CN101333516A - 嵌合体动物及其制备方法 - Google Patents
嵌合体动物及其制备方法 Download PDFInfo
- Publication number
- CN101333516A CN101333516A CNA2008100099774A CN200810009977A CN101333516A CN 101333516 A CN101333516 A CN 101333516A CN A2008100099774 A CNA2008100099774 A CN A2008100099774A CN 200810009977 A CN200810009977 A CN 200810009977A CN 101333516 A CN101333516 A CN 101333516A
- Authority
- CN
- China
- Prior art keywords
- human
- cells
- chromosome
- mouse
- gene
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images


























Classifications
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K67/00—Rearing or breeding animals, not otherwise provided for; New or modified breeds of animals
- A01K67/027—New or modified breeds of vertebrates
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K67/00—Rearing or breeding animals, not otherwise provided for; New or modified breeds of animals
- A01K67/027—New or modified breeds of vertebrates
- A01K67/0275—Genetically modified vertebrates, e.g. transgenic
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K16/00—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies
- C07K16/18—Immunoglobulins [IGs], e.g. monoclonal or polyclonal antibodies against material from animals or humans
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/8509—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells for producing genetically modified animals, e.g. transgenic
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/90—Stable introduction of foreign DNA into chromosome
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2217/00—Genetically modified animals
- A01K2217/05—Animals comprising random inserted nucleic acids (transgenic)
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2217/00—Genetically modified animals
- A01K2217/07—Animals genetically altered by homologous recombination
- A01K2217/075—Animals genetically altered by homologous recombination inducing loss of function, i.e. knock out
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2227/00—Animals characterised by species
- A01K2227/10—Mammal
- A01K2227/105—Murine
-
- A—HUMAN NECESSITIES
- A01—AGRICULTURE; FORESTRY; ANIMAL HUSBANDRY; HUNTING; TRAPPING; FISHING
- A01K—ANIMAL HUSBANDRY; AVICULTURE; APICULTURE; PISCICULTURE; FISHING; REARING OR BREEDING ANIMALS, NOT OTHERWISE PROVIDED FOR; NEW BREEDS OF ANIMALS
- A01K2267/00—Animals characterised by purpose
- A01K2267/01—Animal expressing industrially exogenous proteins
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2517/00—Cells related to new breeds of animals
- C12N2517/02—Cells from transgenic animals
Landscapes
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Biotechnology (AREA)
- General Health & Medical Sciences (AREA)
- Molecular Biology (AREA)
- Biophysics (AREA)
- General Engineering & Computer Science (AREA)
- Biochemistry (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biomedical Technology (AREA)
- Environmental Sciences (AREA)
- Animal Husbandry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Physics & Mathematics (AREA)
- Microbiology (AREA)
- Plant Pathology (AREA)
- Animal Behavior & Ethology (AREA)
- Biodiversity & Conservation Biology (AREA)
- Veterinary Medicine (AREA)
- Immunology (AREA)
- Medicinal Chemistry (AREA)
- Mycology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Peptides Or Proteins (AREA)
Abstract
一种制备嵌合体非人动物的方法,包括制备保持单一或多个外源染色体或其片段的微细胞,通过与该微细胞融合将该外源染色体或其片段转移到分化多能性细胞;一种嵌合体非人动物,可通过上述方法进行构建,或其后代;来源于它们的组织和细胞;一种应用这些组织和细胞的方法;含有单个或多个外源染色体或其片段和持续分化多能性细胞;一种构建该细胞的方法;一种应用该细胞的方法。
Description
本申请是申请日为1996年8月29日,申请号为96197787.6的、发明名称和本发明相同的发明专利申请的分案申请。
技术领域
本发明涉及嵌合体非人动物,其制备方法及使用方法。若使用本发明的嵌合体非人动物,则可以在动物体内保持,表达1Mb(百万碱基对)以上的巨大DNA片段,这在目前尚不能做到。因此,利用本发明可做到:
·可制作保持、表达编码生物活性物质基因的全长,如人抗体基因全长的动物。从该动物得到的生物活性物质具有医药品的利用价值。
·可对动物中的人巨大基因(组织相容性抗原、肌营养不良白蛋白等)进行功能分析。
·可用于制备人显性遗传性疾病和染色体异常症的动物模型。
技术背景
在动物中表达外来基因的技术,即转基因动物的制作技术,不仅有利于获得有关基因在生物体内功能的信息,而且可用于检测调控基因表达的DNA序列(例,Magram等,自然,315:338,1985),建立人类疾病的动物模型(山村等,疾病小鼠模型手册,中山书店,1994),家畜的繁殖(例,Muller等,Experientia,47:923,1991),以及利用它生产有用的物质(例,Velander等,美国国家科学院院刊,89:12003,1992)。目前为止应用最多的是以小鼠作为转基因的对象。作为实验动物,已对小鼠进行了全面研究,且已建立了胚胎操作技术,所以小鼠是最适宜的哺乳动物。
已知外源基因转移到小鼠的方法大致有两种。1种是将DNA注入受精卵的原核(Gordon等,美国国家科学院院刊,77:7380,1980),另一种是将DNA转移到全能胚胎干细胞(以下称作ES细胞),制作嵌合体小鼠的方法(Takahashi等,发育,102:259,1988)。在后者的嵌合体小鼠中,仅仅在提供ES细胞的细胞和组织中保留转移基因,而在经ES细胞分化的生殖细胞而获得的后代所有细胞和组织中均保留转移基因。目前应用该技术已制备了大量的转基因小鼠。
但是,由于目前可转移DNA的大小受到限制,也大大限制了该技术的应用范围。这种限制依赖于可以被克隆的DNA的大小,目前为止转移最大DNA片段的1例是将大约670kb的DNA片段克隆到酵母人工染色体(YAC)中(Jakobovits等,自然,365:255,1993)。该实验是将含有YAC的酵母与ES细胞融合而进行的。YAC中可克隆大约2Mb的外源DNA(Den Dunnen等,人类分子基因,1:19,1992),但由于在出芽酵母细胞中,同源DNA序列间重组的发生率高,所以以完整形式保持具有很多重复序列的人的DNA片段是很困难的。事实上,在含有人基因组DNA的YAC文库中大约发生20~40%的重组(Green等,基因组,11:584,1991)。
另外偿试的一个方法是,在显微镜下切断由人的培养细胞而得到的中期染色体,然后将片段(估计10Mb以上)注入小鼠受精卵中(Richa等,科学,245:175,1989)。在所得到的小鼠中能检测到人特异的DNA序列(Alu序列),但不能证实有人基因的表达。此外,这里所应用的制备染色体的方法中,将染色体固定于载玻片时,由于使用了醋酸甲醇,不可避免地DNA出现小片段,所注入的DNA以完整形式存在的可能性很小。
无论如何,目前为止尚无将1Mb以上的外源连续DNA转移到小鼠进行表达的报道。
已知那些希望转移到小鼠、有益的且吸引人的人类基因,如抗体(例Cook等,Natune Genetics,7:162,1994)、T细胞受体(Hood等,冷泉港分析生物学研讨会,Vol.LVIII,339,1993)、组织相容性抗原(Carrol等,美国国家科学院院刊,84:8535,1987)、肌营养不良蛋白(DenDunnen等,同前),其编码区域的大小至少为1Mb。由于人类抗体是很重要的医药品,所以人们期望能够制作保持、表达人免疫球蛋白重链(~1.5Mb,Cook等,同前)、轻人类分子遗传学链κ(~3Mb,Zachau,基因,135:167,1993)、轻链λ(~1.5Mb,Frippiat等,4:983,1995)基因全长的小鼠,但目前该领域内的技术尚无法完成(Nikkei Biotec,1993,7.5.)。
人类显性遗传疾病、引起先天异常的染色体异常症(唐氏综合征等)的许多诱因基因尚不能克隆,只能利用染色体上大致位置的信息。例如,对中期染色体进行吉姆萨染色所得到的G带,通常在数Mb~10Mb大小。由此可发现某些诱因基因存在于特定的G带上,而将这些异常表型转移到小鼠的时候,有必要将诱因基因周围的染色体片段(数Mb以上)进行转移,这也是目前的技术达不到的。
因此,希望能开发一种能将长度超过1Mb限制的外源DNA导入到小鼠中并使其表达的技术。
应用目前的技术,可以将超过上述界限的DNA转移到动物的培养细胞中。这主要以染色体(人染色体的大小为50~300Mb)为介质进行。有一些关于将染色体移入细胞的方法的报道(例,McBride等,美国国家科学院院刊,70:1258,1973),其中微细胞法是只将1条所选染色体进行转移的最佳方法(Koi等,日本癌症研究杂志,80:413,1989)。叫做微细胞的构造体是用核膜和质膜包裹一至数条染色体。经抑制某种细胞中纺缍体形成的抑制剂诱导形成微细胞,分离它,与受体细胞融合,可转移少数(大多数为1条)染色体。用这种方法所得到的,只含有1条人类染色体的单染色体杂交细胞的文库,可用于已知基因的作图,说明未知抑癌基因、细胞老化基因所存在的染色体(例,Saxon等,EMBO J.,5:3461,1986)。此外,还可用γ-射线照射微细胞,将所得染色体片段的一部分进行转移(Koi等,科学(Science),260:361,1993)。如上所述,微细胞法是将1Mb以上的DNA转移到动物培养细胞中的一种适宜的方法。
随着ES细胞这一能稳定保持全能性细胞的发现(Evans等,自然(Nature),292:154,1981),从培养细胞到能制备小鼠的愿望成为现实。可以将外源基因、各种突变及目的基因重组造成的突变基因均可转移到ES细胞,可在小鼠水平进行大范围的基因修饰(例,Mansour等,自然(Nature),336:348,1988)。导入巨大DNA的意思是以前面所讲的克隆到YAC载体中的可能的外源DNA片段的大小为界限的。目前将较长的DNA导入培养细胞的染色体转移技术还不能用于小鼠的基因转移,相信这是难以完成的(村松等,转基因生物学,讲谈社Scientific,p143-,1989)。
理由如下。
将人的染色体转移到正常核型的受体细胞ES细胞是转移染色体异常。至今人们认为如果发生显微镜下可识别的染色体水平的基因异常,通常对小鼠的胚胎发生是致死性的(Gropp等,实验动物杂志(实验动物学,228:253,1983);相沢慎一,生物技术手册系列8,基因打靶,羊土社,1995)。
