在上文中定義,但可以包括對以下的負面影響:屏障完整性;組織與腔之間化學實體的調節和交換;化學物質向腔中的分泌;以及感覺。
適合地,RAGE-EGFR介導的疾病和/或作用的特徵為EGFR活性異常。適合地,RAGE-EGFR介導的疾病和/或作用的特徵為RAGE-EGFR傳訊異常。適合地,RAGE-EGFR介導的作用和/或RAGE-EGFR傳訊係RAGE-EGFR介導的疾病的特徵。
適合地,RAGE-EGFR介導的疾病可以是特徵為上皮生理學異常的疾病。
適合地,RAGE-EGFR介導的疾病可以是特徵為呼吸上皮中的上皮生理學異常的疾病。
適合地,RAGE-EGFR介導的疾病可以是特徵為黏膜纖毛生理學異常的疾病。
適合地,RAGE-EGFR介導的疾病可以是特徵為呼吸上皮中的黏膜纖毛生理學異常的疾病。
適合的RAGE-EGFR介導的疾病可以選自下文定義的呼吸疾病中的任一種。 呼吸疾病
本揭露關於IL-33拮抗劑用於藉由以下來預防或治療呼吸疾病的醫學用途:改進上皮生理學或調節EGFR介導的作用,適合地,抑制EGFR介導的作用,適合地,抑制RAGE/EGFR介導的作用。
適合地,上皮生理學異常可能是呼吸疾病的症狀。因此,適合地,IL-33拮抗劑可以用於治療或預防特徵為上皮生理學異常的呼吸疾病。
如在另一個實施方式中所定義,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於藉由改進上皮生理學來預防或治療呼吸疾病。
上皮生理學異常如本文別處所定義。
適合地,改進上皮生理學可以包含改進上皮生理學異常。
上文描述了改進上皮生理學異常的適合手段。
適合地,藉由改進上皮生理學異常來治療呼吸疾病可以包含:
改進或增加黏膜纖毛清除;
減少或抑制黏液產生;
抑制黏液組成異常;
減少或抑制上皮重塑異常;和/或
減少或抑制杯狀細胞的分化或增殖。
上文結合上皮生理學異常的治療或預防提供了關於該等作用中的每一個的其他細節,並且在此可以與呼吸疾病的治療組合。
適合地,EGFR活性異常可能是呼吸疾病的症狀。因此,適合地,IL-33拮抗劑可以用於治療或預防特徵為EGFR活性異常的呼吸疾病。
如在另一個實施方式中所定義,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於藉由抑制EGFR介導的作用來預防或治療呼吸疾病。
EGFR介導的作用如本文別處所定義。
適合地,呼吸疾病係下呼吸疾病,適合地,呼吸疾病係影響氣管、支氣管、細支氣管、肺泡管和/或肺泡的疾病。適合地,呼吸疾病係支氣管疾病。
適合地,呼吸疾病可以選自:COPD;支氣管炎;肺氣腫;支氣管擴張,諸如CF-支氣管擴張或非CF-支氣管擴張;氣喘;氣喘與COPD重疊(ACO)。
適合地,呼吸疾病係COPD。適合地,呼吸疾病係支氣管炎性COPD。支氣管炎性COPD係COPD的一種特定形式,其中患有COPD的患者中存在慢性支氣管炎。由於肺功能下降更快,症狀增加以及繼發感染的風險增加,支氣管炎性COPD引起的死亡率高於COPD。特別地,支氣管炎性COPD患者具有較高的總黏蛋白濃度並且黏液分泌過多。因此,藉由IL-33拮抗劑抑制EGFR活性並改進黏膜纖毛生理學的發現,支氣管炎性COPD患者可以特別受益於用本文所述之IL-33拮抗劑治療。
適合地,出於相同的原因,呼吸疾病可以是支氣管炎性氣喘。
在一個實施方式中,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於預防或治療支氣管炎性COPD。
在一個實施方式中,提供了一種預防或治療患者的支氣管炎性COPD之方法,該方法包括:向有需要的患者投與有效量的IL-33拮抗劑。 ST2 傳訊
儘管本揭露關於IL-33拮抗劑抑制RAGE-EGFR介導的傳訊和作用的醫學用途,但是已經知道IL-33拮抗劑可以抑制ST2介導的傳訊和作用。因此,本文所述之醫學用途設想了EGFR介導的作用和ST2介導的作用兩者的調節。
適合地,IL-33拮抗劑用於藉由調節EGFR介導的作用和ST2介導的作用來預防和治療上皮生理學異常。適合地,IL-33拮抗劑用於藉由抑制EGFR介導的作用和ST2介導的作用來預防和治療上皮生理學異常。適合地,IL-33拮抗劑用於藉由抑制RAGE/EGFR介導的作用和ST2介導的作用來預防和治療上皮生理學異常。上皮生理學異常如本文別處所定義。
適合地,IL-33拮抗劑用於藉由調節EGFR介導的作用和ST2介導的作用來預防和治療EGFR介導的疾病。適合地,IL-33拮抗劑用於藉由抑制EGFR介導的作用和ST2介導的作用來預防和治療EGFR介導的疾病。適合地,IL-33拮抗劑用於藉由抑制RAGE/EGFR介導的作用和ST2介導的作用來預防和治療EGFR介導的疾病。EGFR介導的疾病如本文別處所定義。
適合地,ST2介導的作用包括炎症。因此,適合地,IL-33拮抗劑用於藉由調節EGFR介導的作用和ST2介導的作用來預防和治療ST2介導的疾病。適合地,IL-33拮抗劑用於藉由抑制EGFR介導的作用和ST2介導的作用來預防和治療ST2介導的疾病。適合地,IL-33拮抗劑用於藉由抑制RAGE/EGFR介導的作用和ST2介導的作用來預防和治療ST2介導的疾病。適合的ST2介導的疾病可以包括特徵為炎症的疾病。適合的ST2介導的疾病可以包括炎症性疾病。
ST2介導的疾病或炎症性疾病可以包括:COPD;過敏性障礙,諸如氣喘、慢性鼻腔鼻竇炎、食物過敏、濕疹和皮炎;纖維增生性疾病,諸如肺纖維化;肺嗜酸性粒細胞增多;胸膜惡性腫瘤;類風濕關節炎;膠原血管病;動脈粥樣硬化性血管病;蕁麻疹;炎症性腸病;克羅恩氏病(Crohn’s diseases);乳糜瀉病;全身性狼瘡;進行性全身性硬化;韋格納肉芽腫(Wegner’s granulomatosis);敗血性休克;以及白塞病(Bechet’s disease)。
適合地,ST2介導的疾病或炎症性疾病係呼吸疾病。適合地,ST2介導的疾病或炎症性疾病存在於如上文所定義的呼吸道中。
適合地,IL-33拮抗劑另外用於預防和治療炎症或炎症性疾病。適合地,IL-33拮抗劑可以另外用於預防和治療ST2介導的炎症或ST2介導的炎症性疾病。
適合地,ST2介導的疾病和EGFR介導的疾病重疊。換句話說,ST2介導的作用和EGFR介導的作用,適合地,RAGE-EGFR介導的作用,有助於疾病病理學。有利地,據信單一IL-33拮抗劑的醫學用途可以抑制由IL-33活化RAGE和ST2兩者。因此,單一IL-33拮抗劑可以同時治療RAGE-EGFR介導的疾病和ST2介導的疾病兩者。
在一個實施方式中,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於預防或治療上皮生理學異常和炎症。在一個實施方式中,提供了一種預防或治療患者的上皮生理學異常和炎症之方法,該方法包括:投與有效量的IL-33拮抗劑。
適合地,上皮生理學異常和炎症可能是呼吸疾病的症狀。因此,關於該等症狀的治療和預防的陳述可以在呼吸疾病的背景下進行,並且可以適合地包含治療或預防呼吸疾病中的上皮生理學異常和炎症。
在一個實施方式中,提供了一種IL-33拮抗劑,該IL-33拮抗劑用於預防或治療EGFR介導的疾病和ST2介導的疾病。
在一個實施方式中,提供了一種預防或治療患者的EGFR介導的疾病和ST2介導的疾病之方法,該方法包括:投與有效量的IL-33拮抗劑。
適合地,IL-33拮抗劑係還原型IL-33拮抗劑。適合地,還原型IL-33拮抗劑如上文所定義。
適合地,IL-33拮抗劑如上文所定義。適合地,IL-33拮抗劑係33_640087-7B。
可替代地,可以將不同IL-33拮抗劑以組合療法的形式使用,以抑制由IL-33活化RAGE和ST2兩者。因此,設想IL-33拮抗劑的組合同時治療RAGE-EGFR介導的疾病和ST2介導的疾病兩者。
適合地,呼吸疾病如上文所定義。適合地,呼吸疾病的特徵為EGFR活性異常和ST2活性異常。
因此,適合地,在一個實施方式中,提供了用於預防或治療上皮生理學異常的第一IL-33拮抗劑與用於預防或治療炎症的第二IL-33拮抗劑的組合。
因此,適合地,在一個實施方式中,提供了一種預防或治療患者的上皮生理學異常和炎症之方法,該方法包括:投與有效量的第一IL-33拮抗劑與有效量的第二IL-33拮抗劑的組合。
因此,適合地,在一個實施方式中,提供了用於預防或治療EGFR介導的疾病的第一IL-33拮抗劑與用於預防或治療ST2介導的疾病的第二IL-33拮抗劑的組合。
因此,適合地,在一個實施方式中,提供了一種預防或治療患者的EGFR介導的疾病和ST2介導的疾病之方法,該方法包括:投與有效量的第一IL-33拮抗劑與有效量的第二IL-33拮抗劑的組合。
適合地,第一IL-33拮抗劑用於預防或治療上皮生理學異常和/或EGFR介導的疾病。
適合地,第二IL-33拮抗劑用於預防或治療炎症和/或ST2介導的疾病。
適合地,第一和第二IL-33拮抗劑不同。
適合地,第一IL-33拮抗劑如上文所定義。適合地,第二IL-33拮抗劑可以是已知抑制ST2介導的作用的任何其他IL-33拮抗劑。適合地,第二IL-33拮抗劑也如上文所定義。
適合地,第一拮抗劑可以是還原型或氧化型IL-33拮抗劑。適合地,第二IL-33拮抗劑係還原型IL-33拮抗劑。
適合地,IL-33拮抗劑中的至少一種係33_640087-7B。適合地,第一拮抗劑係33_640087-7B。
適合地,第一和第二IL-33拮抗劑可以組合投與。適合地,第一IL-33拮抗劑和第二IL-33拮抗劑可以同時或在不同時間組合投與。適合的劑量方案可以由醫學專業人員確定。
該等陳述同樣適用於上文提及的醫學用途/治療方法,在該醫學用途/治療方法中也可以預防或治療ST-2介導的疾病。
可替代地,在其他實施方式中,可以將IL-33拮抗劑與ST2抑制劑組合投與。適合地,ST2抑制劑可以不是IL-33拮抗劑,但是可以藉由其他方式抑制ST2受體。適合地,ST2抑制劑可以起治療或預防如上文所鑒定的ST2介導的疾病的作用。
因此,在一個實施方式中,提供了一種用於治療或預防上皮生理學異常的IL-33拮抗劑與用於治療或預防炎症的ST2抑制劑的組合。
在一個實施方式中,提供了一種預防或治療患者的上皮生理學異常和炎症之方法,該方法包括:投與有效量的IL-33拮抗劑與有效量的ST2抑制劑的組合。
適合地,上皮生理學異常和炎症可能是呼吸疾病的症狀。因此,關於該等症狀的治療和預防的陳述可以在呼吸疾病的背景下進行,並且可以適合地包含治療或預防呼吸疾病中的上皮生理學異常和炎症。
因此,在一個實施方式中,提供了一種用於治療或預防EGFR介導的疾病的IL-33拮抗劑與用於治療或預防ST2介導的疾病的ST2抑制劑的組合。
在一個實施方式中,提供了一種預防或治療患者的EGFR介導的疾病與ST2介導的疾病的組合之方法,該方法包括:投與有效量的IL-33拮抗劑與有效量的ST2抑制劑的組合。
適合地,IL-33拮抗劑如本文別處所定義。適合的EGFR介導的疾病和ST2介導的疾病如本文別處所定義。
適合地,ST2抑制劑可以是本領域中已知的任何此種抑制劑,例如GSK3772847(在WO 2013/165894中描述)和RG6149(WO 2013/173761),兩個文獻均藉由引用併入本文中。
適合地,IL-33拮抗劑和ST2抑制劑可以組合投與。適合地,IL-33拮抗劑和ST2抑制劑可以同時或在不同時間組合投與。適合的劑量方案可以由醫學專業人員確定。 患者
將該等方法和醫學用途針對患者或受試者進行實踐。患者可以是需要鑒定、診斷或治療生理學病症或疾病諸如上皮生理學異常、EGFR介導的疾病或呼吸疾病的患者。
適合地,患者可以是人類。患者可以正在經歷醫學護理,或係需要醫學護理的個體。適合地,患者係男性或女性。適合地,患者係成人或兒童。
適合地,在本文所述之方法中,適合的患者可以是被認為患有上皮生理學異常,或EGFR介導的疾病或呼吸疾病的患者。例如,適合的患者可能具有與此類病症一致的症狀。
可替代地,本文所述方法的背景下的適合患者可以是被認為具有產生上皮生理學異常、或EGFR介導的疾病或呼吸疾病的風險之患者。例如,此種患者可能已經與患有此種病症的個體接觸,可能患有相關病症,或者可能滿足與所述病症相關的風險因素,如吸煙、老齡,過敏等。具體實施方式
本揭露的部分可以藉由以下實施方式來表徵,其中:
實施方式1描述了一種IL-33拮抗劑,該IL-33拮抗劑用於藉由抑制EGFR介導的作用來預防或治療疾病。
實施方式2描述了用於根據實施方式1所述使用的IL-33拮抗劑,其中該EGFR介導的作用係RAGE-EGFR介導的作用。
實施方式3描述了用於根據實施方式1或2所述使用的IL-33拮抗劑,其中該EGFR介導的作用係RAGE-EGFR介導的傳訊。
實施方式4描述了用於根據實施方式1-3中任一項所述使用的IL-33拮抗劑,其中該疾病係呼吸疾病。
實施方式5描述了用於根據實施方式1-4中任一項所述使用的IL-33拮抗劑,其中該疾病的特徵為上皮生理學異常和/或EGFR活性異常。
實施方式6描述了用於根據實施方式1-5中任一項所述使用的IL-33拮抗劑,其中該疾病選自:COPD;支氣管炎;肺氣腫;支氣管擴張,諸如CF-支氣管擴張或-CF-支氣管擴張;氣喘;或氣喘與COPD重疊(ACO)。
實施方式7描述了用於根據實施方式4-6中任一項所述使用的IL-33拮抗劑,其中該呼吸疾病係支氣管炎性COPD。
實施方式8描述了用於根據實施方式1-7中任一項所述使用的IL-33拮抗劑,其中該治療:改進黏液清除;抑制黏液產生異常;抑制上皮重塑異常;和/或抑制杯狀細胞分化異常。
實施方式9描述了用於根據任何前述實施方式所述使用的IL-33拮抗劑,其中該IL-33拮抗劑抑制氧化型IL-33的活性。
實施方式10描述了用於根據任何前述實施方式所述使用的IL-33拮抗劑,其中該IL-33拮抗劑阻止氧化型IL-33與RAGE結合,從而抑制RAGE-EGFR傳訊。
實施方式11描述了用於根據任何前述實施方式所述使用的IL-33拮抗劑,其中該IL-33拮抗劑係抗IL-33抗體或其抗原結合片段,較佳的是抗還原型IL-33抗體或抗原其結合片段。
實施方式12描述了用於根據實施方式11所述使用的IL-33拮抗劑,其中該抗IL-33抗體或其抗原結合片段包含選自表1的可變重鏈結構域(VH)和可變輕鏈結構域(VL)對的互補決定區(CDR)。
實施方式13描述了用於根據實施方式12所述使用的IL-33拮抗劑,其中該抗IL-33抗體或其抗原結合片段包含選自表1的可變重鏈結構域(VH)和可變輕鏈結構域(VL)對。
實施方式14描述了用於根據實施方式11-13中任一項所述使用的IL-33拮抗劑,其中該抗IL-33抗體或其抗原結合片段包含具有SEQ ID NO: 37的序列的VHCDR1、具有SEQ ID NO: 38的序列的VHCDR2、具有SEQ ID NO: 39的序列的VHCDR3、具有SEQ ID NO: 40的序列的VLCDR1、具有SEQ ID NO: 41的序列的VLCDR2和具有SEQ ID NO: 42的序列的VLCDR3。
實施方式15描述了用於根據實施方式11-14中任一項所述使用的IL-33拮抗劑,其中該IL-33拮抗劑係抗IL33抗體或其抗原結合片段,該抗IL33抗體或其抗原結合片段包含SEQ ID NO: 1的序列的VH結構域和SEQ ID NO: 19的序列的VL結構域。
無
將參考下圖,以舉例的方式描述實施方式,其中:
[圖 1
]:顯示了對於MAP激酶磷酸化抗體陣列上的每個檢測測定,與未處理的對照相比,激酶磷酸化的增加倍數之灰度熱圖。還原型IL-33(分別為IL-33-01和IL-33-16)沒有引起任何高於基線的訊號。oxIL-33(氧化型IL-33-01)使多種激酶的磷酸化增加;
[圖 2