·我们所能得到的人染色体通常是有限增殖的正常纤维母细胞、或是癌细胞等分化的体细胞,将这样来源于体细胞的染色体转移到未分化的ES细胞的时候,会引起ES细胞的分化(Muller等,自然(Nature),311:438,1984)或老化(Sugawara,科学(Science),247:707,1990)。
·将体细胞来源的染色体转移到初期胚胎中,在胚胎的发育过程中能否起到与来源于生殖细胞的染色体一样的功能、确保各种组织、细胞中特异基因的表达,关于这一问题所进行的研究很少。二者最大的差异之一是关于染色体DNA的甲基化状态。甲基化伴随细胞的分化而有所不同,这表明甲基化对于组织特异性基因的表达起到主要作用(Ceder,细胞,53:3,1988)。例如,有这样的报道,在活化抗体基因对所进行的基本的位点特异性DNA重组反应中,将甲基化的底物转移到B细胞,若复制后也保持甲基化状态,就阻碍重组反应(Hsieh等,EMBO.J.,11:315,1992)。此外,与生物体内相比所建立的细胞株中,容易发生从头甲基化(Antequera等,细胞,62:503,1990)。基于目前的研究,不能简单地设想,纤维母细胞、人-鼠杂交细胞中高度甲基化的抗体基因在小鼠的B细胞中正常表达。
必需注意的是这里有III mensee等的两个相关报道(美国国家科学院院刊,75:1914,1978;美国国家科学院院刊,76:879,1979)。一个是人的肉瘤细胞与小鼠EC细胞融合,另一个是大鼠的肝癌细胞与小鼠EC细胞融合,用得到的融合株制作嵌合体小鼠。这些报道中,对实验结果指出了许多问题,其可靠性低(野口等,小鼠畸胎瘤,理工学社,5版,1987)。此外,尽管人们期望能早日探讨清楚,但是报道发表后的17年至今没有成功重复这些实验的报道。因此我们认为根据这些报道,在小鼠体内保持外源染色体,并表达该染色体上的基因是不可能的。
在目前的状况下,将染色体片段那样的巨大DNA转移到小鼠等动物体内并使之表达是很困难的,实际上,自上面III mensee等的报道以来没有对此问题进行研究。
因此,本发明的目的是提供嵌合体非人动物及其后代,以及其制作方法,其中该动物保持外源染色体或其片段,并能表达该染色体或其片段上的基因。
另外,本发明的目的是提供来源于上述嵌合体非人动物及其后代的组织和细胞。
本发明的另一目的是提供杂交瘤,该杂交瘤是上述嵌合体非人动物及其后代的细胞与骨髓瘤融合而形成。
本发明还有一目的是提供制备生物活性物质的方法,它是非人动物、其组织或细胞来表达外源染色体或其片段上基因的表达产物。
发明公开
为了解决上述问题,本发明人进行了多种研究,结果是用微细胞法成功地将来源于人正常纤维母细胞的染色体或其部分片段转移到ES细胞中,得到能稳定保持它们的克隆。并且他们从该ES细胞株得到嵌合体小鼠,该小鼠的正常组织中保持有人染色体,并且能表达包含人抗体重链基因的数个人基因。我们通过这一系列方法能够制备保持巨大DNA片段并表达该DNA片段上基因的动物,而这在目前是不可能的。
本发明的主旨如下:
(1)嵌合体非人动物的制备方法,其特征在于制备含有一个或几个外源染色体或含有其片段的微细胞,通过与该微细胞融合,将上述的单一或几个外源染色体或其片段导入具有分化多能性的细胞中。
(2)含有单一或几个外源染色体或其片段的、具有分化多能性的细胞的制备方法,其特征在于,制作含有一个或几个外源染色体或含有其片段的微细胞,通过与该细胞融合,将上述的单一或几个染色体或其片段移入具有分化多能性的细胞中。
(3)用上述(1)的方法能够制备的保持有单一或多个外源染色体或其片段,并表达该染色体或其片段上基因的嵌合体非人动物及其具有同一特性的后代。
(4)用(2)的方法能够制备的含有一个或几个外源染色体或其片段、具有分化多能性的细胞。
(5)(4)中具有分化多能性的细胞用于制备嵌合体非人动物。
(6)上述(3)中的嵌合体非人动物或其后代交配而得到的保持有一个或几个外源性染色体或其片段、并表达该染色体或其片段上基因的非人动物或其后代。
(7)来源于(3)的嵌合体非人动物或其后代或来源于(6)的非人动物或其后代的组织。
(8)一种细胞,其来源于(3)的嵌合体非人动物或其后代或来源于(6)的非人动物或其后代。
(9)上述(8)的细胞与骨髓瘤细胞融合而得到的杂交瘤。
(10)一种非人动物及其后代,其制备是将(3)的嵌合体非人动物或其后代或将(6)的非人动物或其后代与上述染色体或其片段上同一基因或相似基因缺陷的非人动物系交配而得到的。
(11)一种生物活性物质的制备方法,其特征在于在(3)的嵌合体非人动物或其后代、组织或细胞中或在(6)的非人动物或其后代、组织或细胞中表达一个或几个外源染色体或其片段上的基因,回收作为表达产物的生物活性物质。
(12)一种制备生物活性物质的方法,其特征在于,将(3)或将(6)的嵌合体非人动物或其后代与缺乏上述染色体或其片段上同一基因或相似基因的非人动物系交配,在所得到的子代动物、组织或细胞中表达上述一个或几个外源染色体或其片段上的基因,并回收作为表达产物的生物活性物质。
附图简述
图1表示保持有人2号染色体(片段)的A9细胞的分析结果(PCR分析)。
图2表示E14耐药株中保持有人的22号染色体(片段)(PCR分析)。
图3是一电泳的照片图,它表明在由于移入了22号染色体的ES细胞而来的嵌合体小鼠中保持有人的L 1序列(Southen分析)。
图4是一电泳的照片,它表明在导入了人22号染色体的嵌合体小鼠的脏器中保持有人染色体(PCR分析)。
图5是一电泳的照片,它表明了在导入了人22号染色体的嵌合体小鼠中,人基因表达的结果(RT-PCR)。
图6是一电泳的照片,它表明了在导入了人22号染色体的嵌合体小鼠的脏器中,人基因表达的结果(RT-PCR)。
图7表示在E14耐药株中保持有人的4号染色体(片段)(PCR)。
图8是一电泳的照片,它表示在导入了人4号染色体的E14细胞株中,对人L1序列的检测结果(Southern)。
图9是一电泳的照片,它表明在由导入了人4号染色体的ES细胞而来的嵌合体小鼠中,保持有人L1序列(Southern)。
图10表示在TT2耐药株中保持有人14号染色体(片段)(PCR)。
图11是一电泳的照片,它表明在由导入了人4号染色体的ES细胞而来的嵌合体小鼠的脏器中,保持有人染色体(PCR)。
图12表示尾部来源的纤维母细胞对G418抗性的试验结果。
图13表示经人血清白蛋白免疫的嵌合体小鼠血清中人IgM抗体的浓度(ELISA)。
图14表示经人血清白蛋白免疫的嵌合体小鼠血清中人IgG抗体的浓度(ELISA)。
图15表示产生人IgM的杂交瘤克隆H4B7的分析结果(ELISA)。
图16是一染色体形态的照片,它表明保持有人2号和14号染色体片段的小鼠ES细胞株(TT2细胞株PG15)的FISH分析结果。
图17表示在经人血清白蛋白免疫的嵌合体小鼠血清中抗人血清白蛋白的人IgG抗体的效价增加。
图18表示在经人血清白蛋白免疫的嵌合体小鼠血清中,抗人血清白蛋白的人Igκ抗体的效价增高。
图19是一电泳的照片,它表示在导入了人22号染色体的TT2细胞中,人L1序列的检测结果(Southern分析)。
图20表示在经人血清白蛋白免疫的嵌合体小鼠血清中,抗人血清白蛋白的人Igλ抗体的效价增高。
图21表明在导入了人2号染色体部分片段的嵌合体小鼠的后代中,保持有人2号染色体的部分片段(PCR)。
图22表明在导入了人14号染色体的嵌合体小鼠脾脏中,有细胞表面表达人μ链的细胞存在(流式细胞术分析)。
图23表明LoxP-pst NEO质粒DNA的构建。
图24表明小鼠抗体重链Cμ基因组DNA的结构。
图25表明小鼠抗体轻链κ基因组DNA的结构。
图26表明小鼠抗体重链靶向载体、Southern印迹法中应用的探针及在同源重组体中能够检测到的DNA片段的结构。
图27表明小鼠抗体轻链κ的靶向载体、Southern印迹法中应用的探针及在同源重组体中能够检测到的DNA片段的结构。
图28是一电泳的照片,它表明小鼠抗体重链同源重组体及由同源重组体而来的高浓度G418耐药株的Southern印迹分析结果。
实施本发明的最佳方式
保持有人染色体或其片段、并表达该染色体或其片段上的基因的小鼠,其制备如下:
(1)制备保持有标记的人染色体或其片段的染色体供体细胞
(2)用微细胞法将人染色体或其片段移入具有分化多能性的小鼠细胞中
(3)用上述小鼠细胞制作嵌合体小鼠,
(4)证实在嵌合体小鼠中保持有人染色体,并表达人基因。
这里,保持有人染色体或其片段、表达该染色体或其片段上的基因的非人动物是以小鼠为例的(此后,小鼠称作“导入人染色体的小鼠”)。
术语“人染色体”指自然形成的复合体,它由人细胞的核酸和蛋白质组成。正常的人类染色体有46条,23种(男性为24种),每条染色体约含50-300Mb的DNA。本发明中,独立染色体是指能稳定复制、分离的部分片段,并且也包括能异位到小鼠染色体上,稳定保持的部分片段。DNA的大小通常在1Mb以上,也有小于1Mb的情况。本发明的特征是以染色体为介质在小鼠内保持、表达人基因,而不用在大肠杆菌、酵母等中克隆或从细胞中提取DNA。
导入人染色体的小鼠指,在其全部或部分正常体细胞中保持有一个或几个人染色体或其片段。另外,指在其全部或部分体细胞中表达人染色体上的一个或几个人基因。
(1)制备保持有标记的人染色体或其片段的染色体供体细胞
所希望的染色体供体细胞是1)保持有标记物标记的人染色体,而该标志物可用于受体细胞的筛选2)不含除此之外的人染色体3)形成微细胞的能力高。
来源于人的任何细胞株、癌细胞和原代培养细胞均可用作提供人染色体的物质,但正常纤维母细胞较合适,因为它的染色体发生缺失,扩增等异常现象的可能性较小,且容易培养。
首先,对(1)来讲,人细胞可用表达耐药性(G418,嘌呤霉素,潮霉素,杀稻瘟素)等标记基因的载体进行转化。希望用于调节标记基因的启动子,不仅在人细胞中而且在小鼠ES细胞等受体细胞中也能很好地发挥作用。为了达到这一目的可应用与SV和增强子联结的单纯疱疹病毒胸苷激酶启动子(Katoh等,细胞结构功能(Cen Struct.Funct.,12:575,1987)、小鼠PGK-1启动子(Soriano等,细胞,64:693,1991)。用电穿孔法(石田等,细胞工学实验操作入门,讲谈社,1992)进行转化,将导入的标记基因随机插入23种、46条人染色体上,随后进行筛选,可得到人细胞转化体文库。
对于3),由于大多数的人正常细胞形成微细胞的能力较低,可将上述的转化体与形成微细胞能力较高的细胞,如小鼠A9细胞(Oshimura,M.,环境和健康展望,93:57,1991)进行整个细胞融合,由此可提高其形成微细胞的能力。已知在小鼠-人的杂交细胞中,人染色体的选择性消失,用上述的标记物进行选择,所得到的融合细胞能稳定保持标记了的人染色体。
为了满足2)的条件,希望从这种融合细胞得到微细胞,然后再度与小鼠A9细胞融合。这种情况下用人染色体上的标记物进行选择,所得到的微细胞株大多能满足1)、2)、3)的条件。可在最后得到的小鼠-人单染色体杂交细胞中,通过PCR(聚合酶链式反应,Saiki等,科学,239:487,1988)、Southern印迹分析(Ausukel等,现代分子生物学方法,John Wiley和Sons,Inc.1994)、FISH(荧光原位杂交,Lawrence等,细胞,52:51,1988)分析等来鉴定标记的人染色体。如果想转移特定的染色体,则对多个人细胞转化体克隆的每个克隆重复上述过程,检测目的染色体被标记的克隆。或者对人细胞转化体克隆的混合物实施上述过程,对所得的大量小鼠-人单染色体杂交细胞进行人染色体的鉴定。
此外,可通过所期望染色体上DNA序列的同源重组(Thomas等,细胞,51:503,1987),将标记基因插入到特定的部位。
用γ射线照射小鼠-人杂交细胞制备的微细胞,标记的人染色体成为片段,可移到小鼠A9细胞中。即使微细胞不经γ射线照射,部分片段化的人染色体也可以一定的比例转移。这时所得到的微细胞融合株保持了标记的人染色体的部分片段。当想把染色体部分片段转移到受体细胞的时候可应用这些克隆。
(2)向保持有分化多能性的小鼠细胞移入人染色体或其片段。
到目前为止已有这样的报道,将来源于各种品系小鼠的胚胎肿瘤细胞(EC细胞,Hanaoka等,分化,48:83,1991)、胚胎干细胞(ES细胞,Evan等,自然,292:154,1981)、胚胎生殖细胞(EG细胞,Matsui等,细胞,70:841,1992)注入小鼠的初期胚胎或与之共同培养,可形成正常体细胞,即可制成嵌合体小鼠。ES、EG细胞的这种能力尤其高,许多情况下它们可对生殖细胞起作用,由这些细胞可得到其后代。EC细胞主要从生殖细胞癌获得,ES细胞来源于胚泡的内部细胞块,EG细胞来源于胚胎发生初期出现的原始生殖细胞。作为本发明中移入人染色体的可用这些细胞株及受体细胞其突变株,以及在小鼠中能分化成其全部或部分正常体细胞的任何未分化细胞。
可用由(1)中得到的人染色体供体细胞制备的微细胞或经γ射线照射的微细胞作为移到受体细胞中的人染色体的物质。可用清水素行,细胞工程学手册,羊土社,1992中所记载的方法,将受体细胞与微细胞融合,从而把人的染色体转移到受体细胞。微细胞供体细胞中保持有在受体细胞中可以筛选某些人染色体或其片段的标记。应用同(1)一样的PCR、Sonthem印迹分析、FISH分析等方法,可以从中筛选保持有导入的目的基因或染色体或其片段的克隆,这样可转移任何人染色体或其片段。另外,依次导入含有不同选择标记的几个染色体或其片段,可得到同时保持这些标记的受体细胞。此外,可从已转移了人染色体的细胞克隆中选择移入的染色体数目增加了的克隆。这通常是通过提高加到培养液中的筛选药物的浓度而完成的。
通过下面的方法来证实经人染色体上的标记(G418耐性,等)所筛选的受体细胞是否保持供体细胞所保持的全部或部分人染色体。用从筛选的受体细胞中提取的基因组DNA、以人特异的重复序列(L1,Alu等,Korenberg等,细胞,53:391,1988)或人基因作探针进行Sonthern印迹分析。或者,用人基因特异的引物进行PCR,以及用人染色体特异的探针进行荧光原位杂交(FISH)等染色体分析方法来进行验证。
(3)从导入人染色体的ES细胞制备嵌合体小鼠
参照相沢慎一,基因工程手册,8,基因打靶,羊土社,1995等所描述的方法,从(2)中得到的ES细胞株制备嵌合体小鼠。为了有效地制备嵌合体小鼠,在选择宿主胚胎的发生期、品系时,希望用对各个ES细胞株已回顾过的条件。例如,对于CBA×C57BL/6F1来源的TT2细胞(野鼠色,Yagi等,分析生物化学,214:70,1993),希望选用Balb/c(白化体,日本CREA社)或ICR(白化体,日本CREA社)来源的8细胞期胚胎作宿主胚。
(4)嵌合体小鼠中人染色体的保持及人基因的表达。
通过毛色来粗略判断在由注入了ES细胞的胚胎而产生的小鼠中,ES细胞的贡献率。但是,即使对毛色一点贡献也没有也不能判断没有贡献给其它组织。更为详细的,可以嵌合体小鼠的各组织中提取基因组DNA,用Sonthern印迹分析、PCR等方法来证实各组织中是否保持人染色体。
通过以下的方法来证实导入的人染色体上基因的表达。用各组织的RNA、通过RT-PCR法(Kawasaki等,美国国家科学院院刊,85:5698,1988)、Northern印迹法(Ausubel等,同前)来检测人染色体mRNA的表达。可用与小鼠同种蛋白质的交差反应性最小的抗人蛋白质抗体,通过酶免疫测定法(ELISA,富山·安东,单克隆抗体实验手册,讲谈社Sentific,1987;石川,超高敏感度酶免疫测定法,学会出版中心,1993)、Western印迹法(Ausubel,同前)或者利用电泳泳动度的差别进行同功酶分析(Koi等,日本癌症研究,80:413,1989)等方法来检测蛋白质水平的表达。另外,嵌合体小鼠细胞中人染色体的保持及该染色体上基因的表达,可通过在嵌合体小鼠原代培养细胞中出现表达耐药性标记基因的耐药性细胞而证实的。
例如,用与小鼠抗体产生交差反应性最小的抗人Ig抗体进行酶免疫测定法,可检测由ES细胞所制备的嵌合体小鼠的血清中人IgM、IgG、IgA等,而该ES细胞保持存在人免疫球蛋白重链的人14号染色体中。或者,用来源于人的抗原(如血清白蛋白)免疫嵌合体小鼠,将其脾细胞与小鼠的骨髓瘤融合,通过ELISA对此杂交瘤(安东·千叶,单克隆抗体实验操作入门,讲谈社Scientific,1991)进行筛选,可得到能产生人免疫球蛋白重链的杂交瘤。
以上以小鼠为例说明了保持人染色体或其片段、表达该染色体或其片段上基因的嵌合体非人动物的制作方法,但本发明中,移入到嵌合体非人动物的染色体或其片段并不限于人,而包括广泛的外源染色体或其片段,并表达该染色体或其片段上的基因。这里“外源染色体”是以转移到保持有分化多能性的细胞中,在嵌合体非人动物中能表达该染色体或其片段上的基因为特征,其来源的生物种类没有特别限制。应用本发明的方法,不仅可制作嵌合体小鼠,也可制作其它的嵌合体动物,如大鼠、猪等哺乳类及其它种类的嵌合体动物。有这样的报道,ES细胞或ES样细胞在小鼠以外的动物种类中建立,如在大鼠(Iannaccone等,发育生物学,163,288-,1994)、猪(Wheeler等,繁殖、生殖和发育,6,563-,1994)、牛(Sims等,美国国家科学院院刊,91,6143-6147,1994)等,也有用鳉、鸡等做过偿试(转基因动物,蛋白质核酸酶,1995年10月增刊,共立出版)。已知,将ES样细胞(ED细胞)至少经过10代传代以上而获得的上皮样细胞的核移植到未受精卵中,这样在绵羊中可正常发育(Campben等,自然,380,64-,1996)。这样通过以ES或ES样细胞作受体细胞,移入外源染色体,可与用小鼠一样,制作出保持外源染色体或其片段、表达该染色体或其片段上基因的嵌合体非人动物。
本发明中,保持被转移的外源染色体或其片段,具有分化多能性的细胞并不局限于上述的ES细胞、EC细胞和EG细胞。例如可以将外源染色体或其片段转移到骨髓干细胞,而通过这种向骨髓干细胞活体的移植,可进行遗传病的治疗。
保持外源染色体的ES细胞在嵌合体非人动物中分化成生殖细胞时,其所繁殖的后代也能保持转移的染色体或其片段,并能表达导入染色体或其片段上的基因。
利用上面所得到的嵌合体非人动物或其后代来表达外源染色体或其片段上的基因,回收表达产物,可制造生物活性物质。具体而言,在能表达外源染色体或其片段上基因的条件下饲养嵌合体非人动物或其后代,然后从动物的腹水和血液中回收表达产物。或者在能够表达外源染色体或其片段上基因的条件下培养嵌合体非人动物或其后代的组织、细胞或获得不死性的细胞(例如,与骨髓瘤细胞融合而形成的不死性杂交瘤),然后从培养物中回收表达产物。此外可将从这些嵌合体非人动物、或其后代的组织、细胞或它们的不死性细胞中提取的外源染色体或其片段、或将构成外源染色体或其片段的DNA、或将嵌合体非人动物或其后代的组织、细胞或它们的不死性细胞中保持的外源染色体或其片段来源的cDNA转化到动物细胞或昆虫细胞如CHO细胞、BHK细胞、肝癌细胞、骨髓瘤细胞、SF9细胞),在表达外源染色体或其片段上基因的条件下培养该细胞,从培养物中回收表达产物(如特定抗原特异的抗体蛋白质等)。用离心等众所周知的方法进行回收,并用硫酸铵分级分离、分配色谱、凝胶过滤色谱、吸附色谱、制备薄层色谱等已知的方法进行纯化。生物学活性物质包括外源染色体上所编码的任何物质,如抗体、尤其是抗人抗体。例如,可克隆从嵌合体动物的脾细胞或其杂交瘤等不死性细胞而来的染色体上的人抗体基因,转移到中国仓鼠卵巢细胞(CHO)和骨髓瘤细胞,就能生产人抗体(Lynette等,生物工程学,10,1121-,1992;Bebbington等,生物工程学,10,169-,1992)。
本发明中所得到的保持人2号、14号、22号染色体(片段)的嵌合体小鼠及其后代保持有人抗体重链(14号染色体上)、轻链κ(2号染色体上)、轻链λ(22号染色体上)各种基因功能序列的大部分。也就是说,与利用酵母人工染色体等转移人抗体基因的一部分这一已知的转基因小鼠相比(Green等,Nature Genetics,7,13-,1994,Lonberg等,自然,368,856-,1994等),能够表达与人非常相似的、非常多样的人抗体库(repertory)。另外,依照本发明所得到的同时保持2号+14号、22号+14号等染色体(片段)的嵌合体小鼠及其后代、以及它们交配所得到的同时保持2号+14号+22号等染色体(片段)的小鼠及其后代,可产生重链、轻链二者均来源于人的完全人抗体。这些小鼠可识别人抗原,把它当作异物与之进行免疫反应,这样可产生抗原特异的人抗体。这些特性对于得到治疗用的人单克隆抗体、多克隆抗体非常有益(Green等,同前;Lonberg等,同前)。另一方面,为了更有效地获得对特定抗原亲和性高的人抗体,希望能制成不产生小鼠抗体、只产生人抗体的小鼠(Green等,同前;Lonberg等,同前)。本发明中可根据下面已知的方法A或方法B来完成。
方法A:利用小鼠抗体缺陷的ES细胞及小鼠抗体缺陷的嵌合体宿主胚的方法
方法B:从导入了人染色体的嵌合体小鼠得到保持人染色体的后代,与小鼠抗体基因缺陷的小鼠品系进行交配的方法
以下对A、B每种方法的典型实施例进行具体描述。
方法A的具体过程
1.用目的基因同源重组(Joyner等,基因打靶,1993,IRL出版)的方法破坏小鼠ES细胞中以2拷贝存在的小鼠抗体重链基因中的一个。随后通过定点特异重组除去基因破坏部分两侧可能的序列,如loxP序列(用Cre重组酶进行重组,Sauer等,同前,用FLP重组酶-FRT序列O′Gorman等,科学,251,1351-,1991等例),在基因破坏的部位插入G418耐药基因等标记基因。
2.在高浓度药物存在的条件下培养上述一个抗体重链基因被破坏了的耐药性小鼠ES细胞,选择耐高浓度药物的克隆。通过筛选这些克隆可获得两个抗体重链基因均被破坏了的克隆(相沢慎一,同前)。
3.将在1中耐药基因的两端插入的重组序列间引起定点特异性重组的酶基因,如Cre重组酶基因(Sauer等,同前)瞬时转移到2中所得到的2个抗体重链基因场被破坏了的小鼠ES细胞中,去除插入到能在loxP序列间引起重组反应的2个抗体重链基因中的耐药性基因,选择这样的药物敏感性克隆(高津圣志等,实验医学分册,免疫研究的基本技术,p255-,1995,羊土社)。
4.对小鼠抗体轻链κ基因重复上述1-3的操作过程,最终可获得抗体重链和κ链完全缺失的药物敏感性克隆。
5.以4的克隆(抗体重链、κ链缺失的小鼠ES细胞)作受体细胞,通过微细胞融合导入用耐药标记基因(如G418耐药基因)标记了的、含有人抗体重链基因的人14号染色体(片段)。
6.以5中所得到的克隆为受体细胞,通过微细胞融合,导入用与5中不同的耐药标记基因(如嘌呤霉素耐药基因)标记了的、含有人抗体轻链基因的人2号染色体(片段)或22号染色体或二者均导入。
7.以从不能产生自身抗体的小鼠品系(如RAG-2基因敲除小鼠,Shinkai等,细胞,68,855-,1992;膜型μ链敲除小鼠,Kitamura等,自然,350,423-,1991)得到的胚胎为宿主胚,用6中得到的ES细胞制作嵌合体小鼠。
8.所得嵌合体小鼠中的大多数功能性B淋巴细胞来源于ES细胞(高津圣志等,实验医学分册,免疫研究的基础技术,P234,羊土社,1995)。由于这种B淋巴细胞缺乏小鼠重链、κ链,所以主要由导入的染色体上功能性人抗体基因的表达而产生人的抗体。
方法B的具体过程
1.从保持含有人抗体重链、轻链κ或轻链λ的人染色体或其片段的嵌合体小鼠得到能稳定保持这些人染色体或其片段并可连续传给下一代的后代。
2.从1中所得能表达抗体重链或轻链的小鼠品系或由其交配而得到表达人抗体重链和轻链的小鼠品系,将该品系与缺乏自身抗体基因的小鼠品系(如膜型μ链基因敲除小鼠,同前;轻链κ敲除小鼠,Chen等,EMBO J.,3,821-,1993)进行交配,可得到缺乏小鼠抗体重链及轻链的纯合子及保持含人抗体重链(14号)+轻链κ(2号)、抗体重链(14号)+轻链λ(22号)或人抗体重链(14号)+轻链κ(2号)+轻链λ(22号)的人染色体的小鼠品系。在这一小鼠品系中由于缺乏小鼠抗体重链、轻链κ基因,所以只要由于转移的染色体上功能性人抗体基因的表达而只产生人的抗体。
上述的方法A和方法B不只用于生产人抗体,而且可用于生产外源染色体上存在的任何基因的表达产物。
以下用实施例进行具体说明,但本发明并不限于这些实施例。(实施例1)保持G418耐性标记的人染色体(片段)的染色体供体细胞的制备
用限制性酶SalI(宝酒造)将含有G418耐性基因的质粒pSTneoB(Katoh等,细胞的结构、功能,12:575,1987;日本研究生物制品保藏中心(JCRB),保藏号:VE 039)线性化,导入人的正常纤维母细胞HFL-1(RIKEN细胞库,RCB0251)。用胰酶处理HFL-1细胞,以5×106细胞/ml的浓度悬浮在Dulbecco′s磷酸缓冲盐水(PBS)中,在10μg DNA存在的情况下,用基因脉冲仪(Bio-Rad)进行电穿孔(石田等,细胞工程学实验操作入门,讲谈社,1992)。以25μF的电容,用4mm距离的电穿孔细胞(165-2088,Bio-Rad)在室温下加1000V的电压。将电穿孔的细胞接种到3-6个100mm的组织培养用的塑料平皿中(Corning),其中含有增加了15%胎牛血清(PBS)的Eagle′s F12培养基(以下称作F12)。1天后,换用含有200μg/ml G418(GENETICIN,Sigma),添加了15%FBS的F12培养基。2-3周后,收集形成的克隆,100个为一组,共52组,然后将每一组的克隆重新接种到100mm的平皿中,进行培养。
用添加了10%胎牛血清(PBS)的Dulbecco′s改良培养液(以下为DMEM),在100mm的平皿中培养小鼠A9细胞(Oshimura,环境和健康展望.,93:57,1991,JCRB0211)。分别在100mm平皿中用补充了15%胎牛血清(FBS)和200μg/ml的G418的F12培养52组G418抗性HFL-1细胞。用胰酶处理小鼠A9细胞和HFL-1细胞,每种细胞取1/4-1/2进行混合,在100mm平皿中将添加了10%胎牛血清(FBS)的DMEM与添加了15%胎牛血清(FBS)的F12进行等量混合,将细胞接种到混合物中培养半天到一天。参照(清水等,细胞工程学手册,羊土社,p.127-,1992)所描述的方法进行细胞融合。用DMEM洗涤细胞表示2次,之后在2ml PEG(1∶1.4)溶液中处理1分钟,再换用2ml PEG(1∶3)溶液处理1分钟。吸去PEG溶液后用无血清培养基(DMEM)洗涤3次,之后用常规的培养基(10%FBS、DMEM)培养1天。用胰酶处理细胞使之分散,悬浮剂含有乌木箭毒苷(1×10-5M,Sigma)和G418(800μg/ml)的二重选择培养基中,接种到3枚100mm的平皿中。大约培养3周后,用胰酶处理生成的克隆使之分散,在含有G418(800μg/ml)的选择培养基(10%FBS、DMEM)上培养。
用胰酶处理细胞使之分散后,将2组收集为1组,在6个25cm2离心用的培养瓶中(Costar,3025)培养到细胞密度达到70-80%的汇合度。换成含Colcemid(0.05μg/ml,demecolcine,和光纯药)的培养基(20%FBS、DMEM),培养2天以形成微细胞。去除培养液,用预温(37℃)的细胞松弛素B溶液(10μg/ml,Sigma)充满离心用培养瓶,将瓶插入到丙烯酰基离心容器内,在34℃,以800rpm离心1小时。当小鼠A9细胞达到80%汇合状态时,向25cm2瓶中加入纯化的微小核,用PEG溶液进行融合。在含G418的选择培养基中分离形成的克隆。用以下的方法来鉴定各克隆中所保持的人染色体(2号、4号、14号、22号)。除上面以外的所有实验操作及试剂均参照(清水等,细胞工程学手册,羊土社,p.127-)。
(1)PCR分析
培养分离的细胞,用Puregene DNA分离试剂盒(Gentra System社)提取基因组DNA,以该基因组DNA为模板、用人染色体特异的引物、通过PCR来选择保持有2、4、14、22号人染色体的克隆。PCR的扩增大约使用0.1μg的基因组DNA(Innis等,PCR实验手册,HBJ出版局,1991,热循环仪,GeneAmp 9600,Perkin-Elmer社)。Taq聚合酶购自Rerkin-Elmer公司,反应条件是94℃、5分钟1循环后、94℃变性,15秒、54℃~57℃、退火15秒钟(根据引物而适当改变)、72℃延伸20秒,进行35个循环。可用染色体上存在的基因(O′Brien,基因图谱,第6版,Book 5,Cold Spring HarborLaboratony Press,1993)和多形性标记(多形性STS引物对,BIOS社;Weissenbach等,自然359:794,1992;Walter等,Nature Genetics,7:22,1994)等作引物。可以GenBank、EMBL等数据库获得的碱基序列为基础来制作基因引物。下面将以实施例来说明每个染色体的多形性引物的名称和基因引物的序列(2号:实施例1,4号实施例6,14号实施例9,22号实施例2)。用以下所示的基因标记和多形性标记(多形性STS引物对,BIOS社:D2S207,D2S177,D2S156,D2S 159。BIOS社)来鉴定2号染色体。
Cκ(免疫球蛋白κ恒定区):5′-TGGAAGGTGGATAACGCCCT(序列号1),5’-TCATTCTCCTCCAACATTAGCA(序列号2)
FABP1(肝脂肪酸结合蛋白-1):5’-GCAATCGGTCTGCCGGAAGA(序列号3),5’-TTGGATCACTTTGGACCCAG(序列号4)
Vk3-2(免疫球蛋白κ可变区):5′-CTCTCCTGCAGGGCCAGTCA(序列号5),5′-TGCTGATGGTGAGAGTGAACTC(序列号6)