]:顯示了受體酪胺酸激酶(RTK)活性陣列上每種刺激條件之訊號模式。oxIL-33,而非分別而言還原型IL33-01和IL33-16,在RTK陣列上觸發了對應於表皮生長因子受體(EGFR)之陽性訊號。點強度與受體酪胺酸激酶磷酸化有關;
[圖 3A
]:
顯示了在用增加濃度的IL-33或EGFR配位基刺激的正常人支氣管上皮(NHBE)細胞中之pEGFR(Tyr1068)活性。oxIL-33,而非還原型IL-33(IL33-01),類似於EGF、HB-EGF和TGFα促進EGFR的磷酸化;
[圖 3B
]:
顯示了在用增加濃度的IL-33或EGFR配位基刺激的A549細胞中之pEGFR(Tyr1068)活性。oxIL-33(氧化型IL-33-01),而非還原型IL-33(IL-33-01),以類似於NHBE細胞中可見的模式類似於EGF、HB-EGF和TGFα促進了EGFR之磷酸化。
[圖 3C
]:
顯示了在用增加濃度的IL-33、EGFR配位基或RAGE配位基刺激的A549細胞中之pEGFR(Tyr1068)活性。oxIL-33,而非野生型(WT)IL-33(IL-33-01)、C- > S突變的(mut)IL-33(IL-33-16)或RAGE配位基類似於EGF促進了EGFR之磷酸化;
[圖 4
]:顯示如藉由西方墨點法所分析,氧化型IL-33誘導參與EGFR路徑的多個分子(EGFR、PLC、AKT、JNK、ERK 1/2、p38)之磷酸化;
[圖 5
]:顯示藉由增加抗EGFR抗體之劑量,與同種型對照相比,減少了由oxIL-33-01誘導的STAT5磷酸化;
[圖 6
]:顯示用抗EGFR進行免疫沈澱,隨後藉由西方墨點法檢測EGFR、RAGE或IL-33。在用oxIL-33進行NHBE刺激後,IL-33和RAGE與EGFR共沈澱,這表明期形成了複合物。與EGF相比,RAGE對於oxIL-33傳訊複合物似乎係唯一的;
[圖 7A
]:
顯示oxIL-33直接與RAGE結合。HMGB1係一種已知的RAGE配位基,並且在本研究中可作為陽性對照;
[圖 7B
]:
顯示oxIL-33不直接與EGFR結合(但已知的EGFR配位基EGF直接與EGFR結合)。但是,當將RAGE與oxIL-33組合添加到此測定時,則可見EGFR結合;
[圖 8
]:顯示在指定時間點用oxIL-33活化後,在野生型和RAGE缺陷型A549細胞中,用抗EGFR或抗RAGE進行的免疫沈澱,隨後進行EGFR、RAGE和IL-33之西方墨點法;
[圖 9
]:顯示oxIL-33-01誘導的STAT5磷酸化被抗RAGE抗體而非抗ST2抗體減少;
[圖 10
]:顯示EGF和氧化型IL33(oxIL33)誘導EGFR-GFP A549細胞中之EGFR聚集和內在化。刺激5分鐘後顯示代表性圖像。長條圖顯示在用EGF和oxIL-33處理的細胞中非聚類區中EGFR的消耗(長條圖鐘形峰左移),以及該等細胞中由於聚類而引起的飽和像素數量增加(強度255)。
[圖 11
]:顯示用單獨的培養基(未刺激的對照)、30 ng/ml IL-33-01、30 ng/ml IL-33-16、30 ng/mL氧化型IL-33或30 ng/mL EGF刺激24 h後,NHBE和DHBE的IL-8分泌之增加倍數。橫條圖顯示了來自4個NHBE和3個DHBE供體的均值和SEM;
[圖 12A
]:
顯示了用還原型IL-33、oxIL-33或EGF處理後A549細胞之相對劃痕癒合密度。橫條圖顯示了每種條件下6次技術重複的均值和SEM;
[圖 12B
]:
顯示了用還原型IL-33、oxIL-33或EGF處理後NHBE細胞之相對劃痕癒合密度。橫條圖顯示了每種條件下6次技術重複的均值和SEM;
[圖 13
]:顯示了在抗ST2、抗RAGE或抗EGFR存在下,用單獨的培養基(未刺激的對照)、還原型IL-33、氧化型IL-33或氧化型IL-33處理的NHBE細胞之劃痕閉合百分比。橫條圖顯示了每種條件下6次技術重複之均值和SEM;
[圖 14
]:顯示了在經和不經氧化型IL-33刺激下,來自健康受試者、吸煙者和COPD的人支氣管上皮細胞中的相對劃痕癒合密度;
[圖 15
]:顯示了與DHBE COPD細胞(n = 5個供體)和用IgG1對照、抗IL-33(33_640087-7B)、抗RAGE(M4F4)和抗ST2處理的DHBE相比,NHBE細胞(n = 5個供體)24小時的劃痕閉合(%)。橫條圖顯示了n = 5個獨立供體的均值和SEM;
[圖 16A
]:
顯示了來自健康供體的ALI培養物的基底(p63+;藍色)、纖毛(α微管蛋白;紫色)和杯狀(黏蛋白5ac + 黏蛋白B;黃色)細胞的代表性免疫組織化學染色。
[圖 16B
]:
顯示了用抗IL-33(33_640087-7B)或同種型對照抗體處理7天後,使用HALO軟體對各種上皮細胞類型的免疫組織化學之定量;顯示的數據係來自n = 2-3個獨立供體之均值和SEM。
[圖 16C
]:
顯示了用抗IL-33(33_640087-7B)或同種型對照抗體處理7天後,使用HALO軟體對杯狀細胞之定量;顯示的數據係來自n = 2-3個獨立供體之均值和SEM。
[圖 17
]:顯示了來源於健康者(1個供體)或COPD者(1個供體)的ALI培養物中的獨立黏蛋白(黏蛋白5AC和黏蛋白5B)之示例性染色,以及用抗IL-33(33_640087-7B)處理7天後COPD培養物中之黏蛋白染色減少。
[圖 18
]:顯示了tSNE圖,該等tSNE圖說明瞭與未處理相比,來自用抗IL-33處理的獨立供體的COPD ALI培養物中發現的細胞亞型的不同比例。
[圖 19A
]:
顯示了檢測來自正常供體的ALI培養物中杯狀細胞之代表性流動式細胞測量術等高線圖。Muc5B在x軸上,並且Muc5AC在y軸上。用蛋白質處理ALI培養物7天。用還原型IL-33(IL-33[C- > S])處理不會使杯狀細胞增加至高於基線。oxIL-33(氧化型IL-33-01)使杯狀細胞百分比增加,IL-13也如此。已知IL-13可增加ALI培養物中的杯狀細胞,並在本研究中用作陽性對照。象限中的數字顯示總群體數的百分比:左上象限中是Muc5AC單陽性杯狀細胞,右下象限中是Muc5B單陽性杯狀細胞,並且右上象限中是Muc5AC和Muc5B雙陽性杯狀細胞。
[圖 19B
]:
顯示了來自正常供體(n = 6)的ALI培養物之合併流動式細胞測量術數據,該等數據顯示了杯狀細胞(合併的Muc5AC單陽性、Muc5B單陽性以及Muc5AC和Muc5B雙陽性杯狀細胞)占總上皮群體數的百分比。還原型IL-33(IL-33[C- > S])不會使杯狀細胞增加至高於基線。oxIL-33(氧化型IL-33-01)與IL-13一樣使杯狀細胞百分比增加。小提琴圖顯示了所有數據點和中位數。
[圖 19C
]:
顯示了來自正常供體(n = 6)的ALI培養物之合併流動式細胞測量術數據,該等數據顯示了Muc5AC單陽性杯狀細胞。還原型IL-33(IL-33[C- > S])不會使杯狀細胞增加至高於基線。oxIL-33(氧化型IL-33-01)與IL-13一樣使杯狀細胞百分比增加。小提琴圖顯示了所有數據點和中位數。
[圖 19D
]:
顯示了來自正常供體(n = 4)的ALI培養物之合併RT-qPCR數據,該等數據顯示了MUC5AC
mRNA的倍數變化。還原型IL-33(IL-33[C- > S])沒有增加MUC5AC
mRNA。oxIL-33(氧化型IL-33-01)與IL-13一樣引起MUC5AC
mRNA增加。小提琴圖顯示了所有數據點和中位數。
[圖 20A
]:
顯示了來自健康供體的ALI培養物之基底(p63+;紫色)、纖毛(α微管蛋白;深青色)和杯狀(Muc5ac + Muc5B;黃色)細胞之代表性免疫組織化學染色。還原型IL-33(IL-33[C- > S])在視覺上未能增加杯狀細胞。oxIL-33(氧化型IL-33-01)引起杯狀細胞的可見增加。
[圖 20B
]:
顯示了使用HALO軟體從免疫組織化學圖像(每個條件最少n = 3個供體)對黏蛋白5ac + 黏蛋白5b區域(總上皮組織區域%)進行的定量。與未處理的和還原型IL-33處理的對照相比,oxIL-33和IL-13增加了黏蛋白染色的區域。
[圖 21A
]:
顯示了檢測來自COPD供體的ALI培養物中杯狀細胞之代表性流動式細胞測量術等高線圖。如所描繪,Muc5B在x軸上,並且Muc5AC在y軸上。用抗體處理ALI培養物7天。抗IL-33(33_640087-7B)處理減少了總杯狀細胞數量。象限中的數字顯示總群體數之百分比:左上象限中是Muc5AC單陽性杯狀細胞,右下象限中是Muc5B單陽性杯狀細胞,並且右上象限中是Muc5AC和Muc5B雙陽性杯狀細胞
[圖 21B
]:
顯示了來自COPD供體(n = 6)的ALI培養物之合併流動式細胞測量術數據,該等數據顯示了總杯狀細胞數(合併的Muc5AC單陽性、Muc5B單陽性以及Muc5AC和Muc5B雙陽性杯狀細胞)。用抗體處理ALI培養物7天。抗IL-33(33_640087-7B)處理減少了總杯狀細胞數量。小提琴圖顯示了所有數據點和中位數。
[圖 21C
]:
顯示了來自COPD供體(n = 6)的ALI培養物之合併流動式細胞測量術數據,該等數據顯示了Muc5AC單陽性杯狀細胞。用抗體處理ALI培養物7天。抗IL-33(33_640087-7B)處理減少了Muc5AC單陽性杯狀細胞數量。小提琴圖顯示了所有數據點和中位數。
[圖 21D
]:
顯示了來自COPD供體(n = 5)的ALI培養物之合併RT-qPCR數據,該等數據顯示了MUC5AC
mRNA之倍數變化。抗IL-33(33_640087-7B)處理減少了MUC5AC
表現。小提琴圖顯示了所有數據點和中位數。
[圖 21E
]:
顯示了來自COPD供體(n = 6)的ALI培養物的合併流動式細胞測量術數據,該等數據顯示了藉由LD陰性細胞染色判斷的治療條件中的總活力。
[圖 22A
]:
顯示了來自COPD供體的ALI培養物的基底(p63+;紫色)、纖毛(α微管蛋白;深青色)和杯狀(Muc5ac + MucB;黃色)細胞之代表性免疫組織化學染色。抗IL-33(33_640087-7B)處理7天使杯狀細胞可見地減少。
[圖 22B
]:
顯示了使用HALO軟體從免疫組織化學圖像(n = 4個供體)對Muc5ac + Muc5b區域(總上皮組織區域%)進行之定量。與未處理的和人IgG1處理的對照相比,抗IL-33(33-640087_7B)減少了黏蛋白染色的區域。
[圖 23A
]:
顯示了對從COPD和健康ALI培養物獲得的頂端洗液中Muc5AC之定量。如藉由Muc5AC ELISA所判斷,COPD培養物中之Muc5AC水平較高
[圖 23B
]:
顯示了對從健康ALI培養物獲得的頂端洗液中Muc5AC之定量。用還原型IL-33mut16(IL-33[C- > S])、oxIL-33和野生型IL-33處理ALI培養物,藉由Muc5AC ELISA測定。
[圖 23C
]:
顯示了對從COPD ALI培養物獲得的頂端洗液中Muc5AC之定量。用人和小鼠IgG1對照(hIgG1和mIgG1)、33-640087_7B或抗ST2抗體處理細胞。如藉由Muc5AC ELISA所測定,用抗IL-33(33-640087_7B)處理降低了Muc5AC水平。實例 實例 1- 氧化型 IL-33 驅動 RAGE 與 EGFR 之間的傳訊複合物之形成
在Cohen, E. S. 等人, Nat. Commun. [自然通訊] 6: 8327 (2015) 中,申請人描述了IL-33的氧化型二硫鍵鍵合形式(DSB IL-33)之發現,並顯示此形式不結合ST2,且不能活化ST2依賴性傳訊。隨後(參見WO 2016156440 A1),申請人顯示oxIL-33結合晚期糖基化終產物受體(RAGE),並且以RAGE依賴性方式進行傳訊以活化STAT5並影響上皮遷移。
為了進一步探索oxIL-33的功能,用還原或氧化形式的IL-33刺激上皮細胞,並研究傳訊路徑。在本文中,本發明人顯示,oxIL-33係晚期糖基化終產物(RAGE)受體和表皮生長因子受體(EGFR)的複合物的新型配位基,從而對上皮功能產生深遠的影響。1. IL33 的人成熟和半胱胺酸突變變體的選殖和表現
藉由引物延伸PCR合成編碼人IL-33成熟組分(112-270)(登錄號(UniProt)095760)(也稱為IL33-01或IL-33)和4個半胱胺酸殘基突變為絲胺酸的變體(也稱為IL33-16或IL-33[C- > S])的cDNA分子,並將其選殖至pJexpress 411(DNA 2.0)中。修飾野生型(WT)和突變型IL-33編碼序列以在蛋白質的N末端含有10xHis、Avitag和因子Xa蛋白酶切割位點(MHHHHHHHHHHAAGLNDIFEAQKIEWHEAAIEGR SEQ ID NO: 43)。藉由轉化大腸桿菌BL21(DE3)細胞產生N末端標籤化的His10/Avitag IL33-01(WT,SEQ ID NO: 44)和N末端標籤化的His10/Avitag IL33-16(WT,SEQ ID NO: 45)。在37°C下,在自體誘導培養基(Overnight Express™自體誘導系統1,默克密理博公司(Merck Millipore),71300-4)中培養轉化的細胞18小時,隨後藉由離心收穫細胞且儲存在-20°C下。將細胞再懸浮於含有完全無EDTA的蛋白酶抑制劑混合物片劑(羅氏公司(Roche),11697498001)和50 U/ml全能核酸酶(Benzonase)(默克密理博公司,70746-3)的2x DPBS中,並藉由超音波處理裂解。藉由在4°C下以50,000 x g離心30 min來使細胞裂解物變澄清。藉由固定的金屬親和層析法,以5 ml/min載入到用2 x DPBS、1 mM DTT平衡的HisTrap excel柱(GE醫療公司(GE Healthcare),17371205)上來從上清液中純化IL-33蛋白。用2 x DPBS、1 mM DTT、20 mM咪唑(pH 7.4)洗滌柱以去除雜質,然後用2x DPBS、0.1% Triton X-114洗滌以去除固定的內毒素蛋白。用2x DPBS、1 mM DTT、20 mM咪唑(pH 7.4)進一步洗滌後,將樣本用2x DPBS、1 mM DTT、400 mM咪唑(pH 7.4)洗脫。藉由尺寸排阻層析法,使用在2x DPBS中的HiLoad Superdex 75 26/600 pg柱(GE醫療公司,28989334),以2.5 ml/min進一步純化IL-33。藉由SDS PAGE分析峰級分。將含有純IL-33的級分合併,並且藉由280 nm下的吸光度來確定濃度。藉由SDS-PAGE分析最終樣本。
為產生未標籤化的IL-33,使N末端標籤化的His10/Avitag IL33與每毫克蛋白質10個單位因子Xa(GE醫療公司,27084901)在室溫下在2x DPBS緩衝液中一起孵育1小時。在2x DPBS中,在HiLoad 16/600 Superdex 75 pg柱(GE醫療公司,28989333)上,以1 ml/min的流速使用SEC層析法來純化未標籤化的IL-33。2. 氧化型 IL-33 ( oxIL-33 )的產生和純化