Vk1-2(免疫球蛋白κ可变区):5’-AGTCAGGGCATTAGCAGTGC(序列号7),5’-GCTGCTGATGGTGAGAGTGA(序列号8)
(2)荧光原位杂交(FISH)
根据(松原等,FISH实验方案,秀润社,1994)所记载的方法,用人2号、4号、14号、22号染色体特异的探针(CHROMOSOMEPAINTING SYSTEM,Cambio社)进行FISH分析。
例如在26组(745克隆)中有10组可以得到保持2号染色体的克隆。其中有5个克隆对2号染色体特异的引物全部呈阳性,将这些克隆进行FISH分析。根据(松原等,FISH实验方案,秀润社,1994)所记载的方法,用人2号染色体特异的探针(CHROMOSOMEPAINTING SYSTEM,CANBIO社)进行FISH分析。在对所有引物呈现阳性的细胞中可观察到人2号染色体的完整形态,在只对一部分引物阳性的一些克隆中可观察到比人2号染色体较小的独立染色体,或观察到含有与人2号染色体以外的染色体发生融合的染色体的细胞(图1)。图1中横轴为克隆名称,纵轴为PCR中应用的引物。●表示阳性,×表示阴性。底行表示通过FISH所观察到的人2号染色体存在的形态。未做描述的表明没有实施。
同样可得到保持人2号、14号、22号染色体的A9细胞。(实施例2)用微细胞法将人22号染色体导入小鼠ES细胞。
用(实施例1)中得到的保持人22号染色体的A9细胞株(以下称作A9/#22)作染色体供体细胞。用小鼠ES细胞株E14(Martin L.Hopper,Hooper等,自然,326:292,1987)作染色体受体细胞。参照(相沢慎一,生物手册,8,基因打靶,羊土社,1995)所描述的方法培养E14细胞,并用丝裂霉素c(Sigama)处理的G418耐性STO细胞株(大阪大学,近藤寿人教授)作饲养细胞。首先,参照清水等(细胞工程学手册,羊土社,1992)所报道的方法,从大约108A9/#22制备微细胞。将所得微细胞悬浮到5ml的DMEM中。用胰酶分散大约107的E14,之后用DMEM洗涤3次,悬浮到5ml的DMEM中,然后混合微细胞,1250rpm、离心10分钟,弃上清。轻扣沉淀使之分散,加入0.5ml的1∶1.4PEG溶液(5g PEG 1000(和光纯药)、1mlDMSO(Sigma)溶解到6ml DMEM中),室温静置1分30秒,然后缓慢加入10ml的DMEM。立即离心1250rpm,10分钟,弃上清,将沉淀悬浮到30ml培养ES细胞用的培养基中,接种到3个直径100mm、接种了饲养细胞的组织培养用塑料瓶(Corning)中。24小时后,换成添加了300μg/ml G418(GENETICIN,Sigma)的培养基,其后,每天换培养基。1周-10天后出现耐药性克隆,其出现频率为每107个E14细胞出现0-5个。挑选克隆进行增殖,每5×106个悬浮到1ml的保存培养基<ES细胞培养液+10%DMSO(Sigma)>中,-80℃冻存。同时用Puregene DNA分离试剂盒(Gentra System社)从每种耐药克隆的106×107个细胞中制备基因组DNA。
用γ射线照射微细胞使人22号染色体片段化(koi等,科学,260:361,1993)。将从108个A9/#22所得的微细胞悬浮到5ml DMEM中,用Gammacell 40(加拿大原子能公共公司)在冰上进行60Gyγ射线的照射(1.2Gy/分×50分)。经γ射线照射的微细胞的融合与耐药株的选择与未经照射的微细胞一样,其选择结果是,耐药株的出现频率是每107个E14细胞出现1-7个。与未照射的情况一样对耐药株进行冻存、提取DNA。
根据下面(1)~(3)的方法来证实未经照射的微细胞耐药株E14/#22-9、E14/#22-10、γ射线照射的微细胞耐药株E14/#22-14、E14/#22-25中所导入染色体的保持。
(1)PCR分析(图2)
以耐药株基因组DNA为模板,通过PCR来检测人22号染色体上存在的基因((Genetic Maps)基因图谱,同前)和多形性标记(多形性STS引物对,BIOS社:D22S315,D22S275,D22S278,D22S272,D22S274;自然,359:794,1992)的存在。下面是以GenBank、EMBL等的数据库获得的碱基序列为基础制备的基因引物-寡核苷酸序列。
PVALB(小白蛋白):5’-TGGTGGCTGAAAGCTAAGAA (序列号9),5’-CCAGAAGAATGGTGTCATTA(序列号10)
MB(肌红蛋白):5′-TCCAGGTTCTGCAGAGCAAG(序列号11),5’-TGTAGTTGGAGGCCATGTCC(序列号12)
DIA1(细胞色素b-5还原酶):5′-CCCCACCCATGATCCAGTAC(序列号13),5’-GCCCTCAGAAGACGAAG CAG(序列号14)
Igλ(免疫球蛋白λ):5’-GAGAGTTGCAGAAGGGGTGACT(序列号15),5’-GGAGACCACCAAAC CCTCCAAA(序列号16)
ARSA(芳基硫酸酯酶):5’-GGCTATGGGGACCTGGGCTG(序列号17),5’-CAGAGACACAGGCACGTAGAAG(序列号18)
以大约10μg的基因组DNA作模板,用上述10种引物进行PCR扩增(Innis等,同前)。其结果是未经照射的2个克隆对所有的引物、经γ射线照射的2个克隆对部分引物产生所期待长度的扩增产物。图2示上述结果。图2中左边是以人22号染色体的G带为基础制作的染色体模式图和一些位置明确的标记在G带的位置(O′Brien,基因图谱,第6版,第5册等)。根据目前获得的信息(科学,人类基因图,1994;Nature Genetics,7:22,1994;自然,359:794,1992等),基因和多形性标记的排列表明其大致的位置关系,其顺序未必准确。用PCR对4种G418耐性E14细胞克隆进行检测,能得到所期待扩增产物的标记用■表示,不能检测出的标记用□表示。下面是通过FISH分析观察到的结果。A9/#22是染色体供体细胞。
(2)Southern印迹分析
参照(Ausubel等,现代分子生物学方案,John Wiley Sons公司,1994)所描述的方法,以人特异的重复序列L1序列(每单倍体基因组存在104-105拷贝,来自RIKEN DNA Bank;核酸研究,13:7813,1995;来自pUK的1.4kb的EcoRI-BamHI片段)作探针,对经限制酶(BglII,宝酒制造)处理的约2μg的基因组DNA进行Southern印迹分析。其结果是,在各耐药克隆DNA中,大多能检测出与人L1序列进行杂交的杂交带。有2个未照射的克隆,其带型和根据各带密度而推测出的人染色体DNA对小鼠基因组DNA的定量比与A9/#22相同。γ射线照射克隆的全部信号强度与A9/#22相比,与PCR分析所显示的缺损程度相关。
(3)荧光原位杂交(FISH)
参照(松原等,FISH实验方案,秀润社,1994)所记载的方法,应用人22号染色体特异的探针(CHROMOSOME PAINTINGSYSTEM,Cambio社)进行FISH分析。其结果是,在几乎所有观察到的分裂像中,对于E14/#22-9能检测到异位到小鼠染色体上的形态,而对于其它3株则能检测到独立的人22号染色体。
通过以上实验可知,所得到的G418耐药株E14/#22-9、E14/#22-10保持全部或大部分的人22号染色体,E14/#22-14、E14/#22-15则保持有其部分片段。
(实施例3)从保持人22号染色体的ES细胞制备嵌合体小鼠。
(实施例3)由保持人22号染色体的ES细胞制备嵌合体小鼠
参照(相沢慎-,生物手册,8,基因打靶,羊土社,1995)所记载的方法,用常规技术从小鼠获取胚胎、培养,将ES细胞注入胚胎,移植到代孕母亲的子宫等。将(实施例2)中得到的、经证实保持人22号染色体的、冻存的G418耐性ES细胞株E14/#22-9融化,培养,注入到C57B2/6×C3H F1雌性小鼠(日本CREA社)与C3H雄性小鼠(日本CREA社)交配而得到的胚泡期胚中,每一胚胎注入约10-15个细胞。在代孕母亲ICR小鼠(日本CREA社)伪妊娠2.5天后,将注入大约10个细胞的胚胎移植到子宫的两侧。(第1表)表示了其结果。
表1.从保持人22号染色体(片段)的E14细胞制备嵌合体小鼠
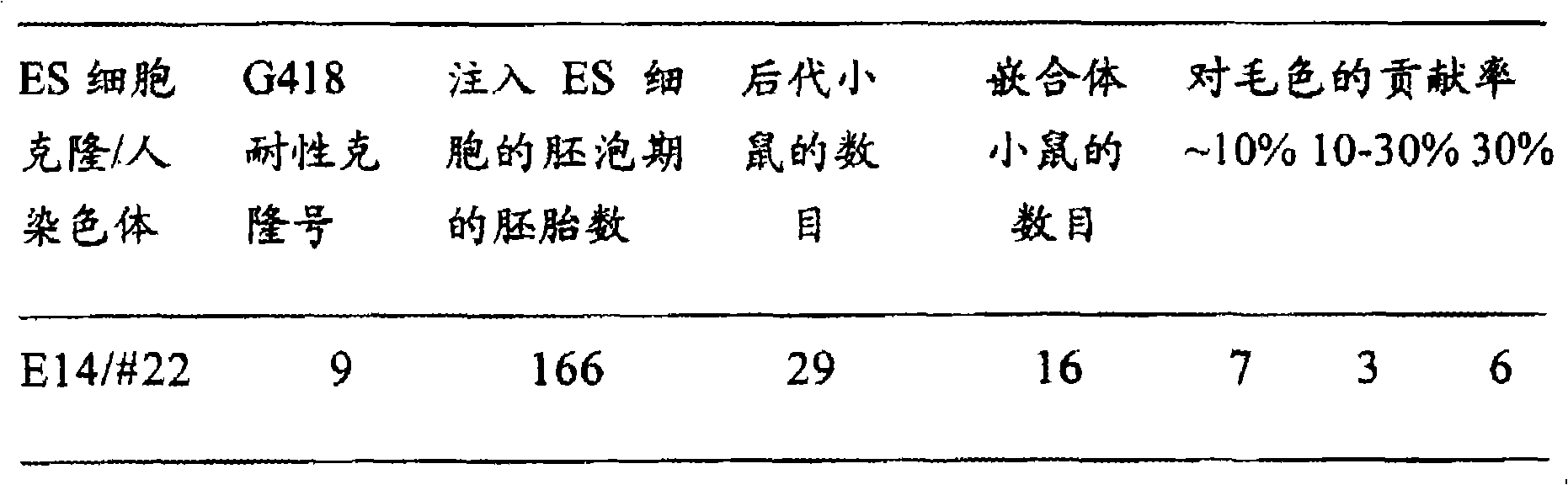
其移植了166个注入胚,诞生了29只子鼠。由宿主胚而来的毛色为野鼠色(深棕色),其中若能见到由E14细胞而来的浅灰色,则可判定为嵌合体。所生29只中具有明显浅灰色的,也就是说能识别出由E14细胞贡献的个体有16只。在K22-22中最大的贡献率为40%。
根据该结果可表明保持人22号染色体的小鼠ES细胞株/#22-19能形成嵌合体,即使有分化或小鼠正常组织的能力。
(实施例4)在保持人22号染色体的ES细胞而来的嵌合体小鼠的各组织中证实人染色体DNA的保持。
在(实施例3)中用毛色判断的基础上,以从尾部制备的基因组DNA为模板,通过PCR来证实导入染色体的保持。参照(腾木元也,发育工程学实验手册,讲谈社Scientific,1987)的方法,从出生后3周的嵌合体小鼠获取尾部,用Puregene DNA分离试剂盒提取DNA。以该基因组DNA为模板,应用(实施例2)中使用的多形性引物中的PVALB、D22S278,来证实扩增产物。对能观察到毛色贡献的10只小鼠进行分析,结果在所有的小鼠中可检测到至少由一引物而产生的扩增产物。
Southern印迹分析与(实施例2)相同,用人L1序列作引物,对从6个嵌合体小鼠、1个非嵌合体小鼠尾部得到的2μg基因组DNA进行分析。结果可证实在所有的嵌合体小鼠中存在多数的人L1序列,其带型与E14/#22-9相类似。与小鼠基因组的定量比率最多可达到10%(图3)。图3中每道中均使用BglII消化的2μg基因组DNA。用32P标记的人L1序列作探针,通过图像分析仪BAS2000(富士照相胶片公司)来检测信号。从右边数是由嵌合体小鼠(K22-6,7,8,9,10,11,12:9为非嵌合体)尾部而来的基因组DNA和对照DNA(C:C是E14/#22-9与E14基因组DNA以1∶9的重量比混合)。左边表示DNA分子量,右边表示各嵌合体的嵌合率。(-:0%;+:~10%;++:10~30%)。
用ISOGEN(日本Gene公司)从毛色贡献达到5%的嵌合体小鼠(K22-7)的脑、肝脏、肌肉、心脏、脾、胸腺、卵巢、肾获取基因组DNA,用(实施例2)中所用基因引物中的MB、DIA1,对各种组织进行PCR分析。结果是在所有的组织中,2个引物均产生所期待的扩增产物。(图4)是用DIA1引物所产生的结果。PCR产物经20%琼脂糖凝胶电泳后,用溴化乙锭染色进行检测。图4中每一泳道从左开始为B:脑,L:肝,SM:骨骼肌,H:心脏,Sp:脾,Th:胸腺,Ov:卵巢,K:肾,nc:非嵌合体小鼠尾部的NDA(阴性对照),pc:人纤维母细胞(HFL-1)DNA(阳性对照)。
该结果表明,E 14/#22-9在小鼠各种正常组织中起作用,并确证保持有人22号染色体。
(实施例5)由保持人22号染色体的ES细胞而来的嵌合体小鼠中人基因的表达
将毛色贡献达5%个体(K22-7)的尾部在液氮中冻结粉碎后,可作为证实人基因表达的材料。这是皮肤、骨、骨肉、血液等组织的混合物。用ISOGEN(日本Gene)从中提取总RNA,通过RT-PCR检测人肌红蛋白(MB)、人细胞色素b5还原酶(DIA1)的mRNA。参照(Innts等,PCR实验手册,HBJ出版局,1991)的方法进行RT-PCR。用随机六聚物寡核苷酸(终浓度1000pmol,宝酒制造)作逆转录反应的引物,逆转录酶使用BRL公司的Super Script。下面是用cDNA作模板进行扩增时用的引物。
MB:5′-TTAAGGGTCACCCAGAGACT (序列号19),5′-TGTAGTTGGAGGCCATGTCC(序列号20)
DIA1:5′-CAAAAAGTCCAACCCTATCA(序列号21),5′-GCCCTCAGAAGACGAAGCAG(序列号22)
该结果可检测出两基因mRNA特异的扩增产物(图5)。RT-PCR产物经20%琼脂糖凝胶电泳后,用溴化乙锭染色进行检测。图5中M表示标准(Hind III消化λDNA+Hae III消化X174DNA,宝酒造),MB表示人肌红蛋白,DIA1为细胞色素-b5-还原酶,WT为野生型C3H小鼠。
由对同一个体(K22-7),用ISOGEN从脑、心脏、胸腺、肝、脾、肾、卵巢、骨骼肌提取总RNA,用上述2种引物对各脏器进行RT-PCR。结果在所有的脏器中发现DIA1所期待的扩增产物,仅在心脏和骨骼肌中发现MB所期待的扩增产物(图6)。肌红蛋白对肌细胞特异性表达(Bassel-Duby等,MCB,12:5024,1992),由此可知导入的人染色体上的基因在小鼠中能接受正常组织特异性表达的调控。PCR产物经20%琼脂糖凝胶电泳后,用溴化乙锭染色进行检测。图6中,各泳道从左开始分别为B:脑,H:心脏,Th:胸腺,L:肝脏,Sp:脾脏,K:肾,Ov:卵巢,SM:骨骼肌,M:标准(同前)。据认为MB的结果中所观察到的低分子量带是非特异性产物。
总之,导入的人22号染色体在嵌合体小鼠的正常组织中能发挥功能。
(实施例6)将人4号染色体或其部分片段导入ES细胞
用(实施例1)中得到的保持人4号染色体的小鼠A9细胞(以下称作A9/#4)作为染色体供体细胞。用小鼠ES细胞的E14(与实施例2相同)作染色体受体细胞。微细胞融合试验及G418耐性克隆的选择与(实施例2)相同。耐药性克隆出现的频率为每107个E14细胞出现1-2个。耐药性克隆的冻存、基因组DNA的提取与(实施例2相同)。用以下(1)-(3)的方法来证实耐药性克隆E14/#4-4、E14/#4-7、E14/#4-11中人染色体或其片段的保持情况。
(1)PCR分析(图7)
以耐药株基因组DNA为模板,通过PCR来检测人4号染色体上存在的基因((Genetic Maps)基因图谱,同前)和多形性标记(多形性STS引物对,BIOS社:D4S395,D4S412,D4S422,D4S413,D4S418,D4S426,F11;自然,359:794,1992)的存在。下面是以Gen Bank、ENBL等数据库获得的碱基序列为基础制备的基因引物-寡核苷酸序列。
HD(享廷顿疾病):5′-TCGTTCCTGTCGAGGATGAA(序列号23),5′-TCACTCCGAAGCTGCCTTTC(序列号24)
IL-2(白介素-2):5′-ATGTACAGGATGCAACTCCTG(序列号25),5′-TCATCTGTAAATCCAGCAGT(序列号26)
KIT(c-kit):5′-GATCCCATCGCAGCTACCGC(序列号27),5′-TTCGCCGAGTAGTCGCACGG(序列号28)
FABP2(脂肪酸结合蛋白2,肠):5′-GATGAACTAGTCCAGGTGAGTT(序列号29),5′-CCTTTTGGCTTCTACTCCTTCA(序列号30)
对上述11种引物进行PCR扩增,结果有3株能得到全部或部分引物所期待的扩增产物。也可看到象E14/#4-4、E14/#4-7一样缺乏一部分区域。以上结果如图7所示。图7中,左边是以人4号染色体的G带为基础制作的染色体模式图和一些位置明确的标记在G带的位置(参照实施例2)。根据目前的资料(参照实施例2)基因和多形性标记的排列表明其大致的位置关系,其顺序末必准确。用PCR对3种G418耐性E14细胞克隆进行检测,能得到所期待扩增产物的标记用■表示,不能检测出的标记用□表示。下面是通过FISH分析观察到的结果。A9/#4是染色体供给细胞。
(2)Southern印迹分析(图8)
与(实施例2)的方法相同,用人L1序列作探针,对E14/#4-4、E14/#4-7的基因组DNA进行Southern印迹分析。其结果是,在两株DNA中,大多能检测出与人L1序列进行杂交的杂交带。与A9/#4相比,全部信号强度与PCR分析所显示的缺损程度相关。图8中,各泳道均使用经BglII消化了的2μg基因组DNA。以32P标记的人L1序列作探针,通过图像分析仪BAS2000(富士照相胶片公司)检测信号。图8中,各道从左依次为1:A9/#4(染色体供体细胞)、2:A9/#4+A9(1∶2)、3:A9/#4+A9(1∶9)、4:A9、5:E14/#4-7、6:E 14/#4-4。2、3中2种DNA按括号内的比例进行混合。左侧为分子量。
(3)荧光原位杂交(FISH)
用与(实施例2)相同的方法,用人4号染色体特异的探针(CHROMOSOME PAINTING SYSTEM,Cambio社)进行FISH分析。结果在3株几乎所有的分裂像中能检测出人4号染色体或其部分片段。E14/#4-4中能检测到易位到小鼠染色体上的形态,其它2株才能检测到独立的染色体。所观察到的人染色体的相对大小当用PCR分析的结果推测出的一致。
以上实验表明,所得G418耐药株保持人4号染色体的全部或部分片段。
(实施例7)由保持人4号染色体部分片段的ES细胞制备嵌合体小鼠
将证实保持人4号染色体部分片段的G418耐性ES细胞株E14/#4-4、E14/#4-7的冻存株融化,注入到按(实施例3)的方法所得胚泡期胚胎中,每个胚胎注入10~15个细胞。代孕母亲ICR小鼠(日本AREA)伪孕2.5天后,移植到其子宫中,每侧子宫移入大约10个ES细胞注入胚。结果如(表2)所示。
表2.由保持人4号染色体(片段)的ES细胞制备嵌合体小鼠
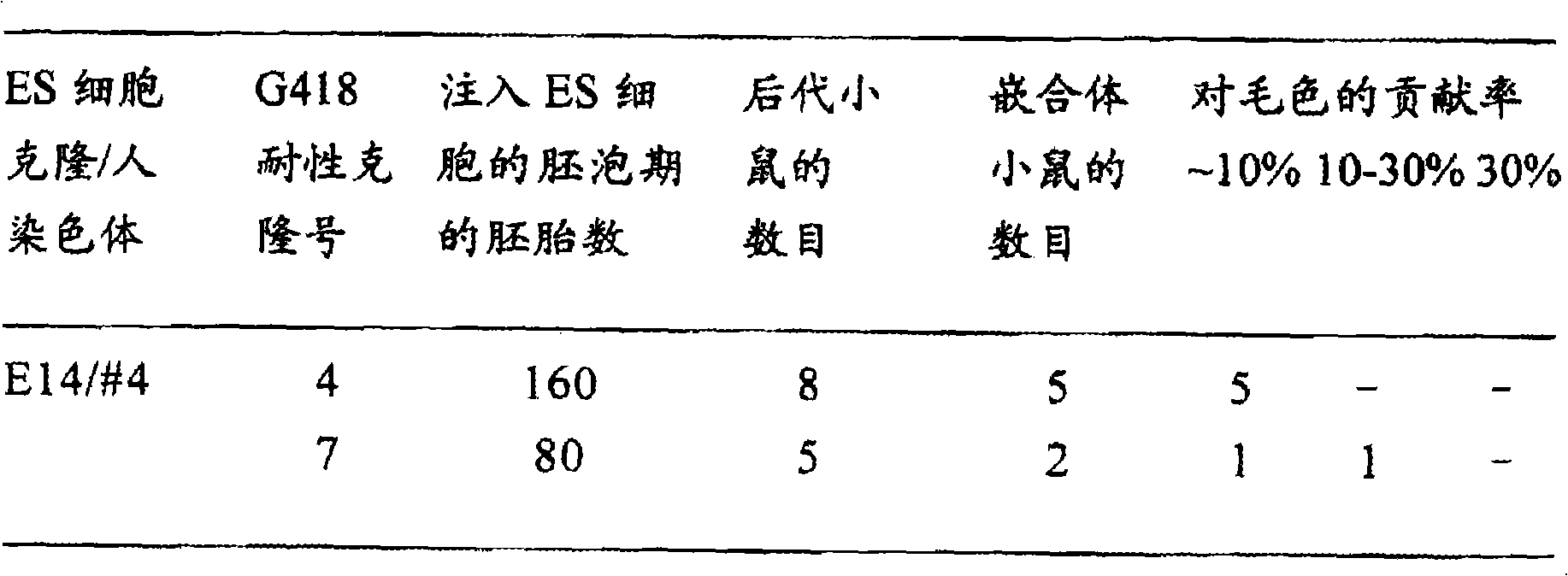
其移植了240个注入胚,诞生了13只子鼠。由宿主胚而来的毛色为野鼠色(深棕),其中若能见到由E14细胞而来的浅灰色,则可判定为嵌合体。所生13只中具有明显浅灰色的,也就是说能识别出由E14细胞贡献的个体有7只。在E14/#4-7的1小鼠中,最大的贡献率为15%。
根据该结果可表明保持人4号染色体的小鼠ES细胞株E14/#4-4、E 14/#4-7能形成嵌合体,即使有分化或小鼠正常组织的能力。(实施例8)在保持人4号染色体的ES细胞而来的嵌合体小鼠中证实人染色体DNA的保持和G418耐性基因的表达
(1)PCR分析
(实施例7)中所得嵌合体小鼠中,来源于E14/#4-7的1只(K#4-7-1:嵌合率约5%)、来源于E14/#4-4的一只(K#4-4-41:嵌合率约5%),用(实施例4)的方法,从它们的尾部制备基因组DNA。以该DNA为模板,用(实施例6)中分析4号染色体用的引物中检测出E14/#4-7、E14/#4-4的多形性标记F11进行PCR分析。结果2只小鼠均检测出所期待的扩增产物。
(2)Southern印迹分析(图9)
与(实施例2)相同,用人L1序列作探针,对E14/#4-7来源的1只小鼠(K#4-7-1:嵌合率约5%)。尾部的2μg基因组DNA进行Southern印迹分析。结果可以看到多数存在人L1序列,其带型与E14/#4-7相类似。与小鼠基因组的质量比约为E14/#4-7的10%。图9中所示,各道均使用经BglII消化了的2μg尾部基因组DNA。用32P标记的人L1序列作探针,通过图像分析仪BAS200(富士照片胶片公司)检测信号。左侧为分子量。各道从左依次为1:K#/4-7-1、2:空白、3:E14/#4-7。
(3)尾部纤维母细胞的G418耐性试验
嵌合体小鼠中,来源于E14/#4-7的1只(K#4-7-1:嵌合率约5%)、来源于E14/#4-4的1只(K#4-4-41“嵌合率约5%),按以下的方法从它们的尾部制备纤维母细胞。DNA的制备与(实施例4)相同,切嵌合体小鼠的尾部5mm~10mm,在PBS/1mM EDTA中洗涤数次后,过网除去切下的表皮,用网研细内部组织。将组织细片移入盛有5ml PBS/1mM EDTA的试管中,室温静置30分钟-1小时。其后余下1ml的PBS/EDTA,除去上清,加入1ml 0.25%胰酶/PBS,室温放置5-10分,边轻扣或边吹打边搅拌组织。1000rpm,离心10分钟,将沉淀悬浮到2ml的DMEM(10%FCS)中,接种到35mm培养皿中。培养7-10天后,用胰酶处理细胞使之脱离皿壁,接种到2块35mm培养皿,每瓶约104个细胞,其中1瓶中加入终浓度400μg/ml的G418,培养5~7天,观察每瓶中的活细胞。这种条件下,来源于野生型ICR小鼠的纤维母细胞在G418的存在下几乎100%死亡。结果在2只小鼠中均见到G418耐性纤维母细胞的存在。
这些结果表明,E14/#4-7、E14/#4-4在小鼠中贡献给各种正常组织,且保持人4号染色体部分片段。
(实施例9)将人14号染色体或其片段导入小鼠ES细胞
用(实施例1)中得到的保持人14号染色体的小鼠A9细胞株(以下称作A9/#14)作染色体供体细胞。用小鼠ES细胞株TT2(购自Lifetech Oriental公司,Yagi等,分析生化,214:70,1993)作受体细胞。TT2的培养方法参照(相沢慎一,生物学手册系列,8,基因打靶,羊土社,1995)的方法进行,饲养细胞选用丝裂霉素c(Sigma)处理的G418耐性原代培养细胞(购自Lifetech Oriental公司)。微细胞融合实验及G418耐药株的筛选与(实施例2)相同。耐药株的出现频率为每107个TT2细胞出现326个。耐药株的冻存、基因组DNA的制备与(实施例2)相同。
用γ射线照射微细胞可得到人14号染色体的片段(Koi等,科学,260:361,1993)。将从约108个A9/#14获取的微细胞悬浮剂5mlDMEM中,通过用Gammacell 40(同前)、在冰上照射30Gy的γ射线(1.2Gy/分×25分)。经γ射线照射的微细胞的融合以及耐药株的筛选与未经照射的微细胞相同,结果耐药株的出现频率为每107个TT2细胞出现3个。耐药株的冻存、DNA提取与(实施例2)相同。
根据下面(1)(2)的方法来证实未经照射的微细胞G418耐药株1-4、1-5,γ射线照射的微细胞G418耐药株3~1、3-2,4株中人14号染色体的保持。
(1)PCR分析(图10)
以耐药株基因组DNA为模板,通过PCR来检测人14号染色体上存在的基因((Genetic Maps)基因图谱,同前)和多形性标记(多形性STS引物对,BIOS社:D14S43,D14S51,D14S62,D14S65,D14S66,D14S67,D14S72,D14S75,D14S78,D14S81,PCI;自然,359:794,1992)的存在。下面是以GenBank、EMBL等的数据库获得的碱基序列为基础制备的基因引物-寡核苷酸序列。
NP(核苷磷酸化酶):5’-ATAGAGGGTACCCACTCTGG(序列号31),5’-AACCAGGTAGGTTGATATGG(序列号32)
TCRA(T-细胞受体α):5’-AAGTTCCTGTGATGTCAAGC(序列号33),5′-TCATGAGCAGATTAAACCCG(序列号34)
MYH6(心肌球蛋白重链):5’-TGTGAAGGAGGACCAGGTGT(序列号35),5),5’-TGTAGGGGTTGACAGTGACA(序列号36)
IGA2(免疫球蛋白α-2恒定区):5’-CTGAGAGATGCCTCTGGTGC(序列号37),5’-GGCGGTTAGTGGGGTCTTCA(序列号38)
IGG1(免疫球蛋白γ-1恒定区):5′-GGTGTCGTGGAACTCAGGCG(序列号39),5’-CTGGTGCAGGACGGTGAGGA(序列号40)
IGM(免疫球蛋白μ恒定区):5’-GCATCCTGACCGTGTCCGAA(序列号41),5′-GGGTCAGTAGCAGGTGCCAG(序列号42)
IGVH3(免疫球蛋白高链可变区):5′-AGTGAGATAAGCAGTGGATG(序列号43),5’-GTTGTGCTACTCCCATCACT(序列号44)
以4个耐药株的基因组DNA作模板,用与(实施例2)相同的方法对上述18种引物进行了PCR扩增,结果全部或部分引物得到所期待的扩增产物。用经γ射线照射的微细胞而得到的耐药株3-1、3-2才发现14号染色体的一部分区域有缺失倾向。另外,用未经照射的微细胞耐药株1-4株也有缺失现象。结果如图10所示。图10中左边为以人14号染色体的G带为基础制作的染色体模式图和一些位置明确的标记在G带的位置(参照实施例2)。根据目前获得的信息(参照实施例2),基因和多形性标记的排列表明其大致的位置关系,其顺序未必准确。用PCR对4种G418耐性TT2细胞克隆进行检测,能得到所期待扩增产物的标记用■表示,不能检测出的标记用□表示。A9/#22是染色体供体细胞。右端表示实施例11(1)的结果。
(2)荧光原位杂交(FISH)
参照(松原等,FISH实验方案,秀润社,1994)所记载的方法,应用人14号染色体特异的探针(CHROMOSOME PAINTINGSYSTEM,Cambio社)进行FISH分析。其结果是,对于4株在几乎所有观察到的分裂像中,能观察到人14号染色体或其部分片段是独立的染色体。所观察到的人染色体的相对大小与用PCR分析结果进行的推测一致。
通过以上实验可以证实所得G418耐药株1-4、1-5、3-1、3-2保持人14号染色体的全部或其部分片段。
(实施例10)由保持人14号染色体片段的ES细胞制备嵌合体小鼠
将在(实施例9)中得到的,经证实保持人14号染色体的4株G418耐性ES细胞株(1-4,3-1,3-2,1-5)的冻存株融化,注入到ICR或MCH(ICR)(日本CREA)雄雌小鼠进行交配而得到的8细胞期胚胎中,每个胚胎注入8-10个。用ES培养基(实施例9)培养一晚,形成胚泡后,移植到经伪孕处理2.5天后的代孕母亲ICR小鼠(日本CREA)的子宫中,每侧移入大约10个注入胚。结果如(表3)所示。
表3.由保持人14号染色体(片段)的TT2细胞株制备嵌合体小鼠

共移植了494个注入胚,诞生了64只子鼠。由宿主胚而来的毛色为野鼠色(深棕),其中若能见到由E 14细胞而来的浅灰色,则可判定为嵌合体。所生64只中具有明显浅灰色的,也就是说能识别出由E14细胞贡献的个体有8只。在来源于1-4的1小鼠中最高的贡献率为80%。
根据该结果可表明保持人14号染色体的小鼠ES细胞株(1-4,1-5,3-1,3-2)能形成嵌合体,即有分化或小鼠正常组织的能力。
(实施例11)在保持人14号染色体片段的ES细胞而来的嵌合体小鼠中证实人14号染色体片段的保持。
通过下面(1)~(3)的方法来证实(实施例10)中所得嵌合体小鼠中保持人14号染色体部分片段(1)用各组织的DNA进行PCR分析。
用与(实施例4)相同的方法,从嵌合体小鼠中来源于3-1的小鼠(K3-1-1:嵌合率约25%)的尾部制备基因组DNA。以此作模板,用(实施例9)中所示分析14号染色体时应用的引物中的能检测出3-1的全部14种引物进行PCR分析。结果所有14种引物均检测到预期的扩增产物(图10)。
此外,用Puregene DNA分离试剂盒从同一小鼠(K3-1-1)的脑、肾、脾、心、肝、胸腺提取基因组DNA,用IGM引物(实施例9)对各组织进行PCR分析。结果在所有的组织中发现预期的扩增产物(图11)。将PCR产物经2%琼脂糖凝胶电泳后,用溴化乙锭染色进行检测。图11中各泳道从左依次为B:脑,K:肾,Sp:脾,H:心脏,Th:胸腺,Pc:人纤维母细胞(HFL-1)DNA(阳性对照),nc:非嵌合体小鼠尾部DNA(阴性对照),M:标准(HindIII消化入DNA+HaeIII消化 X174DNA,宝酒造)。
X174DNA,宝酒造)。
(2)尾部纤维母细胞的G418耐性试验
用以下方法,从嵌合体小鼠中来源于3-2的小鼠2个(K-3-2-1:嵌合率约25%,K3-2-3:嵌合率约50%)、来源于1-4的1个小鼠(K1-4-1:嵌合率约80%)的尾部制备纤维母细胞。DNA制备与(实施例4)相同,从3~6周龄嵌合体小鼠的尾部取5~10mm,在PBS/1mM EDTA中洗数次后,过网除去切下的表皮,在网上研磨内部组织。将组织细片移到盛5ml PBS/1mM EDTA的试管中,室温静置30分~1小时。其后剩余1ml PBS/EDTA,去除上清,加入1ml 0.25%胰酶/PBS,室温放置5-10分钟,边轻扣或吹打边搅拌组织。1000rpm离心10分钟,将沉淀悬浮到2ml的DMEM(10%FCS)中,接种到35mm培养皿上。培养7~10天后,用胰酶处理细胞使之脱离皿壁,接种到4个35mm培养皿,每个皿中约104个细胞,在其中2皿中加入400μg/ml的G418,培养5-7天后,计算每皿中的存活细胞数。来源于野生型ICR小鼠的纤维母细胞,在G418存在的条件下几乎100%死亡。假如G418耐性纤维母细胞的增殖速度在2种条件下相同,那么选择培养基中存活的细胞数与非选择培养基上存活的细胞数的比率能够反映对G418耐性ES细胞株来源的纤维母细胞的纤维母细胞集落的贡献率。结果如(图12)所示,可看到3株中均存在G418耐性纤维母细胞。在图12中,耐性率是每个个体从2组选择/非选择35mm培养皿得到的值进行平均。ICR表示野生型ICR小鼠。
(3)尾部来源的G418耐药纤维母细胞的FISH分析
用与(实施例2)相同的方法,对上面(2)得到的G418耐药纤维母细胞(K3-2-3,K1-4-1来源)进行FISH分析。用FITC标记从HFL-1细胞(实施例1)提取的人总DNA(松原等,FISH实验方案,秀润社,1994),以此作探针。结果在2株细胞中均在几乎所有分裂像中观察到独立的人染色体部分片段。
这些结果证实,保持人14号染色体部分片段的TT2细胞株在小鼠中贡献给各种正常组织,并且其中保持人14号染色体部分片段。
(实施例12)将人2号染色体部分片段导入ES细胞
用(实施例1)中获得的保持人2号染色体部分片段的小鼠A9细胞W23(以下简称A9/#W23)作染色体供体细胞。用小鼠ES细胞株TT2(实施例9)作染色体受体细胞。微细胞融合及G418耐药株的选择与(实施例2)相同。耐药株的出现频率为每107个TT2细胞出现1-3个。耐药株的冻存、基因组DNA的制备与(实施例2)相同。通过下面(1)、(2)的方法证实耐药株5-1、5-2、5-3中保持有人2号染色体部分片段。