藉由在60% IMDM培養基(無酚紅)、40% DPBS中稀釋至0.5 mg/ml的最終濃度,並在37°C下孵育18小時,將還原型IL33-01氧化。藉由將樣本載入到HiTrap Capto Q ImpRes陰離子交換柱(GE醫療公司,17547055)上,從樣本中去除氧化過程中產生的聚集體。載入之前,藉由添加1 M Tris(pH 9.0)直至pH達到8.3並添加5 M NaCl至125 mM的最終濃度來修飾樣本-在該等載入條件下,聚集體與柱結合並且單體oxIL-33流過而沒有結合並被收集。藉由在22°C下與因子Xa(NEB,P8010L)以每50 µg oxIL-33 1 µg因子Xa的最終濃度孵育120 min來從oxIL-33切割標籤。為了消耗具有任何剩餘還原型IL-33的樣本,將與人IgG1 Fc-His6融合的可溶性人ST2S胞外結構域與樣本在22°C下孵育30 min,並結合還原型IL-33。將樣本在截留值為3,000 Da的離心濃縮器中進行濃縮,並以2 ml/min的流速載入到HiLoad Superdex 75 26/600 pg柱(GE醫療公司,28989334)上,從而將單體oxIL-33與其他樣本組分分離。合併含有純oxIL-33的級分並濃縮,並且藉由UV吸收光譜法在280 nm下測定樣本的最終濃度。最終產品品質藉由SDS-PAGE、HP-SEC和RP-HPLC進行評估。3. 人 ST2 ECD 的選殖、表現和純化
藉由PCR用引物擴增編碼不含內源訊息肽(胺基酸殘基19-328)的ST2的天然存在的ST2S可溶性同型(UniProt登錄號Q01638-2)的cDNA,該等引物編碼與Gibson組裝相容的延伸序列和與ST2S編碼序列的N端融合的CD33訊息肽。類似地擴增具有C末端His6-標籤的人IgG1 Fc的編碼序列。使用Gibson組裝用pDEST12.2 OriP組裝ST2S cDNA和IgG1 Fc-His6 cDNA,pDEST12.2 OriP係一種哺乳動物CMV啟動子驅動的表現載體,其帶有來自EBV的OriP複製起點,從而可在表現EBNA-1蛋白的細胞系中進行游離基因維持(episomal maintenance)。為進行蛋白質表現,使用聚乙烯亞胺作為轉染試劑,將質體暫態轉化到過表現EBNA-1的CHO細胞懸浮培養物中。轉染後7天收集含有分泌的ST2S-Fc-His6融合蛋白的條件培養基,並以2 ml/min載入到HiTrap MabSelect SuRe(蛋白A,GE醫療公司,11-0034-95)親和層析柱上。用2x DPBS洗滌柱,並用25 mM乙酸鈉(pH 3.6)洗脫蛋白質。合併含有ST2S-Fc-His6的級分,並以2 ml/min載入到在2x DPBS中平衡的HiLoad Superdex 200 26/600 pg柱(GE醫療公司,28989336)上。合併含有純ST2S-Fc-His6蛋白的級分,並藉由在280 nm下的吸光度確定濃度。藉由SDS-PAGE分析最終樣本。4. 人去唾液酸糖蛋白受體( ASGPR ) ECD 的選殖、表現和純化
在Geneart化學合成了編碼無細胞質和跨膜結構域(胺基酸殘基62-291)的去唾液酸糖蛋白受體(UniProt登錄號P07306)的胞外結構域(ECD)的cDNA,其具有CD33訊息肽後接與ECD結構域的N末端融合的His10_Avi Tag序列。將構建體直接選殖到pDEST12.2 OriP中,pDEST12.2 OriP係一種哺乳動物CMV啟動子驅動的表現載體,其帶有來自EBV的OriP複製起點,從而可在表現EBNA-1蛋白的細胞系中進行游離基因維持。為進行蛋白質表現,使用293 Fectin作為轉染試劑將質體暫態轉化到HEK Freestyle 293F細胞的懸浮培養物中。轉染後7天,藉由固定金屬親和層析法,以4 ml/min載入到在2x DPBS中平衡的HisTrap excel柱(GE醫療公司,17371205)上來收集含有分泌的HisAVi_hASGPR ECD融合蛋白的條件培養基。用2x DPBS、40 mM咪唑(pH 7.4)洗滌柱以去除雜質,並用2x DPBS、400 mM咪唑(pH 7.4)洗脫樣本。藉由尺寸排阻層析法,使用在2x DPBS中的HiLoad Superdex 75 16/600 pg柱(GE醫療公司,28-9893-33),以1 ml/min進一步純化人ASGPR ECD。藉由SDS PAGE分析峰級分。將含有純單體ASGPR的級分合併,並且藉由280 nm下的吸光度來確定濃度。藉由SDS-PAGE分析最終樣本。5. IL-33 的氧化形式活化 MAP 激酶路徑
從龍沙公司(Lonza)獲得正常人支氣管上皮(NHBE)細胞(CC-2540),並根據製造商的方案維持在完全BEGM培養基(龍沙公司)中。用急性酶(accutase)(PAA,#L1 1-007)收穫NHBE,並以1 x 106
個/2 ml接種在6孔培養皿(康寧公司(Corning)Costar,3516)中的培養基[BEGM(龍沙公司CC-3171)和補充套組(龍沙公司CC-4175)]中。將細胞在37°C,5% CO2
下孵育18-24小時。此後,吸出培養基,並將細胞用1 ml PBS洗滌兩次,隨後添加饑餓培養基(補充有1%青黴素/鏈黴素的BEGM(龍沙公司CC-3171))。然後在刺激前將該等板在37°C、5% CO2
下再孵育18-24小時。
MAP激酶磷酸化抗體陣列套組(ab211061)購自艾博抗公司(Abcam),並根據製造商的說明書進行實驗。對已經饑餓18-24 h的6孔培養皿中的NHBE不處理或用30 ng/ml還原型IL-33、IL-33-16或氧化型IL-33處理,隨後放回37°C,5% CO2
下的孵育箱後維持10 min(對於此測定中使用的活化劑,參見表2)。從孵育箱中移出該等板,並用冰冷的PBS洗滌細胞,隨後添加100 µl/孔由套組提供的1x裂解緩衝液。將蛋白質提取物轉移至1.5 ml管中,隨後在4°C下以14,000 rpm使其變澄清。使用BCA技術(賽默公司(Thermo),23225)測定蛋白質濃度,並且每個陣列膜使用250 µg總蛋白。按照製造商的說明書執行所有後續步驟。將膜在LiCor C-digit上視覺化,並使用Image Lite studio定量。
[表 2
]
| 促效劑 | 識別符 | 重構於 | 最終濃度( µg/ml ) |
| 未標籤化的氧化型IL33-01 | RD15 | PBS | 100 |
| 未標籤化的IL33-01 | 07/24/2015 | PBS | 100 |
| 未標籤化的IL33-16 | 11/12/2015 | PBS | 100 |
| EGF | 236-EG-200 | PBS | 100 |
與野生型(IL-33)和IL-33的C- > S(IL-33[C- > S])還原形式(分別為IL33-01和IL33-16)相比,氧化型IL33-01(oxIL-33)活化了與受體酪胺酸激酶(RTK)參與的路徑一致的多個關鍵傳訊分子(圖1)。6. IL-33 的氧化形式活化了表皮生長因子受體( EGFR )
為了嘗試和鑒定被oxIL-33活化的受體酪胺酸激酶(RTK),使用71 RTK陣列進行篩選。RTK磷酸化抗體陣列套組(ab193662)購自艾博抗公司,並根據製造商的說明書進行實驗。培養NHBE,並以1 x 106
個/2 ml接種在6孔板(康寧公司Costar,3516)中的培養基[BEGM(龍沙公司CC-3171)和補充套組(龍沙公司CC-4175)]中。將細胞在37°C,5% CO2
下孵育18-24小時。此後,吸出培養基,並將細胞用1 ml PBS洗滌兩次,隨後添加饑餓培養基(無補充套組的BEGM(龍沙公司CC-3171))。然後在刺激前將該等板在37°C、5% CO2
下再孵育18-24小時。遵循先前對於MAP激酶陣列所述之相同步驟,將細胞活化(表2的活化劑),裂解,並且每個陣列膜使用250 µg總蛋白。按照製造商的說明書執行所有後續步驟。將膜在LiCor C-digit上視覺化,並使用Image Lite studio定量。未檢測到對還原的野生型(IL-33)或C- > S(IL-33 [C- > S])IL-33(分別為IL33-01和IL33-16)的反應。但是,oxIL-33(氧化型IL-33-01)在RTK陣列上觸發了與表皮生長因子受體(EGFR)相對應的陽性訊號(圖2)。
藉由其他方法證實了oxIL-33(氧化型IL-33-01)刺激EGFR傳訊的能力。活化後,將EGFR在Tyr1068處磷酸化,並且此磷酸-EGFR可以使用均相FRET(螢光共振能量轉移)HTRF®(均相時間分辨螢光,齊斯博國際公司(Cisbio International))測定(Cisbio套組#64EG1PEH)檢測到。簡言之,將NHBE以5 x 105
個/100 µl鋪在96孔板(康寧公司Costar,3598)中的培養基[BEGM(龍沙公司CC-3171)和補充套組(龍沙公司CC-4175)]中。將該等板在37°C,5% CO2
下孵育18-24小時。此後,吸出培養基,並將細胞用0.2 ml PBS洗滌兩次,隨後添加饑餓培養基(無補充套組的BEGM(龍沙公司CC-3171))。然後將板在37°C,5% CO2
下再孵育18-24小時,隨後用增加濃度的IL-33-01、IL-33-16和oxIL-33(氧化型IL-33-01)以及EGFR配位基(表2和表3)刺激,接著放回37°C,5% CO2
下的孵育箱後維持10 min。吸出培養基,並且每孔用50 µl裂解緩衝液(齊斯博公司,64EG1PEH)替換。然後按照製造商的說明書(齊斯博公司,64EG1PEH)進行測定。使用EnVision讀板儀(珀金埃爾默公司)在620 nm和665 nm發射波長下讀取時間分辨螢光。藉由計算665/620 nm比率和使用GraphPad Prism軟體藉由使用四參數對數方程進行曲線擬合確定的EC50值來分析數據。
[表 3
]
| 促效劑 | 供應商 | 識別符 | 重構於 | 最終濃度( µg/ml ) |
| TGFα | R & D系統公司 | 239-A-100 | 10 mM乙酸 | 100 |
| HB-EGF | R & D系統公司 | 259-HE-050/CF | PBS | 100 |
| 雙調蛋白(AREG) | R & D系統公司 | 262-AR-100/CF | PBS | 100 |
| β細胞素/BTC | R & D系統公司 | 261-CE-010/CF | PBS | 100 |
| 上皮調節蛋白 | R & D系統公司 | 1195-EP-025/CF | PBS | 100 |
| 上皮細胞有絲分裂蛋白 | R & D系統公司 | 6629-EP-025/CF | PBS | 100 |
| HMGB1 | R & D系統公司 | 1690-HMB-050 | PBS | 200 |
| S100A8/A9 | R & D系統公司 | 8226-S8-050 | PBS | 500 |
| S100A12 | R & D系統公司 | 1052-ER-050 | PBS | 200 |
| S100B | R & D系統公司 | 1820-SB-050 | PBS | 200 |
類似地,使用如本節先前提及的HTRF測定評定上皮細胞系A549中的EGFR磷酸化。簡言之,從ATCC獲得A549,並在補充有1%青黴素/鏈黴素和10% FBS的RPMI GlutaMax培養基中培養。用急性酶(PAA,#L1 1-007)收穫細胞,以5 x 105
個/100 µl接種到96孔板中,並在37°C,5% CO2
下孵育18-24小時。然後將孔用100 µl PBS洗滌兩次,隨後添加100 µl饑餓培養基(補充有1%青黴素/鏈黴素的RPMI GlutaMax培養基),並在37°C,5% CO2
下孵育18-24小時。用增加濃度的IL-33-01、IL-33-16和oxIL-33-01(氧化型IL-33-01的同義詞)、EGFR配位基和RAGE配位基刺激細胞(表2和表3),隨後放回37°C,5% CO2
下的孵育箱後維持10 min。吸出培養基,並且每孔用50 µl裂解緩衝液(齊斯博公司,64EG1PEH)替換。然後按照製造商的說明書(齊斯博公司,64EG1PEH)進行測定。使用EnVision讀板儀(珀金埃爾默公司)在620 nm和665 nm發射波長下讀取時間分辨螢光。藉由計算665/620 nm比率和使用GraphPad Prism軟體藉由使用四參數對數方程進行曲線擬合確定的EC50值來分析數據。
在NHBE和A549細胞兩者中,oxIL-33均類似於真正的促效劑EGF促進EGFR的磷酸化(圖3)。此未被測試的其他RAGE配位基重複。7. 傳訊元件的西方墨點法
進行了西方墨點法實驗,以進一步研究響應於oxIL-33(氧化型IL33-01),EGFR傳訊複合物的哪些元件被活化。如以上在第5節所述,將NHBE培養並鋪在6孔培養皿中。血清饑餓後,用oxIL-33(30 ng/ml)刺激細胞5至240分鐘之間。然後吸出培養基,並用冰冷的PBS洗滌細胞,隨後添加150 µl裂解緩衝液[1x LDS樣本緩衝液(賽默公司,NP0008)、10 mM MgCl2(VWR,7786-30-3)、2.5% β-巰基乙醇(西格瑪公司(Sigma),M6250)和0.4 µg/ml全能核酸酶(密理博公司,70746)]。將細胞在冰上放置10 min,隨後將裂解物轉移至1.5 ml管中,並加熱至90°C後維持5 min。將溶液轉移至新的1.5 ml管中,並在4-12% SDS-PAGE凝膠(賽默公司,NW04127BOX)上在MES運行緩衝液(B0002)中運行10 µl樣本以及5 µl蛋白梯(伯樂公司(BioRad),1610374)。使用Transblot Turbo(伯樂公司)將凝膠轉移到PVDF膜(伯樂公司,1704156)上。將PVDF膜在含有5%脫脂奶粉(Marvel)的PBS-吐溫(tween)溶液中阻斷10分鐘。然後將膜與一級抗體在含有5% BSA的PBS-吐溫中在4°C下孵育過夜。然後將膜用PBS-吐溫洗滌五次,接著在室溫下在含有5%脫脂奶粉的PBS-吐溫中與HRP標籤化的二級抗體一起孵育1小時。然後用PBS-吐溫將膜洗滌五次,隨後添加ECL(伯樂公司,1705062)並用Licor C-digit視覺化。
結果顯示,oxIL-33-01活化了幾種EGFR傳訊元件(圖4)8. Ox-IL-33 誘導 STAT-5 磷酸化,這被 EGFR 中和 Ab 阻斷
接下來試圖確定是否可以藉由阻止與EGFR結合來抑制oxIL33介導的STAT5活化。簡言之,在補充有1%青黴素/鏈黴素和10% FBS的RPMI GlutaMax培養基中培養A549細胞。用急性酶收穫細胞,以5 x 105
個/100 µl接種到96孔板中,並在37°C,5% CO2
下孵育18-24小時。然後將孔用100 µl PBS洗滌兩次,隨後添加100 µl饑餓培養基(補充有1%青黴素/鏈黴素的RPMI GlutaMax培養基),並在37°C,5% CO2
下孵育18-24小時。將抗EGFR抗體(殖株LA1(05-101,密理博公司)或同種型對照(MAB002,R & D系統公司)以劑量依賴性方式添加至孔中,並將板放回孵育箱後維持30 min。然後用氧化型IL-33(30 ng/ml)刺激板30 min,隨後按照製造商的說明書,使用磷酸-STAT5 ELISA套組裂解緩衝液(85-86112-11,賽默飛世爾科技公司(ThermoFischer Scientific))裂解並顯影,接著讀取450 nM下的吸光度。如圖5所示,用oxIL-33-01活化的細胞呈現出STAT5磷酸化,該STAT5磷酸化在存在抗EGFR抗體的情況下會降低(圖5)。 實例 2- 氧化型 IL-33 誘導 EGFR 和 RAGE 之間的複合物形成 9. oxIL-33 誘導 EGFR 和 RAGE 之間的複合物形成
為了理解RAGE和EGFR如何參與促進oxIL-33的傳訊,進行了免疫沈澱實驗以探索傳訊複合物。首先,使抗EGFR抗體與Dynabead共價偶聯。按照製造商的說明書,將兩個100 µg小瓶的抗EGFR抗體(R & D系統公司,AF231)與40 mg Dynabead(賽默公司,14311D)一起孵育,並共價偶聯。成功偶聯後,將珠粒以30 mg/ml再懸浮於PBS中,並保持在4°C下。