(1)PCR分析
以耐药株基因组DNA为模板,存在于人2号染色体上的基因(基因图谱,同前)中选择在染色体供给细胞A9/#W23中检测出的Cκ、FABPI进行PCR。
对各引物进行扩增,结果在3株中均检测出两个引物所期待的扩增产物。
(2)荧光原位杂交(FISH)
与(实施例2)相同,用人2号染色体特异的探针(CHROMOSOMEPAINTING SYSTEM,Cambio社)进行FISH分析。结果在3株几乎所有的分裂像中检测出人2号染色体部分片段是独立的染色体。其大小与在A9/#2W23中观察到的相同。
以上实验证实,所得G418耐药株保持有人2号染色体部分片段。(实施例13)由保持人2号染色体片段的ES细胞制备嵌合体小鼠
将(实施例12)中得到的、证实保持有人2号染色体部分片段的G418耐性ES细胞株5-1的冻存株融化,注入到ICR或MOH(ICR)(日本CREA)雄雌小鼠交配而得到的8细胞期胚胎中,每个胚胎注入10-12个细胞。用ES培养基(实施例9)培养一晚上形成胚泡后,移植到经伪孕处理的2.5天的代孕母亲ICR小鼠(日本CREA)子宫中,每侧注入约10个注入胚。
结果如(表4)所示。
表4.由保持人2号染色体(片段)的TT2细胞株制备嵌合体小鼠

共移植了264个注入胚,结果产生51只子鼠。根据毛色可判断嵌合体小鼠在来源于宿主胚(ICR)的白色中有可识别的来源于TT2细胞的野鼠色(浓茶)。所生51只小鼠中,毛色明显野鼠色,即可看出由ES细胞贡献的个体有18只。另外,最高的嵌合率为80%。
该结果证实,保持人2号染色体部分片段的G418耐性ES细胞(5-1)有形成嵌合体的能力,即有分化成小鼠正常组织的能力。(实施例14)在导入了人14号染色体的嵌合体小鼠血清中检测人抗体重链
用酶联免疫吸附实验(ELISA)来测定血清中人抗体的浓度。依照下面所描述的方法来进行ELISA。富山·安东,单克隆抗体实验手册,讲谈社,1987;安东·千叶,单克隆抗体实验操作入门,讲谈社,1991;石川,超敏感性酶免疫测定法,学会出版中心,1993;EdHarlow,Darid lane,抗体实验室手册,冷泉港实验室,1988;A.Doyle,J.B.Griffiths,细胞和组织培养:实验室方法,John Wiley&Sons Ltd.,1996。参照这些文献中描述的方法调整反应条件,反应在4℃,过夜进行。测定时要把抗人免疫球蛋白的抗体或抗原稀释为0.5-10μg/ml(100-5000倍),铺ELISA板,4℃,过夜。测定血清样品时,用补充了5%小鼠血清(Sigma,M5905)的PBS封闭。稀释样品和标记的抗体,在对杂交瘤培养上清进行测定的时候,用补充了1%胎牛血清的PBS。用PBS将嵌合体小鼠血清稀释20倍。洗涤后将铺层过的板封闭1小时以上。洗板后,加样品,培养30分钟。洗板后,加入稀释了100-5000倍的酶标记的抗人免疫球蛋白抗体温育1小时后,洗板,加底物液进行显色。另外,对于不同的测定系统基本上进行同样的操作步骤,但用生物素标记的抗体时,洗板后加入亲和素-酶复合体,培养后,洗涤,加入底物液。用(酶标仪)微板读数仪(Biotech,EL312e)测定吸光度。
从出生后29-35天的嵌合体小鼠(实施例10,K3-1-2,K-3-2-2,K3-2-2,K3-2-3)中采血,进行ELISA分析。将用50mM碳酸盐-碳酸氢盐缓冲液,pH9.6稀释的抗人IgM小鼠单克隆抗体(Sigma,16385)铺到96孔微滴度板上,加入用补充有小鼠血清(Sigma,M5905)的PBS稀释的血清样品。然后加入过氧化物酶标记的抗人IgM的羊抗体(Tago,2392),培养后加入ABTS底物(Kirkegaard&Perry Laboratories Inc.,506200),用405nm的吸光度测定酶活性。用纯化的人IgM抗体(CAPPEL,6001-1590)或人IgG抗体(Sigma,14506)作标准。用补充了小鼠血清的PBS对标准物进行梯度稀释。在测定人IgG时,将抗人IgG的羊抗体(Sigma,13382)固定到板中,用过氧化物酶标记的抗人IgG羊抗体(Sigma,A0170)进行检测。结果如(表5)所示。人IgM和IgG的被检测出。
将2ml溶于PBS的人血清白蛋白(HSA,Sigma,A3782)与佐剂(MPL+TDM乳液,RIBI免疫化学研究公司)混合制成抗原溶液。在出生后的第27天、34天、41天对保持人14号染色体片段的嵌合体小鼠(实施例10,K3-1-1,K3-2-1)进行3次免疫,每次用0.2ml的0.25mg/ml的抗原溶液。用ELISA对这些嵌合体小鼠的血清进行同样地分析。其结果如(图13、图14)所示。结果是用HSA免疫的嵌合体小鼠血清中,人抗体的浓度在免疫后上升,免疫后的第17天可在K3-1-1的血清中检测出人IgM18μg/ml和IgG2.6μg/ml。在未移入人染色体的小鼠血清中,人抗体的效价不明显。
表5.嵌合体小鼠血清中人抗体的浓度(ELISA)
(实施例15)从导入了人14号染色体的嵌合体小鼠中获取产生人抗体重链的杂交瘤
在(实施例14)中用人白蛋白免疫的嵌合体小鼠(K3-1-1,实施例14)出生后44天,取出其脾脏,与骨髓瘤细胞进行细胞融合,制作杂交瘤细胞。参照<安东,单克隆抗体实验操作入门,讲谈社(Scientific,1991)的方法,使用P3X63-Ag.8.653(购自大日本制药,05-565)作骨髓瘤细胞进行杂交瘤的制备。杂交瘤铺到10块96孔板中,培养1周后用ELISA方法对培养上清进行分析。同实施例14一样,将抗人IgM小鼠单克隆抗体(Sigma,I6385)固定到板中来进行ELISA,得到6个阳性克隆。另外,在50mM的碳酸盐-碳酸氢盐缓冲液,pH9.6中将抗原HSA配成5μg/ml的溶液,以每孔100μl加到ELISA板的全部孔中。用过氧化物酶标记的抗人IgA+IgG+IgM羊抗体(Kirkegaard&Perry Labrrotaries Inc.,04-10-17)进行检测。10块板中检测出1个有阳性克隆的。这个克隆是6个人IgM阳性克隆中的一个。将该克隆(H4B7)再进行培养,稀释培养上清,以HSA为抗原,与上面相同,用过氧化酶标记的抗人IgM羊抗体(Tago,2392)进行ELISA,可以看到随着培养上清稀释度的增加吸光度减少。而从培养人IgM(CAPPEL,6001-1590)得到的稀释为2μg/ml的样品中,吸光度较低,且与稀释度无关。这表明杂交瘤H4B7所产生的抗体是对HSA具有特异性的抗体(图15)。图15中,横轴为培养上清样品的稀释率,纵轴表示405nm处的吸光度。
(实施例16)G418耐性标记的人2号染色体片段对嘌呤霉素耐性的再标记
在100mm平皿中,用含有G418(800μg/ml)的选择培养基(10%FBS、DMEM)培养保持G418耐性标记的人2号染色体片段的A9细胞(W23)(参照实施例1、图1)。在转染前,用限制酶SaII(宝酒造)线性化含有嘌呤霉素耐性基因的质粒pPGKPuro(WHITEHEADINSTITUTE,Dr.Peter W.Laird提供)。用胰酶处理细胞,以5×106/ml的浓度悬浮到Dulbecco′s磷酸盐缓冲液(PBS)中,参照(实施例1),在10μg DNA存在的条件下,用Gene.Pulser(基因脉冲仪)(Bio-rad)进行电穿孔。以25μF的电容、用4mm距离的电穿孔细胞(实施例1)、在室温下施加1000V的电压。将电穿孔后的细胞接种到3-6块100mm平皿中。1日后换成含10μg/ml嘌呤霉素(Sigma,P-7255)和G418(800μg/ml)的双重选择培养基。2-3周后,将所生成的克隆200个左右收集为一组。将3组中每组的细胞培养到2-3个25cm2的培养瓶中,将形成的微细胞与在25cm2培养瓶中培养的小鼠A9细胞进行与实施例1相同的细胞融合。然后将融合细胞移入2块100mm平皿中,用含G418和嘌呤霉素的上述双重选择培养基培养,3组中有1组得到具有2种双重耐药性的克隆。该克隆中,嘌呤霉素耐性标记导入人2号染色体的可能性很高。
(实施例17)导入了人染色体的ES细胞中所导入的人染色体的加倍
用添加了高浓度G418的培养基培养保持G418耐性基因标记的人14号染色体片段的ES细胞(E14/#14-36),可得到染色体加倍了的ES细胞克隆(生物手册,8,基因打靶,羊土社,1985)。将未经丝裂霉素处理的G418耐性小鼠原代细胞(购自Lifetech Oriental)接种到100mm的平皿中作为饲养细胞。将E14/#14-36接种到该100mm平皿中,半日后换成G418浓度为16mg/ml的培养基。每1-2天换一次培养基,1周后将G418变为10mg/ml,继续培养,从生成的克隆中选出15个进行培养,用人14号染色体特异的探针(参照实施例9)对染色体进行FISH分析。结果在8个克隆中出现人14号染色体的加倍。
(实施例18)同时保持人2号和4号染色体部分片段的小鼠ES细胞的获得
以(实施例16)中得到的具有双重耐药性克隆中的PG1为供体细胞、野生型A9细胞为受体细胞进行微细胞转移试验,可以证实PG1所保持的人2号染色体部分片段标记有嘌呤霉素耐性基因。微细胞的获得以及与A9细胞的融合与(实施例1)相同。结果在微细胞融合10天后其出现59个G418耐性克隆。将这些克隆的培养液换为含8μg/ml嘌呤霉素的培养基,然后再培养3天,有45个(76%)克隆生存。由于微细胞法中,大多数情况下1个受体细胞中只能移入1条或少数几个染色体,所以两个耐性基因同时以高频率转移表明PG1中所保持的G418耐性标记的人2号染色体部分片段标记有嘌呤霉素耐性基因。此外,为了检测人2号染色体部分片段上各个标记基因,分别以pSTneoB(实施例1)、pPGKPuro(实施例16)为探针对A9/#2W23(只对G418耐性,实施例16)和PG1进行了FISH分析(松原等,FISH实验方案,秀润社,1994)。结果在A9/#2W23中,在(实施例12)所证实的人2号染色体部分片段的相应姊妹染色体上分别有1个信号,共观察到2个信号,这表明pSTneoB插入到人2号染色体部分片段上的1个位点。在PG1中,在同等大小的染色体片段上观察到4个信号。pSTneoB和pPGKPuro的载体部分具有相同的序列,用pPGKPuro探针也可检测出pSTneoB。因此认为在PG1中所观察到的4个信号,其中2个为pSTneoB,另外2个是来源于pPGKPuro的信号。由此可以表明,PG1所保持的人2号染色体部分片段标记有G418耐性、嘌呤霉素耐性两种标记。
为了获得同时具有人2号、14号染色体部分片段的小鼠ES细胞,就以PG1细胞作为染色体供体细胞。用已保持有人14号染色体部分片段G418耐性TT2细胞1-4(实施例9)作染色体受体细胞。微细胞融合实验及嘌呤霉素耐药株的选择,除嘌呤霉素的浓度为0.75μg/ml外,其它与(实施例9)中G418耐药株的选择相同。该结果中嘌呤霉素耐药株的出现频率为每107个1-4细胞出现3~7个。由此可以说明这些嘌呤霉素耐药株在G418以300μg/ml存在的情况下只能增殖,并同时保持了G418耐性。双重耐药株的冻存、基因组DNA的获得与(实施例2)相同。参照下面(1)的方法来证实双重耐药株PG5、PG15、PG16中保持人2号和14号染色体部分片段,而PG15也可参照(2)进行。
(1)PCR分析
以双重耐药株的基因组DNA为模板进行PCR扩增。在人2号、14号染色体上的基因(基因图谱,同前)中,应用(实施例12;A9/#2W23)中所证实的存在于2号染色体的引物以及在(实施例9;TT2/#141-4)中所证实的存在于14号染色体上的引物。在3株细胞中,所有的引物均有期待的扩增产物。
(2)荧光原位杂交(FISH)
与(实施例11)相同,用FITC标记的人总DNA作探针进行FISH分析。结果几乎在所有的分裂相,可观察到大、小两个染色体片段。与(实施例9;TT2/#141-4)中由人14号染色体特异的探针所确定的部分片段的大小相同,而小的与(实施例12;TT2/#25-1)中由人2号染色体特异的探针所确定的部分片段的大小相同。(图16)为其结果。图中亮度低的染色体为小鼠的染色体,由于FITC荧光染色而呈现高亮度的大小2个染色体片段(箭头所示)是人的,认为分别是人14号、2号染色体部分片段。
以上的实验证实所得到的双重耐药性ES细胞同时保持人2号、14号染色体部分片段。
(实施例19)用同时保持人2号、14号染色体部分片段的ES细胞株制备嵌合体小鼠
将(实施例18)中得到的、经证实保持人2号、14号染色体部分片段、具有G418、嘌呤霉素双重耐药性的TT2细胞株PG5、PG15、PG16的冻存株融化,注入到ICR或MCH(ICR)(日本CREA社)雄雌小鼠交配而得到的8细胞期胚胎中,每个胚胎注入10-12个细胞。用培养ES细胞的培养基(实施例9)培养1晚上形成胚泡后,移植到假孕处理2.5天后代孕母亲ICR小鼠(日本CREA社)的子宫中,每侧子宫大约移植10个注入胚。
结果如(表6)所示。
表6.用同时保持人2号和14号染色体部分片段的ES细胞株制备嵌合体小鼠

共移植了551个注入胚,结果产生了73只子代小鼠。可根据毛色,由宿主胚(ICR)产生的白色中有TT2细胞来源的野鼠色(深棕)来判断嵌合体的左右。73只后代中毛色明显呈野鼠色的,也就是说能够识别由ES细胞贡献的个体有23只。
该结果表明,保持人2号和14号染色体部分片段的ES细胞株(PG5、PG15、PG16)具有形成嵌合体的能力,即保持有分化成小鼠正常组织的能力。
(实施例20)从同时保持人2号和14号染色体部分片段的ES细胞来源的嵌合体小鼠血清中检测人抗体。
将溶于PBS的人血清白蛋白(HSA,Sigma,A3782)与佐剂(MPL+TDM乳液,RIBI)混合,配成0.25mg/ml的HSA溶液,用它来免疫(实施例19)中制备的嵌合体小鼠KPG-15(9周龄,来源于PG5,嵌合率10%)、KPG-18(5周龄,来源于PG5,嵌合率10%)2只小鼠,每只0.2ml。免疫前及免疫8天后,从嵌合体小鼠采血,用ELISA方法(参照实施例14)检测血清中的人抗体μ链和人抗体κ链。将用50mM碳酸盐-碳酸氢盐缓冲液,pH9.6稀释的抗人抗体κ链的羊抗体加到96孔微滴度板中,加入血清样品,然后加入生物素标记的抗人抗体κ链的羊抗体(VECTORLABORATORIES,INC.,BA-3060),培养,再加入生物素化的辣根过氧化物酶和亲和素DH 的复合体(VECTORLABORATORIES,INC.,Vectastain ABC Kit PK 400),培养后加入底物3,3′,5,5′四甲基联苯胺(TMBZ,住友Bakelite ML-1120T),用450nm的吸光度检测酶活性。用纯化的含有κ链的、已知浓度的人IgG(Sigma,I-3889)作标准,用补充有小鼠血清的PBS对标准物进行梯度稀释。μ链的检测是这样的,将用50mM碳酸盐-碳酸氢盐缓冲液,pH9.6稀释的抗人抗体μ链的小鼠单克隆抗体(Sigma,I-6385)加到96孔微滴度板中,加入血清样品,然后加入过氧化酶标记的抗人μ链的小鼠抗体(The Binding Site Limited,MP008),培养后加入TMBZ(住友Bakelite,ML-1120T),用450nm的吸光度测定酶活性。纯化的含有μ链的浓度已知的人IgM(CAPPEL,6001-1590)作标准,用补充了小鼠血清(Sigma,M5905)的PBS进行梯度稀释。结果在免疫前的2只小鼠中均检测出人抗体μ链和κ链,其在血清中的浓度随免疫后上升(表7、表8)。
表7.嵌合体小鼠KPG15中人抗体的浓度(ELISA)

该结果表明由保持人2号和14号染色体部分片段的ES细胞而来的嵌合体小鼠中,人抗体重链和轻链基因能发挥功能。
(实施例21)在导入了人14号染色体片段的嵌合体小鼠血清中检测抗HSA的人抗体γ链
用(实施例10)中的方法制备保持人14号染色体片段的嵌合体小鼠(K9、K11,均来自TT2细胞株3-2,嵌合率分别为50%、30%),在小鼠出生后第79天、93天、107天、133天,用与(实施例20)相同的方法,用HSA进行4次免疫(K9),或在出生后74天、88天、111天,对(K11)进行3次免疫。用ELISA方法(参照实施例14)来检测嵌合体小鼠血清中含有抗人血清白蛋白的人γ链的抗体。将用50mM碳酸盐-碳酸盐缓冲液,pH9.6稀释的HSA(Sigma,A3782)加到96孔微滴度板中,加入样品,然后加入过氧化酶标记的抗人IgG小鼠抗体(Pharmagen,08007E),培养后,加入过氧化酶的底物液邻苯二胺(OPD,住友Bakelite,ML-11300),用490nm的吸光度测定酶活性。用HSA免疫的嵌合体小鼠血清中抗人HSA的人IgG的效价在免疫后增多。在对照ICR小鼠中,HSA免疫后,抗HSA人IgG的效价处于背景水平。结果如(图17)所示。图17中,横轴为用HSA免疫嵌合体小鼠后的天数,纵轴为490nm处的吸光度。该结果表明,在保持人14号染色体部分片段的嵌合体小鼠中,HSA抗原的刺激能引起对抗原特异性的人IgG抗体效价的增加。
(实施例22)在导入了人22号染色体片段的嵌合体小鼠血清中检测人抗体λ链
从9个月龄的嵌合体小鼠(实施例3,K22-7,嵌合率10%)采血,用ELISA方法检测血清中的人抗体λ链(参照实施例14)。将用50mM碳酸盐-碳酸氢盐缓冲液,pH9.6稀释的抗人抗体λ链的羊抗体(VECTOR LABORATORIES,INC.,A1-30%)加到96孔微滴度板中,加入血清样品,然后加入生物素标记的抗人抗体λ链的羊抗体(VECTOR LABORATORIES,INC.,BA-3070),培养,再加入生物素化的辣根过氧化物酶和亲和素DH的复合体(VECTORLABORATORIES,INC.,Vectastain ABC Kit,PK4000),培养,之后加入过氧化物酶的底物TMBZ(住友Bakelite,ML-1120T),用450nm的吸光度测定酶活性。用纯化的含有λ链的浓度已知的人IgG(Sigma,I-4014)作标准,用补充有小鼠血清的PBS进行梯度稀释。从嵌合体小鼠中可检测出相当于180ng/ml人IgG的人抗体λ链。该结果表明,保持人22号染色体的嵌合体小鼠中,人抗体λ链的基因能发挥功能。
(实施例23)在导入了人2号染色体片段的嵌合体小鼠血清中检测人抗体κ链
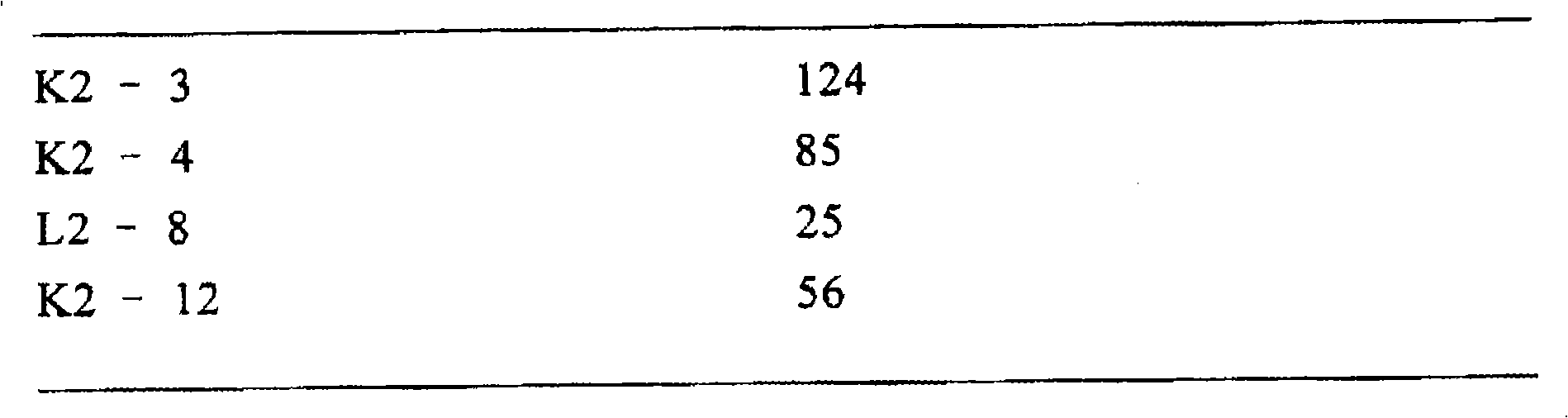
从5周龄的嵌合体小鼠(实施例13,K2-8,嵌合率为70%)和9周龄的嵌合体小鼠(实施例13,K2-3,K2-4,K2-12,嵌合率分别为50%、20%、80%)采血,用ELISA方法(参照实施例14)来检测血清中的人抗体κ链。将用50mM碳酸盐-碳酸氢盐缓冲液,pH9.6稀释的抗人抗体κ链的羊抗体(VECTORLABORATORIES,INC.,AL-3060)加到96孔微滴度板中,加入血清样品,然后加入生物素标记的抗人抗体κ链的羊抗体(VECTORLABORATORIES,INC.,BA-3060),培养,再加入生物素化辣根过氧化物酶和亲合物DH的亲合体(VECTOR LABORATORIES,INC.,Vectastain ABC Kit),培养后加入TMBZ(住友Bakelite,ML1120T),用在450nm的吸光度测定酶活性。用纯化的含有κ链的已知浓度的人IgG(Sigma,I-3889)作标准,用补充有小鼠血清的PBS进行梯度稀释,结果如(表9)所示。
表9.嵌合体小鼠中人抗体κ链的浓度(ELISA)
嵌合体小鼠 Igκ(mg/l)
用与(实施例20)相同的方法,在保持人2号染色体片段的嵌合体小鼠(实施例13、K2-3和K2-4)出生后的第66天、80天、102天,用HSA进行3次免疫。或者在嵌合体小鼠(K2-12)出生后的第13天、77天、91天、116天,用ELISA方法(参照实施例14)检测该嵌合体小鼠血清中抗HSA的人抗体κ链。将用50mM碳酸盐-碳酸氢盐缓冲液,pH9.6稀释的HSA(Sigma,A3782)铺到96孔微滴度板中,加入样品,然后加入生物素标记的抗人抗体κ链的羊抗体(VECTOR LABORATORIES,INC.,BA-3060),培养,由加入生物素化的辣根过氧化物酶和亲和素DH的复合体(VECTORLABORATORIES,INC.,Vectastain ABC Kit),培养后加入过氧化物酶的底物OPD(住友Bakelite,ML-11300),用490nm的吸光度测定酶活性。在HSA免疫的嵌合体小鼠血清中,抗HSA人κ链的效价在免疫后上升。而在对照ICR小鼠中,HSA免疫后抗HSA的人抗体κ链的效价处于背景水平。结果如(图18)所示。图18中,横轴为嵌合体小鼠经HSA初次免疫后的天数,纵轴为490nm处的吸光度。该结果表明,在保持人2号染色体部分片段的嵌合体小鼠中,人抗体κ链基因能发挥功能,而且HSA抗原的刺激能引起抗原特异性人Igκ抗体效价的上升。
(实施例24)由移入人14号染色体的嵌合体小鼠获得能产生人抗体重链(μ链或γ链)的杂交瘤。
在(实施例21)中用HSA免疫的嵌合体小鼠K9出生后第136天,取出其脾脏,与骨髓瘤细胞融合,制备杂交瘤。参照(安东·千叶,单克隆抗体实验操作入门,讲谈社Scientific,1997)的方法,用SP-2%-Ag14(大日本制药,05-554)作骨髓瘤细胞进行杂交瘤的制备。培养液中加入10%ORIGEN杂交瘤克隆因子(HCF,Bokusui.Brown),铺到8块96孔板中,3天后向培养液中加入1mg/ml的G418。培养1~3周后,用ELISA方法对培养上清进行分析(参照实施例14)。检测μ链时,将用50mM碳酸盐-碳酸氢盐缓冲液,pH9.6稀释的抗人μ链小鼠单克隆抗体(Sigma,I-6385)加到96孔微滴度板中,加入用PBS稀释的样品,然后加入过氧化酶标记的抗人μ链小鼠抗体(The Binding Site Limited、MP 008),培养后加入底物2,2′-连氮基-(3-乙基-苯并噻唑啉-6-磺酸)二铵盐(ABTS,Kirkegaard&Perry Laboratories Inc.,04-10-17),可检测出7个阳性孔。检测γ链时,将抗人γ链的小鼠单克隆抗体(Sigma,I-6260)铺到96孔微滴度板中,加入PBS稀释的样品,然后加入过氧化酶标记的抗人γ链小鼠抗体(pharmingen,08007E),培养后,用ABTS(Kirkegaard&Perry Labonatories Inc.,04-10-17)检测出2个人抗γ链阳性的孔。
(实施例25)用导入了人2号染色体的嵌合体小鼠制备能产生人抗体轻链的杂交瘤
在(实施例23)中用HSA免疫的嵌合体小鼠K2-3出生后第105天,取出其脾脏,与骨髓瘤细胞融合制备杂交瘤。参照(安东·千叶,单克隆抗体实验操作入门,讲谈社Scientific,1991)的方法,用P3X63Ag8.653(大日本制药,05-565)作骨髓瘤细胞进行杂交瘤的制备。将培养液中添加10%HCF(Bokusui Brown)铺到10块96孔板中,3天后向培养液中加入G418,1mg/ml,1~3周后用ELISA方法分析出现克隆孔的培养上清。ELISA方法与(实施例23)的相同,得到2个人抗体κ链特性的克隆。
(实施例26)G418耐性标记的人22号染色体对嘌呤霉素耐性的再标记
用与(实施例16)相同的方法,对保持G418耐性标记的人22号染色体的A9细胞(A9/#22γ2;实施例1中获得)进行人22号染色体的嘌呤霉素耐性的再标记。用pPGKPuro对γ2细胞进行电穿孔而得到双重耐药性克隆,每200个克隆收集到一组,这样3组(P1、P2、P3)作供体细胞,野生型小鼠A9细胞为受体细胞,进行微细胞转移。结果从P1得到6个双重耐药性克隆,从P2得到1个,从P3得到3个。在所得双重耐药性克隆中,以P3来源的6-1作微细胞供给细胞,野生型A9细胞为受体细胞进行微细胞转移实验,由此可以证实人22号染色体还标记有嘌呤霉素耐性基因(实施例18)。微细胞的获得和与A9细胞的融合,与(实施例11)中的方法相同。结果在微细胞导入11日后出现28个G418耐性克隆。将这些克隆的培养基换成含嘌呤霉素8μg/ml的培养基后,再培养3天,有21个(75%)克隆生存。微细胞法中,对于1个供体细胞多数情况下只能移入1条或少数几个染色体,所以两个耐性基因能同时、高频率导入表明6-1保持的G418耐性标记的人22号染色体还标记有嘌呤霉素耐性基因。
(实施例27)从产生人抗体重链的杂交瘤获得人抗体重链可变区的cDNA并测定其碱基序列
在(实施例15)中所得能产生人抗体重链(IgM)的杂交瘤中,从H4B7(HSA特异的)和H8F9(非特异的)用ISOGEN(日本基因)提取总RNA。用Ready-To-Go T-primed Ist strand Kit(Pharmacia),使用5μg的每个总RNA来合成cDNA。用下面所示的引物(参照Larrick等,生物/技术,7,934-,1989;Word等,国际免疫学,1,296-,1989制作)对所得cDNA进行PCR,扩增人抗体重链可变区。
CM1(人IgM恒定区):5′-TTGTATTTCCAGGAGAAAGTG(序列号45)
CM2(人IgM恒定区):5′-GGAGACGAGGGGG AAAGGG(序列号46)
HS1(人重链可变区域):5′-ATGGACTGGACCTGGAGG(AG)TC(CT)TCT(GT)C(序列号47)(8种的混合物)
HS2(人重链可变区域):5′-ATGGAG(CT)TTGGGCTGA(GC)CTGG(GC)TTT(CT)T(序列号48)(16种的混合物)
HS3(人重链可变区域):5′-ATG(AG)A(AC)(AC)(AT)ACT(GT)TG(GT)(AT)(GCT)C(AT)(CT)(GC)CT(CT)CTG(序列号49)(6144种的混合物)
*()は表示其位置的碱基是()内任一个形成的混合物。
H4B7、H8F9的第一轮PCR均用HS1×CM1、HS2×SM1、HS3×CM13种引物的组合物进行,(94℃1分、50℃2分、72℃3分,40个循环,Perkin-Elmer公司,热循环仪140),分别用HS1×CM2、HS2×CM2、HS3×CM2的引物对其PCR产物进行再次扩增(温度条件同前,30个循环)。扩增产物经1.5%琼脂糖电泳后,根据溴化乙锭染色进行检测。结果用HS3×CM2的引物时,从H4B7得到490bp的扩增产物。另外,对于H8F9,用HS3×CM2引物时,在相同位置得到一微量带,因此用该引物进行了再次扩增(温度条件同前,30个循环)。结果在扩增产物中检测出非常强的信号。参照石田等(基因表达实验手册,讲谈社Scientific,1995)的方法,将这些PCR产物在pBlueScriptll SK+(Stratagere)载体的Smal部位进行克隆。在插入了扩增产物的质粒中选择质粒#2、#3、#4(H4B7)、#11、#13、#14(H8F9),用自动荧光测序仪(Applyed Bio System社)检测扩增产物的碱基序列。将所得碱基序列和预计的氨基酸序列与已报道的人抗体VH区(Marks等,欧洲免疫学杂志,21,985-,1991)、JH区(Raretch等,细胞,27,583-,1981)的序列进行比较,结果表明H4B7、H8F9均含有VH4家族和JH的组合。该结果表明在保持人14号染色体部分片段的嵌合体小鼠中能产生有完整功能的人抗体重链蛋白质。
(实施例28)从表达人抗体κ链的嵌合体小鼠脾脏获取人抗体κ链cDNA并测定其碱基序列
用与(实施例5)相同的方法,从(实施例13)中获得的。在(实施例23)中证实能表达人抗体κ链的嵌合体小鼠K2-8脾脏中合成cDNA,应用下面的引物(参照Larrick等,生物/技术,7,934-,1989;Whitehurt等,核酸研究,20,4929-,1992制作)进行PCR,扩增人κ链可变区。阴性对照可应用K2-8的肝脏cDNA及(实施例10)的TT2/#143-2来源的嵌合体小鼠K3-2-2的脾脏cDNA。KC2(人Igκ链恒定区):5′-CAGAGGCAGTTCCAGATTTC(序列号50)KC3(人Igκ链恒定区):5′-TGGGATAGAAGTTATTCAGC(序列号51)KVMIX(人Igκ链可变区):5′-ATGGACATG(AG)(AG)(AG)(AGT)(CT)CC(ACT)(ACG)G(CT)(GT)CA(CG)CTT(序列号52)(3456种的混合物)
*()表示其结果的碱基是()内任何一种组成的混合物。
PCR的条件是,使用KVMIX×KC2、KKVMIX×KC3引物的组合物,以94℃15秒、55℃15秒、72℃20秒进行40个循环(Perlin-Elmer,热循环仪9600)。扩增产物经1.5%琼脂糖电泳后,用溴化乙锭染色进行检测。结果检测到所期望的约420bps(KC2)、450bps(KC3)长度的扩增产物。而在2种阴性对照中未检测出特异的扩增产物。参照石田等(基因表达实验手册,讲谈社Scientific,1995)的方法将这些扩增产物克隆到pBlue ScriptII SK+(Stratagene)载体的SmaI或EcoRV部位。在插入了扩增产物的质粒中选择来源于KVMIX×KC2的VK-#1克隆,用自动荧光测序仪(Applyed BioSystem)测定扩增产物的碱基序列。由于在所得到的碱基序列中,从人Igκ链的起始密码子到恒定区不含有终止密码子,所以认为克隆的扩增产物编码功能性人Igκ链的可变区。另外,已报道的人抗体Vκ区(Klein等,欧洲免疫学杂志,23,3248-,1993)、Jκ区(Whitehurst等,同前)的碱基序列相比较,结果表明γ-1克隆含有Vκ3家族、Jκ的组合。该结果表明在保持人2号染色体部分片段的嵌合体小鼠中,能产生完整功能的人抗体κ链蛋白质。