從龍沙公司獲得NHBE(CC-2540),並且將冷凍小瓶以每培養皿1 x 106
個細胞直接接種到15 cm培養皿(賽默公司,157150)中。根據製造商的方案,將NHBE維持在完全BEGM培養基(龍沙公司)中1個月,其中每三天更換一次培養基,直到細胞達到匯合為止。將該等板在37°C,5% CO2
下孵育此時間段。在刺激前一天,將板用20 ml PBS洗滌兩次,隨後添加15 ml饑餓培養基(無補充套組的BEGM(龍沙公司CC-3171))。然後將板在37°C,5% CO2
下再孵育18-24小時,隨後用單獨的培養基(未刺激的對照)、30 ng/ml還原型IL-33-01、30 ng/mL氧化型IL-33-01或30 ng/mL EGF刺激,並放回37°C,5% CO2
後維持10 min。吸出培養基,並用冰冷的PBS洗滌板兩次,隨後每15 cm培養皿添加1 ml含有磷酸酶和蛋白酶抑制劑(賽默公司,78440)的裂解緩衝液(艾博抗公司,ab152163)。將細胞刮入裂解緩衝液中,隨後轉移至2 ml蛋白LoBind管(艾本德公司(Eppendorf),Z666513)中,並藉由在4°C下以14,000 rpm離心變得澄清。使用BCA套組(賽默公司,23225)測定蛋白質濃度,並用裂解緩衝液將所有蛋白質提取物標準化至3 mg/ml。將6 mg總蛋白提取物在具有100 µl抗EGFR Dynabead(如上所述)的2 ml潔淨LoBind管中孵育。然後將管放置在4°C下的立式圓筒形混合機上5 h。使用磁鐵(伯樂公司,1614916)將Dynabead固定,吸出蛋白質提取物,並用2 ml洗滌緩衝液1(50 mM Tris-HCl pH 7.5(賽默公司,15567027)、0.5 % TritonX 100(西格瑪公司,X100)、0.3 M NaCl)替換。將此再重複四次。然後用洗滌緩衝液2(50 mM Tris-HCl pH 7.5)以相同方式將珠粒再洗滌十次。在最後的洗滌步驟之後,將50 mM含50 µl 1% Rapigest(w/v)(沃特世公司(Waters),186001861)的Tris-HCl pH 8.0添加到珠粒中,並在60°C下加熱10 min。然後將上清液轉移至新的LoBind 2 ml管中。再將100 µl 50 mM Tris-HCl pH 8.0添加到樹脂中並混合,隨後將其與第一次洗脫物組合。然後添加TCEP(西格瑪公司,646547)至最終濃度為5 mM,並將樣本在60°C下加熱10 min。然後藉由在黑暗中在室溫下添加碘乙醯胺(西格瑪公司,16125)至10 mM使洗脫液烷基化20 min。藉由添加DTT(西格瑪公司,D5545)至10 mM淬滅烷基化。然後添加Tris-HCl緩衝液50 mM pH 8.0,得到500 µl的最終樣本體積。每管添加0.5 µg胰蛋白酶(普洛麥格公司(Promega),V5111),並在30°C下在400 rpm的振盪平臺上將樣本消化過夜。然後將樣本用三氟乙酸(西格瑪公司,302031)酸化至最終濃度為2.0%(v/v),並在37°C下孵育1 h時。然後將樣本以14,000 rpm離心30 min,並將上清液轉移至新的2 ml LoBind管中。然後按照製造商的說明書通過C18柱(賽默公司,87784)處理樣本。然後使用speed-vac乾燥樣本,隨後儲存在-20°C下。然後藉由肽質量指紋質譜法(PMF-LC-MS)分析樣本。使用Scaffold軟體分析結果。
在所有4種條件下均類似地檢測到了EGFR,表明免疫沈澱在所有樣本中均表現良好。與用IL33-01(IL-33)或EGF處理的樣本相比,在用oxIL-33處理的樣本中檢測到RAGE和IL-33,表明在傳訊期間oxIL-33和RAGE與EGFR相關。與先前用oxIL-33和EGF在該等細胞中活化EGFR的觀察結果一致,用該等配位基刺激後,檢測到先前報導的參與EGFR傳訊和內吞作用的蛋白質,但不是還原型IL33-01(表4)。
表4顯示了用還原型IL-33-01(IL-33)、oxIL-33(氧化型IL-33-01)或EGF刺激的NHBE的LCMS分析。用oxIL-33刺激後,檢測到IL-33和RAGE與EGFR複合,但用還原型IL33-01(IL-33)或EGF刺激後未檢測到。括弧指示對於每種蛋白質鑒定的獨特肽的數量。
[表 4
]
| 未刺激 | IL-33 | oxIL-33 | EGF |
| EGFR(63) | EGFR(62) | EGFR(60) | EGFR(57) |
| - | - | IL-33(11) | - |
| - | - | RAGE(11) | - |
| - | - | AP-2α1(20) | AP-2α1(14) |
| - | - | AP-2α2(16) | AP-2α2(10) |
| - | - | AP-2β(15) | AP-2β(16) |
| - | - | AP-2µ(20) | AP-2µ(20) |
| - | - | AP-2σ(10) | AP-2σ(11) |
| - | - | CBL-B(5) | CBL-B(4) |
為了證實該等觀察結果,還對根據上述方案製備的細胞裂解物進行了免疫沈澱和西方墨點法。確定NHBE蛋白提取物的濃度後,將3 mg總蛋白與6 µg抗EGFR抗體(R & D系統公司,AF231)一起在1.5 ml管中孵育,並放置在4°C下的立式圓筒形混合機(end-over-end mixer)上2.5 h。然後將1.5 mg蛋白A/G磁珠(賽默公司,88802)添加到每個管中,隨後將管放回4°C後在混合下再維持1 h。然後用磁鐵(伯樂公司,1614916)收集珠粒,並用500 µl(50 mM Tris(pH 7.5)、1% TritonX和0.25 M NaCl)洗滌三次,並用500 µl 10 mM Tris(pH 7.5)洗滌一次。然後使用35 µl具有還原劑(賽默公司,NP0004)的LDS樣本緩衝液(賽默公司,NP0008)從磁珠釋放蛋白質,並在95°C下加熱5分鐘。將溶液轉移至新的1.5 ml管中,並在4%-12% SDS-PAGE凝膠(賽默公司,NW04127BOX)上在MES運行緩衝液(B0002)中運行10 µl樣本以及5 µl蛋白梯(伯樂公司(BioRad),1610374)。使用Transblot Turbo(伯樂公司)將凝膠轉移到PVDF膜(伯樂公司,1704156)上。將PVDF膜在含有5%脫脂奶粉(Marvel)的PBS-吐溫(tween)溶液中阻斷10分鐘。然後將膜與一級抗體(抗EGFR(細胞傳訊技術公司(Cell Signaling Technology),2232)、抗RAGE(細胞傳訊技術公司,6996)或抗IL-33(R & D系統公司,AF3625))在含有5% BSA的PBS-吐溫中在4°C下孵育過夜。然後將膜用PBS-吐溫洗滌五次,接著在室溫下在含有5%脫脂奶粉的PBS-吐溫中與抗兔HRP標籤化的二級抗體(細胞傳訊技術公司,7074)或抗山羊HRP標籤化的二級抗體(R & D系統公司,HAF109)一起孵育1小時。然後用PBS-吐溫將膜洗滌五次,隨後添加ECL(伯樂公司,1705062)並用Licor C-digit視覺化。西方墨點法證實在oxIL-33存在下RAGE與EGFR共沈澱,而在EGF刺激下未檢測到RAGE(圖6)。該等發現揭露,RAGE和EGFR係氧化型IL-33傳訊複合物的功能部分。10. oxIL-33 需要 RAGE 與 EGFR 形成複合物
上述實驗顯示,oxIL-33係引起下游傳訊的EGF受體(EGFR)複合物的配位基。本節中的實驗經設計以判定oxIL-33是否係RAGE或EGFR的直接結合配位基。為了更多地瞭解有關傳訊複合物的形成並評定oxIL-33是否直接與EGFR相互作用,使用ELISA形式探索oxIL-33與RAGE、ST2-Fc和EGFR的結合。
蛋白質和修飾:遵循製造商的方案使用生物素連接酶(BirA)(Avidty,Bulk BirA)將含有Avitag序列模體(GLNDIFEAQKIEWHE SEQ ID NO: 46)的蛋白質生物素化。遵循製造商的方案使用EZ連接磺基-NHS-LC-生物素(賽默公司/皮爾斯公司(Pierce),21335)經由游離胺將本文所用的所有修飾的無Avitag的蛋白質生物素化。表5係所用生物素化蛋白質的清單。
[表 5
].
| 試劑 |
| 生物素化的EGF(賽默公司) |
| Avitag-人ASGPR |
| Avitag_IL-33-01(還原型IL-33) |
| Avitag_IL-33-01(氧化型IL-33) |
| Avitag_IL-33-16 |
| HMGB1 |
在室溫下用100 µl/孔生物素化的抗原(10 µg/ml,PBS中)塗布抗生蛋白鏈菌素板(賽墨科技公司(Thermo Scientific),AB-1226)後維持1小時。將板用200 µl PBS-T(PBS + 1%(v/v)吐溫-20)洗滌3次,並用300 µl/孔阻斷緩衝液(含1% BSA的PBS(西格瑪公司,A9576))阻斷1小時。將板用PBS-T洗滌3次。將RAGE-Fc(R & D系統公司#1145-RG)或ST2-Fc(R & D系統公司#523-ST)在含PBS的阻斷緩衝液中稀釋至10 µg/mL,添加至相關孔中,並在室溫下孵育1小時。可替代地,在存在或不存在含10 µg/mL未標籤化的RAGE(義翹神州生物公司(Sino Biological),11629-HCCH)的PBS的情況下,添加100 µl含10 µg/mL EGFR-Fc(R & D系統公司#344-ER-050)的PBS後維持1小時。將板用200 µl PBS-T洗滌3次。然後在室溫下用100 µl/孔在阻斷緩衝液中1 : 10000稀釋的抗人IgG HRP(西格瑪AO170,5.1 mg/mL)檢測RAGE-Fc、ST2-Fc和EGFR-Fc持續1小時。將板用PBS-T洗滌3次且用100 µl/孔TMB(西格瑪公司,T0440)顯影。用50 µl/孔的0.1 M H2
SO4
淬滅反應。在Cytation Gen5或類似設備上讀取450 nm下的吸光度。結果顯示,oxIL-33呈現出與RAGE明顯相互作用(圖7A),而oxIL-33與EGFR的直接結合可忽略(圖7B)。僅藉由向此測定中添加sRAGE可觀察到EGFR與oxIL-33結合(圖7B)。如果用oxIL-33取代真正的RAGE促效劑HMGB1,則此不能重現(圖7B)。
使用RAGE缺陷型細胞系進一步證實了oxIL-33觸發的EGFR傳訊中需要RAGE。如下產生敲除RAGE的A549細胞系:
產生哺乳動物質體,其含有紅色螢光蛋白(RFP)的表現載體、靶向AGER的外顯子3的指導RNA(TGAGGGGATTTTCCGGTGC SEQ ID NO: 47)和Cas9內切核酸酶。藉由使A549細胞在T-175燒瓶中在F12K堅果混合物(吉博科公司(Gibco),補充有10% FBS和1%青黴素/鏈黴素)中生長兩天來產生A549條件培養基。將消耗的培養基從A549取出,過濾,並在新鮮的吉博科公司F12K堅果混合物(補充有20% FBS和1%青黴素/鏈黴素)中5倍稀釋。將A549以2 x 105
個細胞/ml接種到三個T-75燒瓶中,共15 ml,並置於37°C,5% CO2
孵育箱中過夜。使用1.6 ml F12K堅果混合物(補充1%青黴素/鏈黴素)以及8 µg AGER指導RNA質體和22.5 µg PEI(波利塞斯公司(Polysciences),23966-2)製備轉染混合物。然後將混合物渦旋10秒,並在室溫下放置15 min。接著將0.75 ml轉染混合物添加到每個T-75燒瓶中。將燒瓶放回孵育箱後維持兩天。然後使用急性酶分離A549細胞,轉移到含1% FBS的PBS中,並根據RFP在96孔培養皿中的表現,在Aria細胞分選儀(BD)上分選單個細胞。每3-5天用條件培養基餵養細胞。細胞超過50%匯合後,將其轉移到24孔板中並使其生長。此升級過程一直持續到每個成功的殖株被分到T15燒瓶中。然後將細胞分到12孔板中,並生長直至超過50%匯合,隨後分析基因組PCR的成功敲除情況。使每孔的細胞在100 µl DNA裂解緩衝液(維亞根生物技術公司(Viagen Bitoech),301-C,補充有0.3 µg/ml蛋白酶K)中裂解。將該等樣本在55°C下孵育4小時,隨後在85°C下孵育15 min。使用具有以下序列的正向和反向引物進行RAGE的PCR:正向-gttgcagcctcccaacttc(SEQ ID NO: 48),反向-aatgaggccagtggaagtca(SEQ ID NO: 49)。反應和循環的設定如下:反應體積為50 µl [25 µl Q5聚合酶混合物、2.5 µl正向引物(10 µM儲備液)、2.5 µl反向引物(10 µM儲備液)、2 µl模板DNA裂解物、18 µl無核酸酶的水]。進行PCR反應,其中在98°C下進行初始變性30秒,隨後進行如下35個如下循環:在98°C下5秒,在57°C下10秒以及在72°C下20秒,隨後最後一步在72°C下2分鐘。將4 µl PCR產物與6 µl無核酸酶的水和2 µl 6x DNA載入緩衝液(賽默科技公司,R0611)混合。將樣本在90V下在1%瓊脂糖凝膠(1 : 10000 SYBR safe)上運行1小時,隨後在Versadoc成像儀上視覺化。然後遵循製造商的方案,使用QIAquick PCR純化套組(凱傑公司(Qiagen),28104)清理其餘的PCR產物。使用nanodrop測量DNA-50濃度。發送幾個殖株(從結果中選擇)進行內部定序。結果顯示在殖株RAGE09和RAGE10中成功插入了終止密碼子。
為了確定RAGE對oxIL-33介導的EGFR傳訊的必要性,然後對A549和RAGE缺陷型A549細胞進行了免疫沈澱和西方墨點法。簡言之,用oxIL-33-01在各時間點(0-15分鐘)活化細胞系。隨後進行EGFR或RAGE的免疫沈澱,然後遵循第9節中詳述的相關實驗方案用抗RAGE、抗EGFR和抗IL-33進行西方墨點法。結果顯示RAGE在與oxIL-33和EGFR形成複合物中的關鍵作用(圖8)11. 氧化型 IL-33 誘導 STAT5 磷酸化,該 STAT5 磷酸化被 RAGE 阻斷,但未被 ST2 中和抗體阻斷
為了證實oxIL-33傳訊中RAGE的重要性優於ST2,測試了阻斷抗體。簡言之,在補充有1%青黴素/鏈黴素和10% FBS的RPMI GlutaMax培養基中培養A549。用急性酶收穫細胞,以5 x 105
個/100 µl接種到96孔板中,並在37°C,5% CO2
下孵育18-24小時。然後將孔用100 µl PBS洗滌兩次,隨後添加100 µl饑餓培養基(補充有1%青黴素/鏈黴素的RPMI GlutaMax培養基),並在37°C,5% CO2
下孵育18-24小時。將抗RAGE(M4F4;WO 2008137552);抗ST2(AF532;RnD系統公司)或同種型對照(MAB002,R & D系統公司)以劑量依賴性方式添加至孔中,並將板放回孵育箱後維持30 min。然後用氧化型IL-33(30 ng/ml)刺激板30 min,隨後按照製造商的說明書,使用磷酸-STAT5 ELISA套組裂解緩衝液(85-86112-11,賽默飛世爾科技公司)裂解並顯影,接著讀取450 nM下的吸光度。如圖9所示,用oxIL-33-01活化的細胞呈現出STAT5磷酸化,該STAT5磷酸化在存在抗RAGE抗體而非抗ST2抗體的情況下會降低(圖9)。 實例 3-oxIL-33 觸發上皮細胞中 EGFR 的內在化
接下來研究了與EGF相比,oxIL-33是否誘導EGFR動力學變化。12. EGF 內在化的共聚焦實驗
此實驗旨在利用共聚焦成像研究用EGF、IL-33的還原或氧化形式刺激後上皮細胞中EGFR的動力學。將EGFR-GFP A549上皮細胞系(西格瑪公司,CLL1141-1VL)以20000個細胞/ml的濃度(RPMI培養基 + 10% FCS + Pen/Strep)鋪板,每24孔玻璃底板1 ml(格雷內爾公司(Greiner),662892)。與綠色螢光蛋白(GFP)連接的EGF受體可以追蹤EGFR膜動力學和內在化。用PBS洗滌細胞一次,並在RPMI培養基(無FCS)中孵育。饑餓24小時後,用RPMI洗滌細胞,並與0.5 ml RPMI培養基以及CellMask(英傑公司(Invitrogen)C10046)深紅以1 : 5000稀釋度一起孵育。進行膜標記用處理之前用CellMask對細胞進行短暫染色,並在共聚焦處理時立即以1幀/min進行即時成像,以記錄EGFR-GFP的動力學。將細胞在37°C下染色5分鐘,用PBS洗滌一次,並用200 ng/ml濃度的oxIL-33(氧化型IL-33-01)或IL-33-16以0.5 ml無血清RPMI/孔刺激。立即以40x油物鏡,1 min/幀,5疊間隔2 µm拍攝共聚焦圖像,持續25分鐘(添加蛋白質後約30分鐘)。GFP訊號的中斷(虛線)圖譜指示受體在膜上聚類和內在化。在即時成像的不同時間點產生膜區(被CellMask遮蓋)和胞內區(被倒置的CellMask遮蓋,未顯示)的像素強度長條圖,顯示了非聚類區中EGFR的消耗(長條圖鐘形峰左移),以及藉由聚類引起的飽和像素數量增加(強度255)。oxIL-33誘導EGF受體的聚類和內在化,但EGF刺激引起最明顯的EGFR聚類。相反,IL-33的還原形式(IL -33-16)在EGFR細胞分佈方面未呈現出大的變化(圖10)。 實例 4- 類似於 EGF , oxIL-33 誘導上皮細胞分泌 IL-8 13. 藉由 oxIL-33 進行的 IL-8 的選擇性分泌
根據製造商的方案,將來自健康受試者(NHBE;龍沙公司CC-2540)和慢性阻塞性肺病(COPD)(DHBE;龍沙公司00195275)的人支氣管上皮細胞在完全BEGM培養基(龍沙公司)中維持一個月,其中每三天更換一次培養基直到細胞達到匯合。用急性酶收穫細胞,並以5 x 105
個/100 µl接種在96孔板(康寧公司3596)中的培養基中。將該等板在37°C,5% CO2
下孵育18-24小時。此後,吸出培養基,並將細胞用100 µl PBS洗滌兩次,隨後添加饑餓培養基(無補充有1%青黴素/鏈黴素的補充套組的BEGM(龍沙公司CC-3171))。然後將板在37°C,5% CO2
下再孵育18-24小時,隨後用單獨的培養基(未刺激的對照)、30 ng/ml還原型IL-33-01、30 ng/mL IL-33-16、30 ng/mL氧化型IL-33-01或30 ng/mL EGF刺激,並放回37°C,5% CO2
。刺激後24小時,收集上清液並使用多重測定(Mesoscale Discovery K15047D-2)評價趨化因子產生。如圖11中所示,與未刺激的細胞(單獨的培養基)相比,用oxIL-33活化後,NHBE和DHBE的IL-8分泌增加4倍。其他趨化因子(TARC、MIP-1a、MIP1b、MCP4、MCP1、IP10、嗜酸細胞活化趨化因子(Eotaxin)、嗜酸細胞活化趨化因子-3、MDC-未顯示數據)未觀察到大的差異。 實例 5-oxIL-33 損害深層單層上皮培養物中的劃痕修復反應 14. 與 EGF 相反, oxIL-33 損害 A549 和 NHBE 細胞中的劃痕閉合
從ATCC獲得A549,並在補充有1%青黴素/鏈黴素和10% FBS的RPMI GlutaMax培養基中培養。用急性酶(PAA,#L1 1-007)收穫細胞,以5 x 105
個/100 µl接種到96孔板中,並在37°C,5% CO2
下孵育18-24小時。然後將孔用100 µl PBS洗滌兩次,隨後添加100 µl饑餓培養基(補充有1%青黴素/鏈黴素的RPMI GlutaMax培養基),並在37°C,5% CO2
下孵育18-24小時。使用WoundMakerTM(埃森生物科學公司(Essen Bioscience))劃去細胞,然後將孔用200 µl PBS洗滌2次,隨後添加補充有0.1% FBS(v/v)和1%(v/v)青黴素/鏈黴素的、含有指定刺激的RPMI GlutaMax培養基或單獨的培養基(未刺激的對照)、30 ng/ml還原型IL-33-01、30 ng/mL氧化型IL-33-01或30 ng/mL EGF,並放回37°C,5% CO2
。將板放置在IncucyteZoom中以在48小時內進行劃痕癒合成像和分析。藉由Incucyte Zoom軟體中的劃痕癒合演算法計算相對劃痕密度。
從龍沙公司獲得NHBE(CC-2540),並根據製造商的方案維持在完全BEGM培養基[BEGM(龍沙公司CC-3171)和補充套組(龍沙公司CC-4175)]中。用急性酶收穫細胞,並以5 x 105
個/100 µl接種在96孔ImageLock板(賽多利斯公司(Sartorius),4379)中的培養基中。將該等板在37°C,5% CO2
下孵育18-24小時。此後,吸出培養基,並將細胞用100 µl PBS洗滌兩次,隨後添加饑餓培養基(無補充有1%青黴素/鏈黴素的補充套組的BEGM(龍沙公司CC-3171))。然後在產生劃痕前將該等板在37°C、5% CO2
下再孵育18-24小時。使用WoundMakerTM(埃森生物科學公司)劃去細胞,然後將孔用200 µl PBS洗滌2次,隨後添加補充有0.1% FBS(v/v)和1%(v/v)青黴素/鏈黴素的、含有指定刺激的BEBM培養基(龍沙公司)或單獨的培養基(未刺激的對照)、30 ng/ml還原型IL-33-01、30 ng/mL氧化型IL-33-01或30 ng/mL EGF,並放回37°C,5% CO2
。將板放置在IncucyteZoom中以在48小時內進行劃痕癒合成像和分析。藉由Incucyte Zoom軟體中的劃痕癒合演算法計算相對劃痕密度。如圖12所示,oxIL-33抑制A549細胞(圖12A)和NHBE細胞(圖12B)的深層培養物中的劃痕癒合,與觀察到劃痕細胞密度增加的EGF有相反的作用。15. 可以藉由中和 RAGE 或 EGFR 而非 ST2 的抗體來阻止氧化型 IL-33 對劃痕閉合的損害
為了瞭解oxIL-33的該等功能作用是否藉由RAGE/EGFR介導,如第14節所述,但在中和不同受體組分的抗體存在下,在NHBE細胞中進行劃痕測定。在10 µg/mL抗ST2(AF532,R & D系統公司)、抗RAGE(M4F4,WO 2008137552)或抗EGFR(殖株LA1,05-101密理博)存在下,用單獨的培養基(未刺激的對照)、還原型IL-33或氧化型IL-33處理NHBE細胞。oxIL-33,但不是還原型IL-33,抑制劃痕閉合。oxIL-33的此作用可被抗RAGE和抗EGFR而非抗ST2逆轉,再次證明RAGE和EGFR係參與氧化型IL-33傳訊路徑的必需受體(圖13)。 實例 6- 抗 IL-33 改進了深層培養物中 COPD 細胞的表型 16. oxIL-33 可以在劃痕閉合測定中驅動健康的 NHBE 中的 COPD 樣反應
接下來,研究了氧化型IL-33在健康者、吸煙者和COPD支氣管上皮細胞中的作用。從龍沙公司獲得NHBE(CC-2540)、來自吸煙者的NHBE(CC-2540)和DHBE(COPD,00195275),並根據製造商的方案保持在完全BEGM培養基(龍沙公司)中。如在第14節中所述進行劃痕測定。用單獨的培養基(未刺激的對照)或30 ng/mL氧化型IL-33處理細胞。與來自健康受試者的細胞相比,來自吸煙者或COPD的支氣管上皮細胞顯示的劃痕閉合能力受損,這與用oxIL-33處理健康細胞後觀察到的損害類似(圖14)。與健康細胞相比,在吸煙者和COPD HBE細胞中,oxIL-33不會進一步損害劃痕閉合反應(圖14)。17. 藉由 RAGE/EGFR 路徑阻斷內源 IL-33 可以改進 COPD 基底細胞受損的劃痕修復表型。
由於已知上皮細胞會產生IL-33,所以自分泌IL-33分泌可能會導致在COPD細胞中觀察到的劃痕修復表型受損。為了研究,在IL-33中和存在下在來自COPD的支氣管上皮細胞中進行了劃痕閉合測定。根據製造商的方案,將NHBE(龍沙公司CC-2540)和DHBE(龍沙公司,COPD 00195275)維持在完全BEGM培養基(龍沙公司)中。用急性酶收穫細胞,並以5 x 105
個/100 µl接種在96孔ImageLock板(賽多利斯公司(Sartorius),4379)中的培養基中。將該等板在37°C,5% CO2
下孵育18-24小時。此後,吸出培養基,並將細胞用100 µl PBS洗滌兩次,隨後添加饑餓培養基(無補充有1%青黴素/鏈黴素的補充套組的BEGM(龍沙公司CC-3171))。然後在產生劃痕前將該等板在37°C、5% CO2
下再孵育18-24小時。使用WoundMakerTM(埃森生物科學公司)劃去細胞,然後將孔用200 µl PBS洗滌2次,隨後添加含有10 µg/mL抗IL-33(33_640087-7B,描述於WO 2016/156440中)、抗ST2(AF532,R & D系統公司)、抗RAGE(M4F4,WO 2008137552)或NIP228(IgG1同種型對照)的補充有0.1% FBS(v/v)和1%(v/v)青黴素/鏈黴素的BEBM培養基(龍沙公司),並放回37°C,5% CO2
。將板放置在IncucyteZoom中以在48小時內進行劃痕癒合成像和分析。藉由Incucyte Zoom軟體中的劃痕癒合演算法計算相對劃痕密度。如先前所觀察到的,與來源於健康受試者的細胞相比,COPD細胞的劃痕閉合反應受損。抗IL-33和抗RAGE而非抗ST2能夠將COPD細胞的劃痕閉合反應改進到與健康細胞類似的水平(圖15),證明上皮細胞產生自分泌的IL-33,其通過RAGE/EGFR路徑傳導訊號(圖15)。 實例 7- 抗 IL-33 減少 3D 上皮培養物中的杯狀細胞 18. 氣道基底細胞的氣液介面( ALI )培養
接下來,本發明人試圖確定氣液介面細胞培養物(「ALI培養物」)中IL-33傳訊的相關性。ALI培養係基底細胞生長的一種方法,其中該等基底細胞的基底表面與培養基接觸,並且頂部(頂端)細胞層暴露於空氣。ALI培養能夠在體外產生類似於氣管上皮的具有假複層上皮的黏膜纖毛表型的三維細胞結構。因此,ALI培養物可以用於研究呼吸上皮的基本方面,諸如細胞間傳訊、疾病建模和呼吸再生。
從北卡羅來納大學(University of North Carolina)和匹茲堡大學(University of Pittsburgh)接收到具有來自健康對照或COPD患者的冷凍肺基底細胞的冷凍小瓶。將細胞融化並鋪在塗有在1X PBS(麻塞諸塞州沃爾瑟姆市吉博科公司(Gibco, Waltham, MA))中1 : 70稀釋的Purecol I型牛膠原蛋白(加利福尼亞州聖地牙哥市高級生物基質公司(Advanced BioMatrix, San Diego, CA))的T-75燒瓶上,並在Epix培養基(276-201,馬里蘭州羅克維爾市普羅帕吉尼斯公司(Propagenix, Rockville, MD))中生長。達到匯合後,將該等細胞分到適當數量的T-75燒瓶一次,隨後收穫以進行ALI培養。含有12 mm 0.4 µM聚酯膜插入物(Costar,紐約州康寧公司)的ALI培養用Transwell藉由用1 : 70 Purecol溶液塗布插入物,並在37°C下孵育1-16小時之間來製備。去除Purecol溶液,並將Transwell放置在UV光下30分鐘,然後用PBS洗滌。使用4 ml胰蛋白酶溶液(賽默飛世爾公司,15400054)分離T-75燒瓶中的基底細胞。將細胞懸浮液添加到含有5 ml FBS的50 ml管中,然後在ViCell計數器(加利福尼亞州佈雷亞市貝克曼庫爾特公司(Beckman Coulter, Brea, CA))上計數,並以1,000 RPM離心5分鐘。然後將細胞以3.57 x 105
個/ml的密度再懸浮於Pneumacult ALI培養基(不列顛哥倫比亞省溫哥華市幹細胞技術公司(Stemcell Tech, Vancouver, BC))中,並將700 µl分配到每個Transwell上。將1 mL ALI培養基添加到插入物下方的空間中。使細胞處於ALI培養基的深層,直到形成匯合和緊密連接(典型地7天),此時從頂側去除培養基,使細胞分化2週,其中每隔一天在基底側更換培養基一次。藉由在供應至培養物基底側的培養基中包括處理來將完全分化的培養物在無抗體下處理,用1 µg/ml抗IL-33(33_640087-7B)或1 µg/ml NIP228(IgG1同種型對照)處理7天。每隔一天進行一次培養基(含有相關處理)更換。19. IHC 三重染色(基底、杯狀和纖毛)和定量
如第18節中所述產生來自COPD供體的ALI培養物並處理。將ALI上皮培養物在10%中性緩衝福馬林(formalin)中固定24小時,並包埋在石蠟中。將石蠟切片(4 um)封固在帶正電荷的載玻片上,並藉由連續的3重發色測定在Ventana Discovery Ultra上染色。用細胞調節劑1(CC1)(目錄號5424569001,羅氏公司)進行抗原修復,並用Discovery抑制劑(目錄號7017944001,羅氏公司)阻斷內源過氧化物酶12 min。在36°C下施加抗p63(殖株4A4)(目錄號790-4509,瑞士巴塞爾市羅氏公司(Roche, Basel, Switzerland))24 min,並用小鼠抗HQ(12 min)(目錄號7017782001,羅氏公司)和抗HQ HRP(12 min)(目錄號7017936001,羅氏公司)視覺化,並在Teal底物(目錄號8254338001,羅氏公司)中孵育12 min。藉由抗體變性步驟(100°C,24 min),用細胞調節劑2(CC2)(目錄號5424542001,羅氏公司),然後用以Dako抗體稀釋劑(目錄號S3022)稀釋的0.01 µg/ml抗微管蛋白(目錄號ab24610,英國劍橋市艾博抗公司)處理載玻片16 min,用小鼠OmniMap-HRP(目錄號5269652001,羅氏公司)檢測(8 min),並用Discovery Purple底物(目錄號7053983001,羅氏公司)視覺化16 min。將載玻片用CC2再進行抗體變性,然後施加1.1 µg/ml兔抗黏蛋白5AC和7 µg/ml兔抗黏蛋白5B(分別為目錄號ab198294和目錄號ab87376,艾博抗公司)的混合物20 min,並用抗兔NP(4 min)(目錄號7425317001,羅氏公司)、抗NP-AP(8 min)(目錄號7425325001,羅氏公司)視覺化,然後用Discovery Yellow(目錄號7698445001,羅氏公司)視覺化20 min。將染色的載玻片用Dawn洗滌劑沖洗,用蘇木精(目錄號5277965001,羅氏公司)複染,沖洗,用分級的乙醇和二甲苯系列脫水,並用永久封片劑(mounting media)封固。使用HALO軟體進行的定量顯示,來源於用抗IL-33治療的健康供體的ALI培養物中的杯狀細胞減少(圖16)。 實例 8- 抗 IL-33 調節來自 COPD 的 3D 上皮培養物中的黏蛋白並改進黏液運動 20. IHC 雙重 IF 染色(黏蛋白 5B + 黏蛋白 5AC )
如第18節中所述產生來自COPD供體的ALI培養物並處理。將ALI上皮培養物在10%中性緩衝福馬林(formalin)中固定24小時,並包埋在石蠟中。將石蠟切片(4 um)封固在帶正電荷的載玻片上,並藉由連續的2重免疫螢光測定在Ventana Discovery Ultra上染色。用細胞調節劑1(CC1)進行抗原修復,用Discovery抑制劑阻斷內源過氧化物酶12 min並用羅氏診斷公司(Roche Diagnostics)的S Block(RUO)(目錄號760-4212)阻斷8 min,並且在36°C下在用Dako Ab稀釋劑S3022稀釋的7 µg/ml抗黏蛋白5B中孵育24 min,並用抗兔HQ(羅氏診斷公司目錄號760-4815)檢測4 min且用抗HQ-HRP(羅氏診斷公司目錄號760-4820)檢測8 min。然後將樣本與酪醯胺軛合物(conjugate)Discovery FITC(羅氏診斷公司目錄號760-232)一起孵育8 min。在Discovery Ultra程式中選擇了雙重序列,並藉由抗體變性步驟(100°C 24 min)用細胞調節劑2(CC2)處理樣本,然後用Discovery抑制劑(40C,24 min)中和,隨後在36°C下施加抗黏蛋白5AC,1.1 µg/ml,20 min。用抗兔-HQ(羅氏診斷公司目錄號760-4815)4 min和抗HQ-HRP(羅氏診斷公司目錄號760-4820)8 min檢測黏蛋白5AC,並用酪醯胺軛合物Discovery Red610視覺化8 min。完成此步驟後,將染色的載玻片從Discovery Ultra自動染色機取出,並用Dawn洗滌劑沖洗,然後用去離子水沖洗。將樣本在1 µg/ml在去離子水中稀釋的4′,6-二脒基-2-苯基吲哚二鹽酸鹽(DAPI核酸染料)(賽默飛世爾公司目錄號D1306)中孵育2 min。用去離子水沖洗樣本,並在ProLong Gold Antifade封片劑(賽默飛世爾公司,目錄號P36930)存在下置放蓋玻片,並儲存在不透光的載玻片盒中。用Zeiss LSM 880共聚焦顯微鏡(紐約州白原市卡爾蔡司顯微鏡有限責任公司(Carl Zeiss Microscopy, LLC, White Plains, NY))對染色的載玻片成像。圖17顯示ALI培養物的抗IL-33處理可能引起COPD培養物中不同黏蛋白的下調21. 抗 IL-33 逆轉了 COPD ALI 培養物中觀察到的受損的黏膜纖毛清除
如第18節中所述產生來自COPD供體的ALI培養物並處理。然後將30 µl在PBS中1 : 33稀釋的0.2 µM FluoSpheres(賽默飛世爾公司,F8811)添加到頂端表面,並使用蔡司LSM800顯微鏡捕獲FluoSphere運動的短片,並且顯示用抗IL-33(33_640087-7B)而非對照抗體處理後黏膜纖毛運動增加。 實例 9-ALI 培養物的單細胞 RNA 分析顯示用抗 IL-33 處理後杯狀細胞變化
如第18節中所述產生來自COPD供體的ALI培養物並處理。為了獲得單細胞懸浮液,將過濾器插入物與0.25%胰蛋白酶在37°C下孵育5 min。藉由用PBS上下吸取洗滌來輕輕地從過濾器分離上皮細胞,然後轉移到15 ml Falcon管中。將細胞在4°C下以1000 RPM離心5 min。去除上清液後,將細胞再懸浮於含0.4% BSA的PBS中,並將細胞濃度調節至1000個細胞/µl以進行定序。根據標準方案,將細胞懸浮液載入到Chromium單細胞3’套組中,以捕獲5000至10.000個細胞/個通道。使用版本2化學法。遵循製造商的方案(Chromium™單細胞3’套組,v2化學法)獲得用於依諾米那定序(Illumina sequencing)的單細胞3’文庫。評定文庫的品質(TapeStation 4200,安捷倫公司(Agilent)),然後在NextSeq 500或NovaSeq 6000儀器(依諾米那公司(Illumina))上定序。使用Cell Ranger 2.0版過程(10x Genomics)進行初始數據處理。使用Seurat包進行後處理,以排除低細胞品質和標準化。每個樣本均作為獨立數據進行分析,以捕獲樣本內異質性(細胞亞型)。使用t分佈隨機近鄰嵌入(t-Distributed Stochastic Neighbor Embedding;tSNE)實現聚類和視覺化。COPD中細胞類群的鑒定由標記基因指導。對於其他樣本,手動檢查初始類群,然後將Seurat的標記轉移演算法(Label Transfer algorithm)施加於亞型鑒定。對來自進行和未進行如第18節中所提及的抗IL-33處理的COPD ALI培養物的細胞之間的每個類群進行MUC5AC和MUC5B基因表現分析。使用Seurat產生熱圖和tSNE圖。圖18顯示了tSNE圖,該等tSNE圖說明瞭與未處理相比,用抗IL-33(33_640087-7B)處理的ALI培養物中發現的細胞亞型的不同比例。如圖18所示,抗IL-33處理後注意到MUC5B高細胞減少。 實例 10- 抗 IL-33 減少了 COPD 3D 上皮培養物中的杯狀細胞 22. 氣道基底細胞的氣液介面( ALI )培養
為了定量和詢問oxIL-33在生理學相關的氣液介面(ALI)培養系統中的作用,開發了流動式細胞測量術測定,該測定旨在區分杯狀細胞類型(MUC5ac相對於MUC5b)與其餘上皮群體(黏蛋白陰性)。
從龍沙公司接收到具有來自健康(CC-2540)對照或COPD(195275)患者的冷凍肺基底細胞的冷凍小瓶。將每個供體一個小瓶融化並鋪在4個T-175燒瓶中的Epix培養基(276-201,馬里蘭州羅克維爾市普羅帕吉尼斯公司)中。達到匯合後,將該等細胞在P2下以每個小瓶1e6個細胞冷凍。將P2下的細胞新加入2個T-75燒瓶中的Epix培養基中,並生長直至80%匯合。含有12 mm或6.5 mm 0.4 µM聚酯膜插入物(Costar,紐約州康寧公司)的ALI培養用Transwell藉由用1x膠原蛋白I溶液(Celladhere™膠原蛋白I-幹細胞#07001,在dH2O中製備)塗布插入物,並在37°C下孵育1-16小時之間來製備。去除膠原蛋白I溶液,並用PBS洗滌Transwell。用PBS洗滌T-75燒瓶中的基底細胞,並使用6 ml胰蛋白酶溶液(龍沙公司胰蛋白酶繼代培養包-#CC-5034)分離。用6 ml胰蛋白酶中和溶液(龍沙公司胰蛋白酶繼代培養包-#CC-5034)中和胰蛋白酶,並將細胞懸浮液添加到15 ml管中,計數並以1200 RPM離心5分鐘。然後將細胞以8 x 105
個/ml的密度再懸浮於Pneumacult ALI培養基(不列顛哥倫比亞省溫哥華市幹細胞技術公司)中,並將0.5 ml分配到每個12 mm Transwell上,並且將0.25 ml分配到每個6.5 mm Transwell上。將1 mL ALI培養基添加到12 ml插入物下方的空間中,並且將0.5 ml ALI培養基添加到6.5 mm插入物下方的空間中。使細胞處於ALI培養基的深層,直到形成匯合和緊密連接(典型地7天),此時從頂側去除培養基,使細胞分化3週,其中每個週一、週三和週五在基底側更換培養基一次。對完全分化的正常培養物不處理,或藉由在供應至培養物基底側的培養基中包括處理(7天處理)來用還原型或氧化型未標籤化的IL33-01(30 ng/ml)、未標籤化的IL33-16(30 ng/ml)、IL-13(10 ng/ml)、EGF(30 ng/ml)或HMGB1(30 ng/ml)處理7天。對完全分化的COPD培養物不處理,或藉由在供應至培養物基底側的培養基中包括處理來用1 µg/ml抗IL-33(33_640087-7B)、1 µg/ml NIP228(IgG1同種型對照)、10 µg/ml mNIP228、10 µg/ml抗ST2、1 µg/ml抗RAGE或1 µg/ml抗EGFR處理7天。每個週一、週三和週五進行一次培養基更換(含有相關處理)。
[表6]
23. ALI 培養物中杯狀細胞的 FACS 分析
| 抗體 | 識別符 |
| hIgG1 | NIP228_ SP14-266 |
| 抗IL-33(33_640087-7B) | SP15-124 |
| mIgG1 | mNIP228_ SP14-108 |
| 抗ST2 | Ab1440361 |
| 抗RAGE | M4F4 |
| 抗EGFR | LA1(默克公司(Merk),05-101) |
處理7天後(表6),藉由流動式細胞測量術分析了6.5 mm插入物上4週齡的正常對照或COPD ALI培養物。將200 µl 37°C PBS添加到每個Transwell的頂端區(Transwell表面),並放置在孵育箱中30 min。將頂端洗液儲存在-80°C下以進行黏蛋白分析。將150 µl胰蛋白酶(龍沙公司胰蛋白酶繼代培養包-#CC-5034)添加到頂端和基底外側(在Transwell下方)隔室中。將Transwell放回孵育箱後維持30 min。藉由輕輕地上下吸取胰蛋白酶來分離ALI。將150 µl胰蛋白酶中和溶液(龍沙公司胰蛋白酶繼代培養包-#CC-5034)添加到每個頂腔並混合。將細胞懸浮液移至U形90孔板,計數細胞並在4°C下以1200 RPM離心5 min。去除胰蛋白酶/TNS,並向每個孔中添加200 µl活死染料(eBioscience™可固定活力染料eFluor™780賽默公司65-0865-14,在PBS中1 : 2000稀釋)。將細胞再懸浮並在黑暗中在冰上孵育10 min。將該板在4°C下以1200 RPM離心5 min,去除活死染料,並向每個孔中添加200 µl PBS。將板在4°C下以1200 RPM離心5 min,去除PBS,並用200 µl固定/滲透溶液(賽默公司00-5123和00-5223)替換。將板在黑暗中在冰上孵育40 min。將板在4°C下以1200 RPM離心5 min,並去除溶液。將細胞再懸浮於300 µl 1x滲透溶液(賽默公司00-8333)中。將來自每個孔的5e4個細胞添加到在4°C下以1200 RPM離心5 min的新96孔U底板中,並將細胞再懸浮於50 µl 1x滲透溶液中。50 µl抗體染色混合物(1 : 400的抗Muc5AC和1 : 800的抗Muc5B)或相同稀釋度的同種型染色混合物。將板在黑暗中在冰上孵育30 min。將板在4°C下以1200 RPM離心5 min,並去除溶液。用PBS洗滌細胞,離心,然後再懸浮於150 µl PBS中。然後在BD FACSymphony™上採集數據,並使用FlowJo軟體分析。
[表7]
24. ALI 培養物的 qPCR/ 批量 RNA 定序分析。
| 名稱 | 識別符 | 供應商 | 螢光團 | 細胞標記 |
| 抗Muc5AC | ab3649 | 艾博抗公司 | 偶聯至AF488(Expedeon,332-0005) | 杯狀 |
| 抗Muc5B | ab105460 | 艾博抗公司 | 偶聯至AF647(Expedeon,336-0005) | 杯狀 |
| AF488同種型 | 400109 | 生物傳奇公司(BioLegend) | FITC | 同種型 |
| AF647同種型 | 400130 | 生物傳奇公司 | AF647 | 同種型 |
處理7天後(表6),使6.5 mm插入物上4週齡的正常對照或COPD ALI培養物裂解以進行RNA分析。首先將200 µl 37°C PBS添加到每個ALI頂端表面,並將板放回孵育箱後維持30 min。將頂端洗液儲存在-80°C下以進行黏蛋白分析。使用MagMAX™-96總RNA分離套組(賽默公司,AM1830)來裂解ALI培養物並提取RNA。然後使用High-Capacity RNA-to-cDNA™套組(賽默公司,4388950)使用RNA合成cDNA。從而將9 µl的每種RNA樣本與10 µl的2X RT緩衝劑混合物和1 µl的20X RT酶混合物在PCR管(賽默公司,AM12230)中孵育,置於賽默循環儀(thermo cycler)上,並且在37°C下孵育60分鐘。藉由加熱至95°C後維持5分鐘並保持在4°C下來終止反應。向含有20 µl cDNA的每個管添加60 µl無核酸酶的水(賽默公司,750024)。為進行RT-qPCR,將4 µl cDNA與5 µl TaqMan快速高級預混液(賽默公司,4444557)、0.5 µl Muc5AC FAM探針(賽默公司,Hs01365616_m1)和0.5 µl GAPDH VIC探針(賽默公司,Hs02786624_g1)一起添加到帶有條碼的MicroAmp™ EnduraPlate™光學384孔透明反應板(賽默公司,4483273)中。將板密封並短暫離心,隨後使用QuantStudio™ 7 Flex即時PCR系統(賽默公司)進行分析。然後藉由根據未處理的正常對照將數據歸一化來計算Δ-Δ-ct。
oxIL-33而非還原型IL-33使杯狀細胞數量增加,特別是MUC5AC+杯狀細胞亞群(圖19A至19C)。相應地,如藉由qPCR所判斷,用oxIL-33處理後,MUC5AC
mRNA的拷貝增加(圖19D)。25. IHC 三重染色(基底、杯狀和纖毛)和定量
接下來,評價了來自ALI免疫組織化學的定量圖像分析。如第22節:氣道基底細胞的氣液介面(ALI)培養中所述產生來自COPD供體的ALI培養物並處理。將ALI上皮培養物在10%中性緩衝福馬林(formalin)中固定24小時,並包埋在石蠟中。將石蠟切片(4 um)封固在帶正電荷的載玻片上,並藉由連續的3重發色測定在Ventana Discovery Ultra上染色。用細胞調節劑1(Ultra CC1)(目錄號5424569001,羅氏公司)進行抗原修復,並用Discovery抑制劑(目錄號7017944001,羅氏公司)阻斷內源過氧化物酶12 min。施加抗p63(殖株4A4)(目錄號790-4509,瑞士巴塞爾市羅氏公司)24 min,並用抗小鼠HQ(12 min)(目錄號7017782001,羅氏公司)和抗HQ HRP(12 min)(目錄號7017936001,羅氏公司)視覺化,並與Discovery Purple套組(目錄號07053983001,羅氏公司)一起孵育12 min。藉由抗體變性步驟(92°C,24 min),用細胞調節劑2(Ultra CC2)(目錄號5424542001,羅氏公司),然後用以Dako抗體稀釋劑(目錄號S3022)稀釋的抗微管蛋白(目錄號ab24610,英國劍橋市艾博抗公司)(載玻片上的濃度為0.003 µg/ml)處理載玻片16 min,用小鼠OmniMap-HRP(目錄號5269652001,羅氏公司)檢測(8 min),並用Discovery Teal HRP套組(目錄號82544338001,羅氏公司)視覺化。將載玻片用CC2再進行抗體變性,然後施加1.1 µg/ml(分配器濃度)兔抗黏蛋白5AC和7 µg/ml(分配器濃度)兔抗黏蛋白5B(分別為目錄號ab198294和目錄號ab87376,艾博抗公司)的混合物20 min,並用抗兔NP(4 min)(目錄號7425317001,羅氏公司)、抗NP-AP(8 min)(目錄號7425325001,羅氏公司)視覺化,然後用Discovery Yellow套組(目錄號7698445001,羅氏公司)視覺化20 min。將染色的載玻片用蘇木精II(8 min)(目錄號5277965001,羅氏公司)和藍化試劑(4 min)(目錄號5266769001,羅氏公司)複染色,用洗碗洗滌劑沖洗,用分級的乙醇和二甲苯系列脫水,並用永久封片劑封固。
在HALO v3.1(Indica Labs)中對IHC圖像進行分析,其中首先對該等圖像進行手動注釋,以排除焦點和組織損傷區。習得了一種隨機森林分類器,以識別上皮並將其與跨膜和載玻片背景分離。為進行纖毛區域定量,習得了另一種隨機森林分類器,以粗略檢測微管蛋白染色,隨後使用演算法區域定量(Area Quantificatio)v2.1.7進行精細檢測。為進行黏蛋白區域定量,直接使用區域定量v2.1.7來檢測染色。為進行基底(p63+)細胞計數,使用演算法CytoNuclear 2.0.9基於核染色對細胞進行分段,藉由計數p63陽性核進一步檢測基底細胞。所有定量方法均經過人識別驗證,並且準確性超過90%。
與先前的發現一致,oxIL-33深刻影響杯狀細胞(MUC5ac + b)的數量(圖20A和20B)。
總之,該等研究顯示oxIL-33在促進肺上皮內杯狀細胞分化方面發揮了作用。此表明長期暴露於ox-IL33的上皮向杯狀增生表型發展,這對肺功能產生負面影響。
26. 用阻斷劑逆轉COPD杯狀表型
COPD的一個重要標誌係由於杯狀細胞和黏液分泌的增加而引起的黏液過量(Gohy等人, 2019 Sci Rep [科學通訊] 9: 17963)。為了研究氧化型IL-33是否可以在杯狀COPD表型中發揮直接作用,建立了來自COPD供體的ALI培養物,其中讀數如第22-25節中所述。
在抗IL-33(33-640087_7B)、抗RAGE或抗EGFR中和抗體存在下培養COPD ALI。所有三種處理均使MUC5AC+杯狀細胞數量減少(圖21A至21D)。沒有處理影響ALI培養物的活力(圖21E),這證實了該處理現象不是假像或抗體毒性的結果。與先前的結果一致,抗ST2處理未引起杯狀細胞數量減少,進一步證明此係由IL-33,主要是ox-IL-33,通過oxIL-33-RAGE-EGFR路徑直接介導的疾病表型。在免疫組織化學分析中進一步證實了抗IL-33抗體(33-640087_7B)對COPD ALI的作用,其中在成對分析中,阻斷IL-33使杯狀細胞數量減少(圖22A和22B)。用抗IL-33抗體(33-640087_7B)處理後,COPD ALI培養物的上皮類似於健康上皮,如圖20A所示。
最後,使用ELISA測量了來自健康和COPD ALI培養物的頂端黏液中釋放的MUC5AC和MUC5B。為了定量從ALI培養物中釋放的黏蛋白,根據製造商的方案,藉由免疫測定(諾偉司(Novus)NBP2-76703)分析了頂端上清液中MUC5AC的水平。將樣本在樣本稀釋劑中1 : 2000稀釋,並從重組MUC5AC蛋白標準曲線外推出濃度。如圖23所示,與來自健康供體的ALI相比,來自COPD患者的ALI培養物釋放的MUC5AC水平增加(圖23A)。用外源oxIL-33處理使得健康ALI培養物的黏蛋白分泌增加(圖23B)。在COPD ALI供體中,黏蛋白水平的增加藉由用抗IL-33(33_640087_7B)阻斷而減少,該抗IL-33抑制從ALI培養物釋放的MUC5AC蛋白水平,但抗ST2或同種型mAb對照不抑制(圖23C)。
總體而言,本實例突出了氧化型IL-33在肺中上皮細胞分化失調中的作用。結果表明,在不受控制的情況下,氧化型IL-33可能是一些COPD表型中所見的杯狀細胞增生和黏液產生過多的原因。因此,用oxIL-33傳訊軸拮抗劑,諸如抗IL-33、抗RAGE或抗EGFR結合分子治療,可能藉由恢復正常的上皮生理學,例如藉由減少杯狀細胞數量和減少黏液產生過多,而對COPD患者可能具有巨大的治療益處。其他序列
除了表1中列出的序列,我們還提供以下其他CDR序列:
SEQ ID NO 37:SYAMS
SEQ ID NO 38:GISAIDQSTYYADSVKG
SEQ ID NO 39:QKFMQLWGGGLRYPFGY
SEQ ID NO 40:SGEGMGDKYAA
SEQ ID NO 41:RDTKRPS
SEQ ID NO 42:GVIQDNTGV
N末端His10/Avitag/因子Xa蛋白酶切割位點
SEQ ID NO 43:MHHHHHHHHHHAAGLNDIFEAQKIEWHEAAIEGR
IL-33-01
SEQ ID NO 44: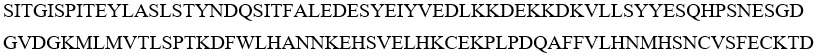
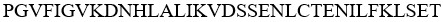 IL-33-16
SEQ ID NO 45:
IL-33-16
SEQ ID NO 45: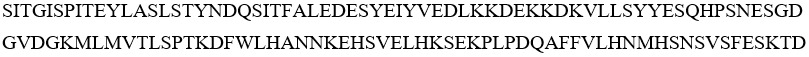
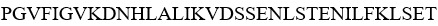 Avitag序列模體
SEQ ID NO 46:GLNDIFEAQKIEWHE
靶向RAGE外顯子3的gRNA載體
SEQ ID NO 47:TGAGGGGATTTTCCGGTGC
RAGE正向引物
SEQ ID NO 48:gttgcagcctcccaacttc
RAGE反向引物
SEQ ID NO 49:aatgaggccagtggaagtca
人ST2S(訊息肽加有底線)
SEQ ID NO 50:
Avitag序列模體
SEQ ID NO 46:GLNDIFEAQKIEWHE
靶向RAGE外顯子3的gRNA載體
SEQ ID NO 47:TGAGGGGATTTTCCGGTGC
RAGE正向引物
SEQ ID NO 48:gttgcagcctcccaacttc
RAGE反向引物
SEQ ID NO 49:aatgaggccagtggaagtca
人ST2S(訊息肽加有底線)
SEQ ID NO 50: 人ST2S-huIgG1 Fc-His6(訊息肽加有底線)
SEQ ID NO 51:
人ST2S-huIgG1 Fc-His6(訊息肽加有底線)
SEQ ID NO 51: SEQ ID NO 52:
His10/Avitag人ASGPR ECD(訊息肽加有底線,並且標籤加有雙底線)
SEQ ID NO 52:
His10/Avitag人ASGPR ECD(訊息肽加有底線,並且標籤加有雙底線)
無


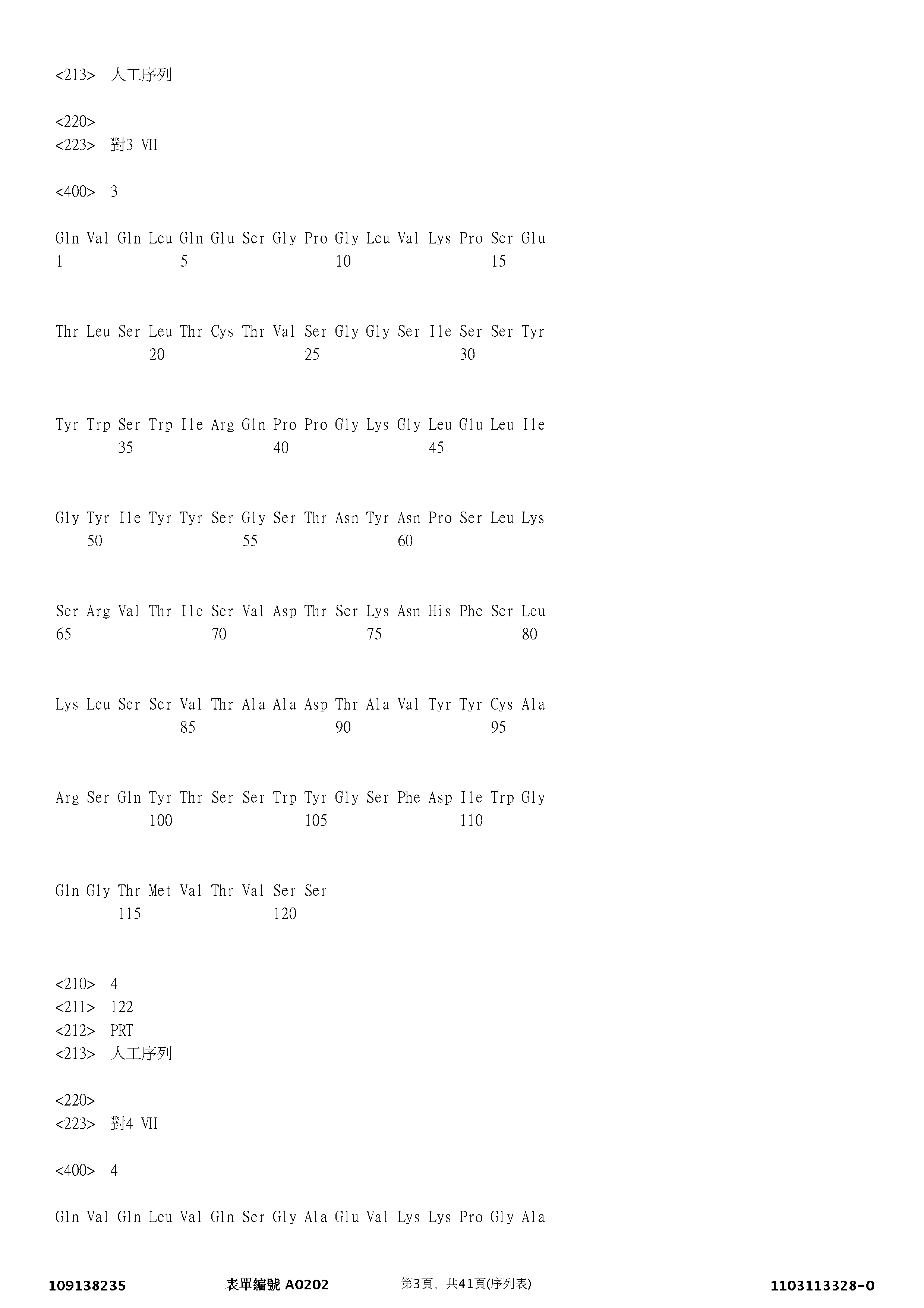

















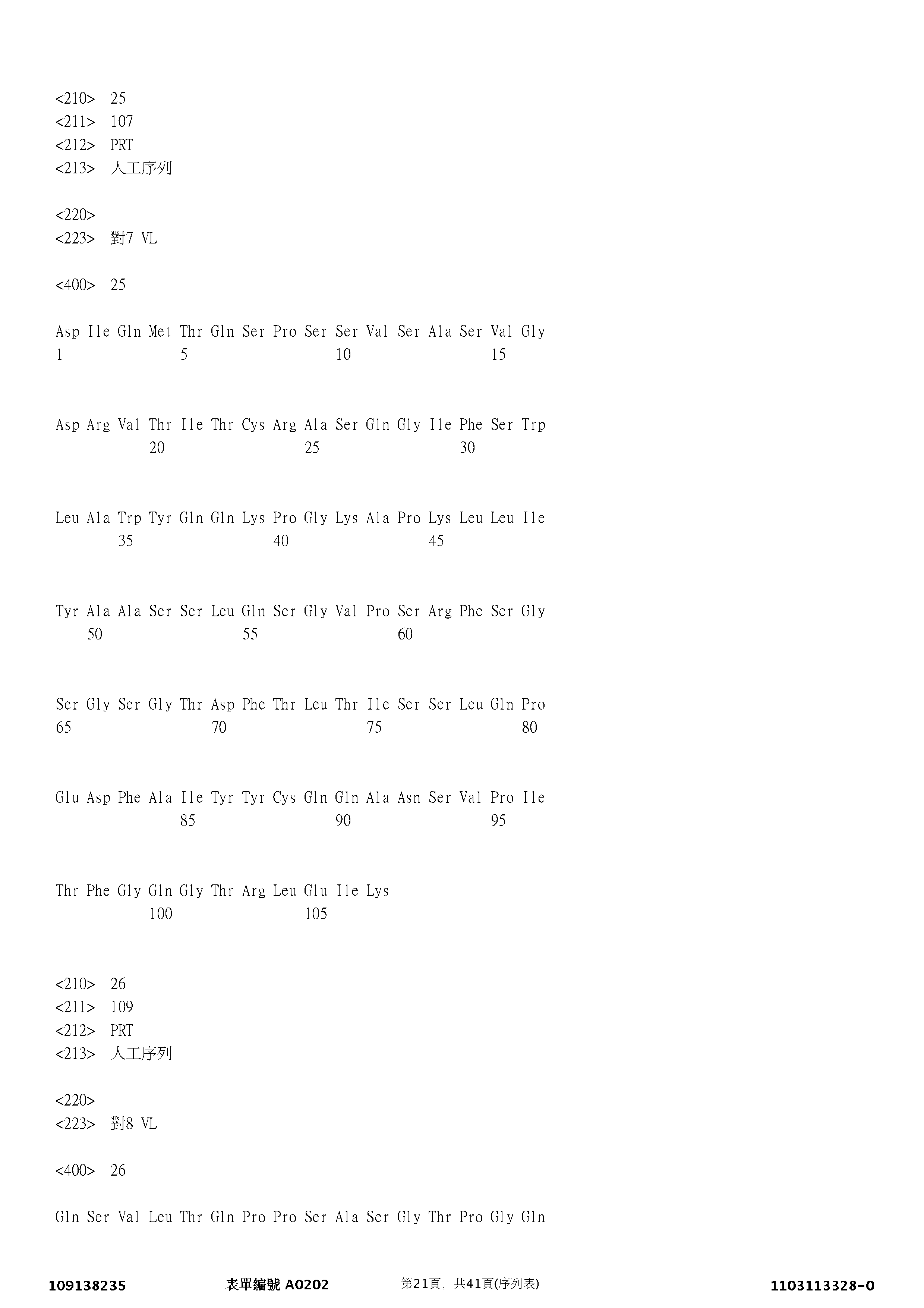














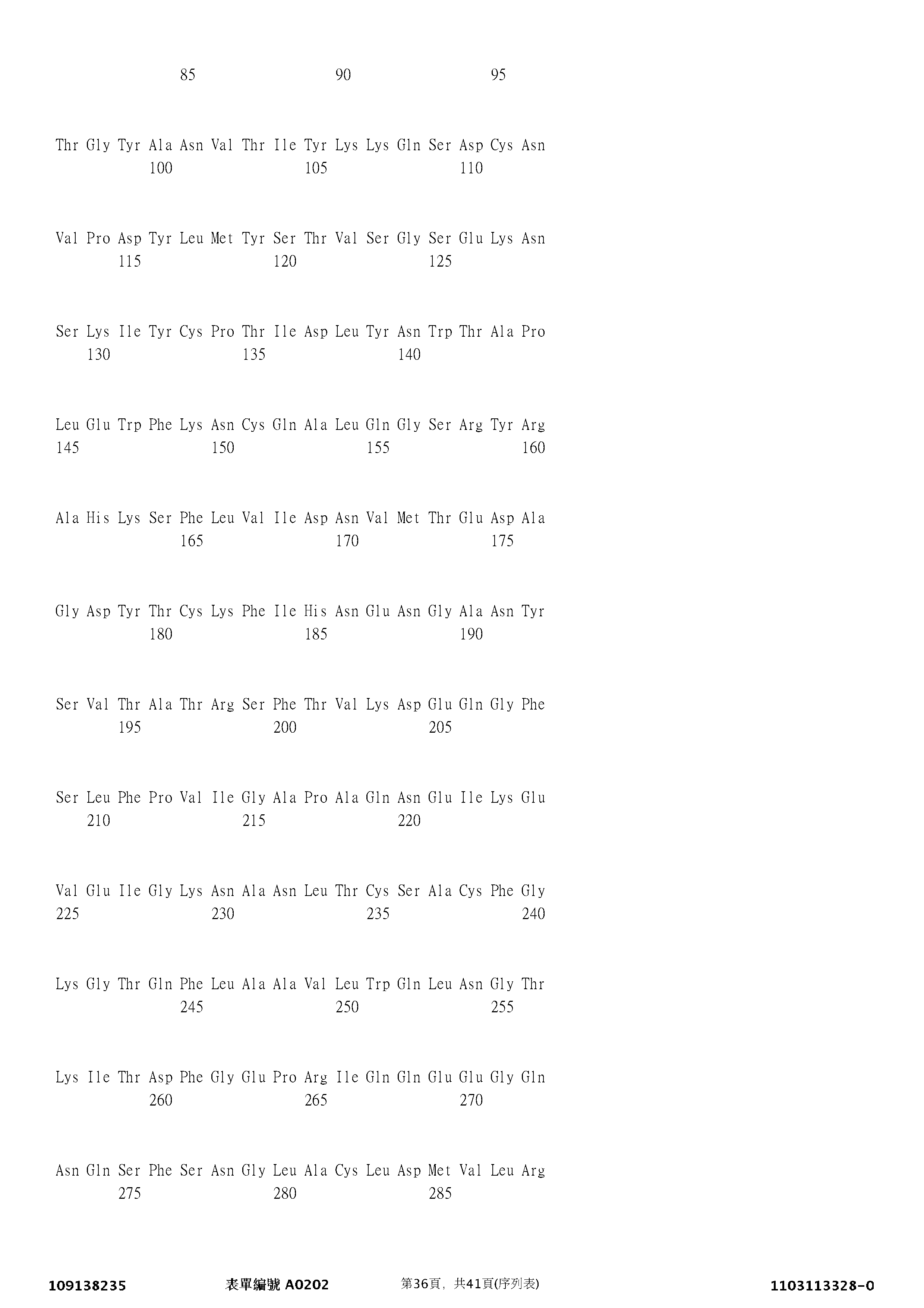





Claims (89)
- 一種IL-33拮抗劑,該IL-33拮抗劑用於藉由調節或抑制RAGE-EGFR介導的作用來預防或治療上皮生理學異常。
- 用於如請求項1所述使用的IL-33拮抗劑,其中該上皮生理學異常是黏膜纖毛生理學異常,較佳的是呼吸上皮的黏膜纖毛生理學異常。
- 用於如請求項2所述使用的IL-33拮抗劑,其中黏膜纖毛生理學異常選自:黏液產生異常;杯狀細胞分化異常;杯狀細胞增殖異常;上皮厚度異常;黏液清除異常;和/或黏液組成異常。
- 用於如請求項3所述使用的IL-33拮抗劑,其中黏液產生異常包括MUC5AC產生異常;和/或其中杯狀細胞分化異常包括MUC5AC+杯狀細胞分化異常;和/或其中杯狀細胞增殖異常包括MUC5AC+杯狀細胞增殖異常;和/或其中該上皮厚度異常包括該上皮的總組織區域中MUC5AC+ 杯狀細胞的量異常。
- 用於如請求項2、3或4所述使用的IL-33拮抗劑,其中黏膜纖毛生理學異常包括:黏液產生增加;杯狀細胞分化增加;杯狀細胞增殖增加;上皮厚度增加;和/或黏液清除減少。
- 用於如請求項5所述使用的IL-33拮抗劑,其中黏液產生增加包括MUC5AC產生增加;和/或其中杯狀細胞分化增加包括MUC5AC+杯狀細胞分化增加;和/或其中杯狀細胞增殖增加包括MUC5AC+杯狀細胞增殖增加;和/或其中該上皮厚度增加包括該上皮的總組織區域中MUC5AC+ 杯狀細胞的量增加。
- 用於如請求項3所述使用的IL-33拮抗劑,其中黏液組成異常包括黏液中含有的不同黏液化合物的比率增加或減少;一種或多種黏液化合物增加或減少;和/或黏液濃度或厚度增加或減少。
- 用於如請求項7所述使用的IL-33拮抗劑,其中黏液組成異常包括MUC5AC : MUC5B的比率增加;和/或其中黏液組成異常包括黏液中含有的MUC5AC增加;和/或其中黏液組成異常包括黏液厚度增加。
- 用於如請求項1所述使用的IL-33拮抗劑,其中該上皮生理學異常是上皮重塑異常。
- 用於根據任何前述請求項所述使用的IL-33拮抗劑,其中該上皮生理學異常是呼吸道的上皮生理學異常,較佳的是呼吸道的黏膜纖毛生理學異常。
- 用於如請求項10所述使用的IL-33拮抗劑,其中該呼吸道係下呼吸道,較佳的是支氣管。
- 一種IL-33拮抗劑,該IL-33拮抗劑用於預防或治療EGFR介導的疾病。
- 用於如請求項12所述使用的IL-33拮抗劑,其中該EGFR介導的疾病係RAGE-EGFR介導的疾病。
- 用於如請求項12或13所述使用的IL-33拮抗劑,其中該EGFR介導的疾病的特徵為EGFR活性異常。
- 用於如請求項12-14中任一項所述使用的IL-33拮抗劑,其中該EGFR介導的疾病的特徵為上皮生理學異常。
- 一種IL-33拮抗劑,該IL-33拮抗劑用於藉由改進上皮生理學來預防或治療疾病。
- 一種IL-33拮抗劑,該IL-33拮抗劑用於藉由抑制EGFR介導的作用來預防或治療疾病。
- 用於如請求項17所述使用的IL-33拮抗劑,其中該EGFR介導的作用係EGFR傳訊。
- 用於如請求項17或18所述使用的IL-33拮抗劑,其中該EGFR介導的作用係RAGE-EGFR介導的作用。
- 用於如請求項17-19中任一項所述使用的IL-33拮抗劑,其中該EGFR介導的作用係RAGE-EGFR介導的傳訊。
- 用於如請求項16-20中任一項所述使用的IL-33拮抗劑,其中該疾病係呼吸疾病。
- 用於如請求項21所述使用的IL-33拮抗劑,其中該呼吸疾病的特徵為上皮生理學異常和/或EGFR活性異常。
- 用於如請求項21或22所述使用的IL-33拮抗劑,其中該呼吸疾病係下呼吸道疾病,較佳的是支氣管的呼吸疾病。
- 用於如請求項21-23中任一項所述使用的IL-33拮抗劑,其中該呼吸疾病選自:COPD;支氣管炎;肺氣腫;支氣管擴張,諸如CF-支氣管擴張或-CF-支氣管擴張;氣喘;或氣喘與COPD重疊(ACO)。
- 用於如請求項21-24中任一項所述使用的IL-33拮抗劑,其中該呼吸疾病係COPD,較佳的是支氣管炎性COPD。
- 用於如請求項21-24中任一項所述使用的IL-33拮抗劑,其中該呼吸疾病係氣喘,較佳的是支氣管炎性氣喘。
- 用於如請求項16-26中任一項所述使用的IL-33拮抗劑,其中該預防或治療改進黏液清除。
- 用於如請求項16-27中任一項所述使用的IL-33拮抗劑,其中該預防或治療抑制或減少黏液產生異常。
- 用於如請求項28所述使用的IL-33拮抗劑,其中該預防或治療抑制或減少MUC5AC產生。
- 用於如請求項16-29中任一項所述使用的IL-33拮抗劑,其中該預防或治療抑制黏液組成異常。
- 用於如請求項30所述使用的IL-33拮抗劑,其中該預防或治療抑制或減少MUC5AC : MUC5B的比率;和/或其中該預防或治療抑制或減少黏液中的MUC5AC;和/或其中該預防或治療減少黏液的厚度。
- 用於如請求項16-31中任一項所述使用的IL-33拮抗劑,其中該預防或治療抑制上皮重塑異常。
- 用於如請求項16-32中任一項所述使用的IL-33拮抗劑,其中該預防或治療抑制杯狀細胞分化或增殖異常。
- 用於如請求項33所述使用的IL-33拮抗劑,其中該杯狀細胞分化或增殖異常是MUC5AC+ 杯狀細胞分化或增殖異常。
- 用於如請求項16-34中任一項所述使用的IL-33拮抗劑,其中該預防或治療減少呼吸上皮的厚度。
- 用於如請求項35所述使用的IL-33拮抗劑,其中該預防或治療減少該上皮的總組織區域中MUC5AC+ 杯狀細胞的量。
- 用於根據任何前述請求項所述使用的IL-33拮抗劑,其中該IL-33拮抗劑抑制氧化型IL-33的活性。
- 用於根據任何前述請求項所述使用的IL-33拮抗劑,其中該IL-33拮抗劑阻止氧化型IL-33與RAGE結合,從而抑制RAGE-EGFR傳訊。
- 用於根據任何前述請求項所述使用的IL-33拮抗劑,其中該IL-33拮抗劑下調或抑制RAGE-EGFR依賴性傳訊和/或RAGE-EGFR介導的作用。
- 用於根據任何前述請求項所述使用的IL-33拮抗劑,其中該IL-33拮抗劑係與IL-33結合,較佳的是與還原型IL-33或氧化型IL-33結合,較佳的是與還原型IL-33結合的結合分子或其片段。
- 用於根據任何前述請求項所述使用的IL-33拮抗劑,其中該IL-33拮抗劑係抗體或其抗原結合片段,較佳的是抗IL-33抗體或其抗原結合片段,較佳的是抗還原型IL-33抗體或其抗原結合片段。
- 用於根據任何前述請求項所述使用的IL-33拮抗劑,其中該IL-33拮抗劑係包含選自表1的可變重鏈結構域(VH)和可變輕鏈結構域(VL)對的互補決定區(CDR)的結合分子。
- 用於根據任何前述請求項所述使用的IL-33拮抗劑,其中該IL-33拮抗劑係包含選自表1的可變重鏈結構域(VH)和可變輕鏈結構域(VL)對的結合分子。
- 用於根據任何前述請求項所述使用的IL-33拮抗劑,其中該IL-33拮抗劑係如下結合分子,該結合分子包含具有SEQ ID NO: 37的序列的VHCDR1、具有SEQ ID NO: 38的序列的VHCDR2、具有SEQ ID NO: 39的序列的VHCDR3、具有SEQ ID NO: 40的序列的VLCDR1、具有SEQ ID NO: 41的序列的VLCDR2和具有SEQ ID NO: 42的序列的VLCDR3。
- 一種用於預防或治療受試者的上皮生理學異常之方法,該方法藉由投與治療有效量的IL-33拮抗劑以調節或抑制RAGE-EGFR介導的作用來進行。
- 如請求項45所述之方法,其中該上皮生理學異常是黏膜纖毛生理學異常,較佳的是呼吸上皮的黏膜纖毛生理學異常。
- 如請求項46所述之方法,其中黏膜纖毛生理學異常選自:黏液產生異常;杯狀細胞分化異常;杯狀細胞增殖異常;上皮厚度異常;黏液清除異常;和/或黏液組成異常。
- 如請求項47所述之方法,其中黏液產生異常包括MUC5AC產生異常;和/或其中杯狀細胞分化異常包括MUC5AC+杯狀細胞分化異常;和/或其中杯狀細胞增殖異常包括MUC5AC+杯狀細胞增殖異常;和/或其中該上皮厚度異常包括該上皮的總組織區域中MUC5AC+ 杯狀細胞的量異常。
- 如請求項47或48所述之方法,其中黏膜纖毛生理學異常包括:黏液產生增加;杯狀細胞分化增加;杯狀細胞增殖增加;上皮厚度增加;和/或黏液清除減少。
- 如請求項49所述之方法,其中黏液產生增加包括MUC5AC產生增加;和/或其中杯狀細胞分化增加包括MUC5AC+杯狀細胞分化增加;和/或其中杯狀細胞增殖增加包括MUC5AC+杯狀細胞增殖增加;和/或其中該上皮厚度增加包括該上皮的總組織區域中MUC5AC+ 杯狀細胞的量增加。
- 如請求項47所述之方法,其中黏液組成異常包括黏液中含有的不同黏液化合物的比率增加或減少;一種或多種黏液化合物增加或減少;和/或黏液濃度或厚度增加或減少。
- 如請求項51所述之方法,其中黏液組成異常包括MUC5AC : MUC5B的比率增加;和/或其中黏液組成異常包括黏液中含有的MUC5AC增加;和/或其中黏液組成異常包括黏液厚度增加。
- 如請求項45所述之方法,其中該上皮生理學異常是上皮重塑異常。
- 如請求項45-53中任一項所述之方法,其中該上皮生理學異常是呼吸道的上皮生理學異常,較佳的是呼吸道的黏膜纖毛生理學異常。
- 如請求項54所述之方法,其中該呼吸道係下呼吸道,較佳的是支氣管。
- 一種用於預防或治療EGFR介導的疾病之方法,該方法藉由投與治療有效量的IL-33拮抗劑來進行。
- 如請求項56所述之方法,其中該EGFR介導的疾病係RAGE-EGFR介導的疾病。
- 如請求項56或57所述之方法,其中該EGFR介導的疾病的特徵為EGFR活性異常。
- 如請求項56-58中任一項所述之方法,其中該EGFR介導的疾病的特徵為上皮生理學異常。
- 一種用於預防或治療疾病之方法,該方法藉由投與治療有效量的IL-33拮抗劑以改進上皮生理學來進行。
- 一種用於預防或治療疾病之方法,該方法藉由投與治療有效量的IL-33拮抗劑以抑制EGFR介導的作用來進行。
- 如請求項61所述之方法,其中該EGFR介導的作用係EGFR傳訊。
- 如請求項61或62所述之方法,其中該EGFR介導的作用係RAGE-EGFR介導的作用。
- 如請求項61-63中任一項所述之方法,其中該EGFR介導的作用係RAGE-EGFR介導的傳訊。
- 如請求項60-64中任一項所述之方法,其中該疾病係呼吸疾病。
- 如請求項65所述之方法,其中該呼吸疾病的特徵為上皮生理學異常和/或EGFR活性異常。
- 如請求項65或66所述之方法,其中該呼吸疾病係下呼吸道疾病,較佳的是支氣管的呼吸疾病。
- 如請求項65-67中任一項所述之方法,其中該呼吸疾病選自:COPD;支氣管炎;肺氣腫;支氣管擴張,諸如CF-支氣管擴張或-CF-支氣管擴張;氣喘;或氣喘與COPD重疊(ACO)。
- 如請求項65-68中任一項所述之方法,其中該呼吸疾病係COPD,較佳的是支氣管炎性COPD。
- 如請求項65-69中任一項所述之方法,其中該呼吸疾病係氣喘,較佳的是支氣管炎性氣喘。
- 如請求項45-70中任一項所述之方法,其中該方法改進黏液清除。
- 如請求項45-71中任一項所述之方法,其中該方法抑制或減少黏液產生異常。
- 如請求項72所述之方法,其中該黏液產生異常是MUC5AC產生增加。
- 如請求項45-73中任一項所述之方法,其中該方法抑制黏液組成異常。
- 如請求項74所述之方法,其中該方法抑制或減少MUC5AC : MUC5B的比率;和/或其中該方法抑制或減少黏液中的MUC5AC;和/或其中該方法減少黏液的厚度。
- 如請求項45-75中任一項所述之方法,其中該方法抑制上皮重塑異常。
- 如請求項45-76中任一項所述之方法,其中該方法抑制或減少杯狀細胞分化或增殖異常。
- 如請求項77所述之方法,其中該方法抑制或減少MUC5AC+ 杯狀細胞分化或增殖異常。
- 如請求項45-78中任一項所述之方法,其中該方法減少呼吸該上皮的厚度。
- 如請求項79所述之方法,其中該方法減少呼吸上皮的總組織區域中MUC5AC+ 杯狀細胞的量。
- 如請求項45-80中任一項所述之方法,其中該IL-33拮抗劑抑制氧化型IL-33的活性。
- 如請求項45-81中任一項所述之方法,其中該IL-33拮抗劑阻止氧化型IL-33與RAGE結合,從而抑制RAGE-EGFR傳訊。
- 如請求項45-82中任一項所述之方法,其中該IL-33拮抗劑下調或抑制RAGE-EGFR依賴性傳訊和/或RAGE-EGFR介導的作用。
- 如請求項45-83中任一項所述之方法,其中該IL-33拮抗劑係與IL-33,較佳的是還原型IL-33或氧化型IL-33,較佳的是原型IL-33結合的結合分子或其片段。
- 如請求項45-84中任一項所述之方法,其中該IL-33拮抗劑係抗體或其抗原結合片段,較佳的是抗IL-33抗體或其抗原結合片段,較佳的是抗還原型IL-33抗體或其抗原結合片段。
- 如請求項45-85中任一項所述之方法,其中該IL-33拮抗劑係包含選自表1的可變重鏈結構域(VH)和可變輕鏈結構域(VL)對的互補決定區(CDR)的結合分子。
- 如請求項45-86中任一項所述之方法,其中該IL-33拮抗劑係包含選自表1的可變重鏈結構域(VH)和可變輕鏈結構域(VL)對的結合分子。
- 如請求項45-87中任一項所述之方法,其中該IL-33拮抗劑係如下結合分子,該結合分子包含具有SEQ ID NO: 37的序列的VHCDR1、具有SEQ ID NO: 38的序列的VHCDR2、具有SEQ ID NO: 39的序列的VHCDR3、具有SEQ ID NO: 40的序列的VLCDR1、具有SEQ ID NO: 41的序列的VLCDR2和具有SEQ ID NO: 42的序列的VLCDR3。
- 如請求項45-88中任一項所述之方法,或用於如請求項1-44中任一項所述使用的IL-33拮抗劑,其中該IL-33拮抗劑係抗IL33抗體或其抗原結合片段,該抗IL33抗體或其抗原結合片段包含SEQ ID NO: 1的序列的VH結構域和SEQ ID NO: 19的序列的VL結構域。
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP19206984 | 2019-11-04 | ||
| EP19206984.7 | 2019-11-04 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| TW202134261A true TW202134261A (zh) | 2021-09-16 |
Family
ID=68426347
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| TW109138235A TW202134261A (zh) | 2019-11-04 | 2020-11-03 | 使用il-33拮抗劑之方法 |
Country Status (10)
| Country | Link |
|---|---|
| US (1) | US20220380450A1 (zh) |
| EP (1) | EP4054711A1 (zh) |
| JP (2) | JP2023500492A (zh) |
| KR (1) | KR20220093334A (zh) |
| CN (1) | CN114901361A (zh) |
| AU (2) | AU2020379171B2 (zh) |
| CA (1) | CA3158323A1 (zh) |
| IL (1) | IL292443A (zh) |
| TW (1) | TW202134261A (zh) |
| WO (1) | WO2021089563A1 (zh) |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AU2022335719B2 (en) * | 2021-08-27 | 2024-11-28 | Medimmune Limited | Treatment of chronic obstructive pulmonary disease with an anti-interleukin-33 antibody |
| TW202402790A (zh) | 2022-03-25 | 2024-01-16 | 英商梅迪繆思有限公司 | 減少呼吸系統感染之方法 |
| TW202423972A (zh) | 2022-08-26 | 2024-06-16 | 英商梅迪繆思有限公司 | 使用抗介白素-33抗體的氣喘治療 |
Family Cites Families (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US5892019A (en) | 1987-07-15 | 1999-04-06 | The United States Of America, As Represented By The Department Of Health And Human Services | Production of a single-gene-encoded immunoglobulin |
| WO2008137552A2 (en) | 2007-05-02 | 2008-11-13 | Medimmune, Llc | Anti-rage antibodies and methods of use thereof |
| US9212227B2 (en) | 2012-04-30 | 2015-12-15 | Janssen Biotech, Inc. | ST2L antibody antagonists for the treatment of ST2L-mediated inflammatory pulmonary conditions |
| UY34813A (es) | 2012-05-18 | 2013-11-29 | Amgen Inc | Proteínas de unión a antígeno dirigidas contra el receptor st2 |
| JO3532B1 (ar) | 2013-03-13 | 2020-07-05 | Regeneron Pharma | الأجسام المضادة لمضاد انترلوكين-33 واستعمالاتها |
| PH12016500425B1 (en) * | 2013-09-05 | 2022-07-29 | Ab2 Bio Sa | Il-18 binding protein (il-18bp) in inflammatory diseases |
| MX381087B (es) | 2013-12-26 | 2025-03-12 | Mitsubishi Tanabe Pharma Corp | Anticuerpo monoclonal neutralizador de anti-il-33 humana. |
| EP3092253B1 (en) | 2014-01-10 | 2021-03-17 | AnaptysBio, Inc. | Antibodies directed against interleukin-33 (il-33) |
| BR112017009728A2 (pt) | 2014-11-10 | 2018-02-06 | Genentech, Inc. | anticorpos isolados, ácido nucleico isolado, vetor, célula hospedeira, método para produzir o anticorpo, composição, usos de um anticorpo e métodos de tratamento de um distúrbio |
| MX387295B (es) * | 2015-03-31 | 2025-03-18 | Medimmune Ltd | Nueva forma de interleucina-33 (il33), formas mutadas de interleucina-33 (il33), anticuerpos, ensayos y métodos para usar los mismos. |
| JP2019516362A (ja) | 2016-04-27 | 2019-06-20 | ファイザー・インク | 抗il−33抗体、その組成物、方法および使用 |
| JOP20190093A1 (ar) | 2016-10-28 | 2019-04-25 | Lilly Co Eli | أجسام مضادة لـ il-33 واستخداماتها |
| WO2018204598A1 (en) | 2017-05-03 | 2018-11-08 | The University Of North Carolina At Chapel Hill | Methods for diagnosing or predicting chronic bronchitis |
-
2020
- 2020-11-03 TW TW109138235A patent/TW202134261A/zh unknown
- 2020-11-03 EP EP20797530.1A patent/EP4054711A1/en active Pending
- 2020-11-03 KR KR1020227018052A patent/KR20220093334A/ko active Pending
- 2020-11-03 AU AU2020379171A patent/AU2020379171B2/en active Active
- 2020-11-03 CN CN202080076133.0A patent/CN114901361A/zh active Pending
- 2020-11-03 CA CA3158323A patent/CA3158323A1/en active Pending
- 2020-11-03 WO PCT/EP2020/080841 patent/WO2021089563A1/en not_active Ceased
- 2020-11-03 IL IL292443A patent/IL292443A/en unknown
- 2020-11-03 US US17/755,605 patent/US20220380450A1/en active Pending
- 2020-11-03 JP JP2022525520A patent/JP2023500492A/ja active Pending
-
2025
- 2025-08-08 AU AU2025213676A patent/AU2025213676A1/en active Pending
- 2025-10-16 JP JP2025174754A patent/JP2026012782A/ja active Pending
Also Published As
| Publication number | Publication date |
|---|---|
| AU2020379171A1 (en) | 2022-06-09 |
| AU2020379171B2 (en) | 2025-05-08 |
| JP2026012782A (ja) | 2026-01-27 |
| KR20220093334A (ko) | 2022-07-05 |
| IL292443A (en) | 2022-06-01 |
| CN114901361A (zh) | 2022-08-12 |
| CA3158323A1 (en) | 2021-05-14 |
| AU2025213676A1 (en) | 2025-08-28 |
| US20220380450A1 (en) | 2022-12-01 |
| WO2021089563A1 (en) | 2021-05-14 |
| EP4054711A1 (en) | 2022-09-14 |
| JP2023500492A (ja) | 2023-01-06 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CA3176518A1 (en) | Methods and compositions for treating virus-associated inflammation | |
| US20250042986A1 (en) | Stem cell factor inhibitor | |
| JP2026012782A (ja) | Il-33アンタゴニストの使用方法 | |
| JP2017165652A (ja) | 新規抗ヒトox40リガンド抗体、及びこれを含む抗インフルエンザ薬 | |
| US20220153826A1 (en) | Methods and compositions for treating virus-associated inflammation | |
| US20220363748A1 (en) | Anti il-33 therapeutic agent for treating renal disorders | |
| HK40016017A (zh) | 用於调节肺中炎性体活性和炎症的方法 |