(实施例29)在保持人14号染色体片段的嵌合体小鼠血清中检测人抗体γ链亚类和μ链,并进行定量
从(实施例10)中出生后11周龄的嵌合体小鼠(K15A和K16A;来源于1-4,嵌合率为70、50%)采血,参照(实施例14),用ELISA方法检测血清中人抗体γ链亚类的浓度。
[人IgG1的测定]用PBS稀释抗人IgG抗体(Sigma,I-6260),铺到96孔微滴度板中。加入血清样品,然后加入过氧化酶标记的抗人IgG1抗体(Phamnigen,08027E),培养后加入7MBZ(住友Bakelite,ML-1120T),用450mm的吸光度测定酶活性。用补充了小鼠血清的PBS梯度稀释纯化的浓度已知的标准人IgG1(Sigma,I-3889)。
〔人IgG 2的测定〕用PBS稀释抗人IgG 2抗体(Sigma,I-9513),铺到96孔微滴度板中。加入血清样品,然后加入过氧化酶标记的抗人IgG抗体(Sigma,A-0170)培养后加入TMBZ(同上),用450nm的吸光度测定酶活性。用纯化的,浓度已知的人IgG 2(Sigma,I-4139)作标准,用补充有小鼠血清的PBS进行梯度稀释。
〔人IgG 3的测定〕用100mM盐酸-甘氨酸缓冲液pH 2.5稀释抗人IgG3抗体(Sigma,I-7260),室温放置5分钟后,用100mM磷酸盐缓冲液,pH 7.0稀释到10倍。铺到96孔微滴度板中。加入血清样品,然后加入过氧化酶标记的抗人IgG抗体(pharmingen,08007E),培养后加入TMBZ(同前),用450nm的吸光度测定酶活性。用纯化的,浓度已知的人IgG 3作标准(Sigma,I-4389),用补充有小鼠血清的PBS进行梯度稀释。
〔人IgG 4测定〕用100mM甘氨酸盐酸缓冲液pH 2.5,稀释抗人IgG 4抗体(Sigma,I-7635),室温放置5分钟后,用100mM磷酸盐缓冲液,pH 7.0稀释到10倍。铺到96孔微滴度板中。加入血清样品,然后加入过氧化物酶标记的抗人IgG抗体(pharmingen,08007E),培养后加入TMBZ(同前),用450nm的吸光度测定酶活性。用纯化的,已知浓度的人IgG 4(Sigma,I-4639)作标准,用补充有小鼠血清的PBS进行梯度稀释。
〔人IgM的测定〕测定μ链时,将PBS稀释的抗人μ链小鼠单克隆抗体(Sigma,I-6385)铺到96孔微滴度板中。加入血清样品,然后加入过氧化酶标记的抗人μ链小鼠抗体(The Binding Site limited,MP008),培养后,加入过氧化酶的底物TMBZ(同前),用450nm的吸光度测定酶活性。以纯化的,含有μ链的浓度已知的人IgM(CAPPEL,6001-1590)作标准,用补充有小鼠血清(Sigma,M 5905)的PBS进行梯度稀释。
结果如表10所示,在嵌合体小鼠K15A和K16A中,检测出IgG1、IgG2、IgG3、IgG4的全部亚类和IgM。
表10嵌合体小鼠中人抗体IgG亚类和IgM的浓度
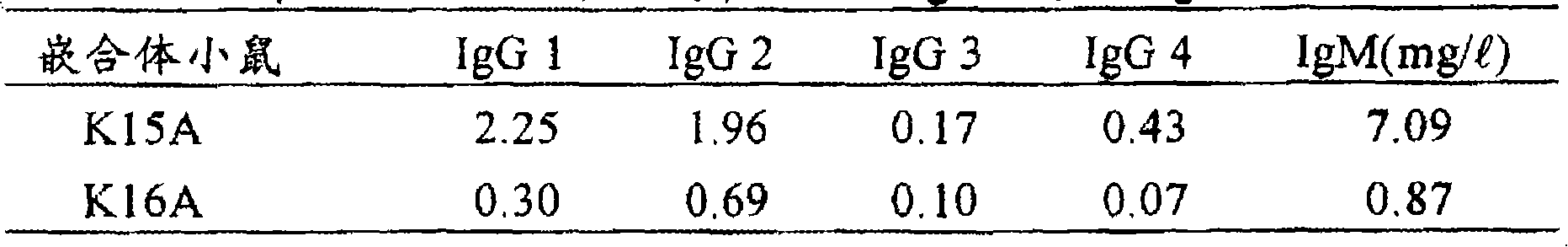
(实施例30)保持人22号染色体的小鼠ES细胞株(TT2)的制备
为了制备保持人22号染色体的小鼠ES细胞(TT2),用(实施例26)中制备的6-1(A9/#22,G418,嘌呤霉素耐性)细胞株作染色体供体细胞。用野生型TT2细胞株(实施例9)作染色体受体细胞。微细胞融合试验和嘌呤霉素耐药株的选择,除嘌呤霉素的浓度为0.75mg/ml外,其它的与(实施例9)中G418耐药株的选择一样。结果嘌呤霉素耐药株的出现频率为107个TT2细胞出现1~2个。嘌呤霉素耐药株的冻存、基因组DNA的制备与(实施例2)相同。根据下面(1)、(2)的方法来证实嘌呤霉素耐药株PG 22-1中保持有人22号染色体。
(1)PCR分析
以嘌呤霉素耐药株基因组DNA为模板进行PCR扩增。存在于人22号染色体的基因(基因图谱,同前),证实在(实施例2;A9/#22)中存在的有10种引物;用这些引物进行PCR扩增,结果检测出(实施例2;A9/#22)上存在的全部标记。
(2)Southern印迹分析
参照(实施例2)的方法,用人L1序列作探针,对来源于阴性对照野生型TT2、染色体供体细胞6-1、嘌呤霉素耐性TT2细胞株PG 22-1的基因组DNA进行Southern印迹分析。结果如(图19)所示。图中左侧为DNA分子量。由于PG 22-1中的带型与6-1的一致,且信号强度也相同,所以可以表明6-1细胞株中的22号染色体确实导入到PG 22-1中。
以上实验证明嘌呤霉素耐性TT2细胞株PG 22-1保持全部或部分人22号染色体。
(实施例31)用保持人22号染色体的小鼠ES细胞株(TT2)制备嵌合体小鼠
将(实施例30)中制备的,经证实保持有人22号染色体的嘌呤霉素耐药性TT2细胞株PG 22-1的冻存株融化,注入ICR或MCH(ICR)(日本CREA)雄雌小鼠交配而得到的8细胞期胚胎中,每个胚胎注入10~12个细胞。用(实施例9)中培养ES细胞用的培养基培养一晚上,形成胚泡后。移植到伪孕处理后2.5天的代孕母亲ICR小鼠(日本CREA)的子宫中,每侧子宫约移植10个注入胚。
结果如(表11)所示。共移植了266个注入胚,结果产生了36只子代小鼠。嵌合体的判断可依据毛色,在来源于宿主胚(ICR)的白色中能见到来源于TT2细胞的野鼠色(深棕)。在所生的36只子代小鼠中,毛色呈明显野鼠色的部分,即能够识别出由ES细胞贡献的有8只。
该结果表明保持人22号染色体的ES细胞株(来源于TT2,PG 22-1)有形成嵌合体的能力,也就是说具有分化成小鼠正常组织的能力。
表11用保持人22号染色体的TT2细胞株制备嵌合体小鼠

(实施例32)在保持人22号染色体的嵌合体小鼠的血清中检测人抗体λ链并进行定量
参照(实施例14),用ELISA方法对(实施例31)的嵌合体小鼠KPG 22-1~3血清中人抗体浓度进行定量。从出生后2个月的嵌合体小鼠采血,用ELISA方法检测血清中的人抗体λ链。将PBS稀释的抗人免疫球蛋白λ链抗体(VECTOR LABORATORIES,INC,BA-3070)铺到96孔微滴度板中,加入血清样品,然后加入生物素标记的抗人免疫球蛋白λ链抗体(VECTOR LABORATORIES,INC.,BA-3070),培养,再加入生物素化的辣根过氧化酶和亲合素DH的复合物(VECTOR LABORATORIES,INC.,Vectastain ABC kit),培养后加入TMBZ(同前),用450nm的吸光度测定酶活性。用纯化的含λ链浓度已知的人IgM(大日本制药,U13200)作标准,用补充有小鼠血清的PBS进行梯度稀释。结果如表12所示。该结果表明人抗体λ链基因保持人22号染色体的嵌合体小鼠中能起作用。
表12嵌合体小鼠中人抗体λ链的浓度
(实施例33)导入了人22号染色体的嵌合体小鼠血清中抗人HSA人抗体λ链的检测
与(实施例20)的方法相同,在嵌合体小鼠(实施例31,KPG 22-3)出生后的第79天、94天、110天,用HSA免疫3次。参照(实施例14),用ELISA方法检测血清中人抗体λ链。将50mM碳酸盐-碳酸氢盐缓冲液,pH 9.6稀释成5μg/ml的HSA(Sigma,A 3782)铺到96孔微滴度板中,加入血清样品,然后加入生物素标记的抗人免疫球蛋白λ链抗体(VECTOR LABORATORIES,Inc.,BA-3070),培养,再加入生物素化的辣根过氧化酶和亲合素DH的复合物(VECTORLABORATORIES INC.,Vectastain ABC kit),培养后,加入TMBZ(同前),用450nm的吸光度测定酶活性。用HSA免疫的嵌合体小鼠血清中抗HSA人λ链的效价在免疫后升高。而在对照的ICR小鼠中,HSA免疫后的抗HSA人抗体λ链的效价处于背景水平。结果如(图20)所示。图20中,横轴为嵌合体小鼠初次HSA免疫后的天数,纵轴为450nm处的吸光度。该结果表明保持人22号染色体的嵌合体小鼠中人抗体λ链基因起到作用,而且HSA抗原的刺激能引起抗原特异性人Igλ的效价上升。
(实施例34)用导入了人22号染色体的嵌合体小鼠制备产生人抗体轻链的杂交瘤
与(实施例25)的方法相同,在(实施例33)中用人白蛋白免疫的嵌合体小鼠KPG 22-3出生后第113天取出其脾脏,与骨髓瘤细胞融合,制备杂交瘤。参照<安东·千叶,单克隆抗体实验操作入门,讲谈社Scientific,1991>的方法,用SP-2/0-Ag 14(大日本制药,05-554)作骨髓瘤细胞进行杂交瘤的制备。在培养液中添加10%的HCF(Air Brown),加到5块96孔板中,培养1~3周,用ELISA方法分析出现克隆孔的培养上清。ELISA方法同(实施例33)相同,得到4个人抗体λ链的阳性克隆。
(实施例35)制备同时保持人22号和14号染色体部分片段的小鼠ES细胞株
为了制备同时保持人22号和14号染色体部分片段的小鼠ES细胞,用在(实施例26)中制备的6-1(A9/#22。G418,嘌呤霉素耐性)细胞株作染色体供体细胞。用已经保持人14号染色体部分片段的G418耐性TT2耐性株1-4(实施例9)作染色体受体细胞。微细胞融合实验和嘌呤霉素耐药株的选择除嘌呤霉素的浓度为0.75μg/ml外,其它与(实施例9)中G418耐药株的选择相同。结果嘌呤霉素耐药株的出现频率为每107个1-4细胞出现1~2个。这些嘌呤霉素耐药株在G418以300μg/ml存在的情况下仍能增殖,这表明它们同时具有G418耐性。双重耐药株的冻存、基因组DNA的制备与(实施例2)相同。根据下面的PCR分析来证实双重耐药株PG 22-5保持人22号和14号染色体部分片段。用双重耐药株的基因组DNA为模板进行PCR扩增。22号、14号染色体上存在的基因(基因图谱,同前)中,选择(实施例2;A9/#22)中证实的存在于22号染色体的引物和(实施例9;TT2/#141-4)中证实的存在于14号染色体上的引物进行扩增,结果检测出22号染色体上10种标记中的3种(D22S275,D22S315,Igλ),在TT2/#141-4中检测出全部存在于14号染色体上的标记。上面的实验表明所得到的双重耐药性TT2细胞株同时保持人22号和14号染色体的部分片段。(实施例36)用同时保持人22号和14号染色体部分片段的小鼠ES细胞株制备嵌合体小鼠
将(实施例35)中制备的,证实保持人22号和14号染色体部分片段的G418、嘌呤霉素双重耐药性TT2细胞株PG 22-5的冻存株融化,注入到ICR或MCH(ICR)(日本CREA)雄雌小鼠交配而得到的8细胞期胚胎中,每个胚胎注入10~12个。用(实施例9)中ES细胞用的培养基培养1晚,形成胚泡后,移植到经伪孕处理2.5天后的代孕母亲ICR小鼠(日本CREA)的子宫中,每侧子宫注入约10个的注入胚。
结果如(表13)所示。其移植了302个注入胚,结果产生了16只子代小鼠。可根据毛色来判断嵌合体,来源于宿主胚(ICR)的白色中有可识别的来源于TT2细胞的野鼠色(深棕)。所生16只子鼠中有5只毛色呈明显的野鼠色,即有5只是ES细胞贡献的个体。
该结果表明,保持人22号和14号染色体部分片段的ES细胞株(PG22-5)具有形成嵌合体的能力,即具有分化成小鼠正常组织的能力。
表13用保持人22号和14号染色体的TT2细胞株制备嵌合体小鼠

(实施例37)由同时保持人22号和14号染色体部分片段的ES细胞而来的嵌合体小鼠血清中人抗体λ链和人抗体μ链的检测和定量
用HSA免疫(实施例36)中的嵌合体小鼠(KPG 22-9、10和12)。KPG 22-9和KPG 22-10在出生后10周龄时进行免疫,免疫2周后采血。KPG 22-12在出生后7周龄和9周龄进行2次免疫,第2次免疫2周后采血。
参照(实施例14)用ELISA方法检测血清中含有人抗体μ链的抗体和含人抗体λ链的抗体以及含有人λ链及人μ链的抗体。
检测完整抗体时,将用PBS稀释的抗人免疫球蛋白λ链抗体(Kirkegaard&Perry Laboratories Inc.,01-10-11)铺到96孔微滴度板中,加入血清样品,然后加入过氧化酶标记的抗人免疫球蛋白μ链抗体(The Binding Site limited MP 008),培养后,加入过氧化物酶的底物TMBZ(同前),用450mm的吸光度测定酶活性,以纯化的,含有λ链浓度已知的人IgM(大日本制药,U13200)作标准,用补充有小鼠血清的PBS进行梯度稀释,与此相比较求出血清中人抗体的浓度。用ELISA,按照与(实施例29)和(实施例32)相同的方法对人抗体μ链和人抗体λ链进行检测和定量。结果如表14所示。在嵌合体小鼠中检测出λ链和μ链。也检测出含人抗体μ链并含λ链的抗体。该结果表明,在由保持人22号和14号染色体部分片段的ES细胞而来的嵌合体小鼠中,人抗体λ链基因和人抗体μ链基因同时起作用,并且在一部分B细胞中能产生重链和轻链均为人源性的完整抗体。
而在对照中,在只保持人14号染色体(实施例10)的嵌合体小鼠(K9),只保持人22号染色体(实施例31)的嵌合体小鼠(KPG 22-2)的血清中,含人抗体μ链或λ链的抗体浓度处于背景水平,这表明,在该检测系统中只检测出保持人λ链及μ链的完整抗体。
表14嵌合体小鼠中人抗体的浓度

(实施例38)在由同时保持人2号和14号染色体部分片段的ES细胞而来的嵌合体小鼠血清中检测人抗体(含有人κ链和μ链的抗体)
将溶于PBS的人血清白蛋白(HSA,Sigma,A3782)和佐剂(MPL+TDM乳液,RIBI免疫化学研究公司)混合,配制成0.75mg/ml的HSA溶液,用该溶液免疫(实施例19)中制备的嵌合体小鼠KPG-15(来源于TT2ES PG5,嵌合率10%),从出生后第2~3个月龄开始免疫,每次0.2ml,免疫3回,然后采血。另外从出生后6周龄的(实施例19)中的嵌合体小鼠KPG-26(来源于TT2ES PG6,嵌合率40%)采血。参照(实施例14)用ELISA方法检测血清中完整人抗体的浓度。将在PBS中稀释的抗人免疫球蛋白κ链抗体(Kirkegaard&PerryLaboratories Inc.01-10-10)铺到96孔微量滴度板中,加入用补充了小鼠血清(Sigma,M 5905)的PBS稀释的血清样品,然后加入过氧化物酶标记的抗人免疫球蛋白μ链抗体(The Binding Site limited,MP 008),培养后加入过氧化物酶底物TMBZ(住友Bakelite,ML-11207),用450mm处的吸光度测定酶活性,用纯化的,含有κ链,浓度已知的人IgM(CAPPEL,6001-1590)作标准,用补充有小鼠血清的PBS进行梯度稀释。用与(实施例20)相同的方法求出κ链,μ链的浓度。结果如表15所示。检测出既含人抗体μ链又含有κ链的抗体。而在对照中,即在只保持人14号染色体(实施例10)的嵌合体小鼠(K9),只保持人2号染色体(实施例13)的嵌合体小鼠(K2-9)的血清中既含有人抗体κ链又含有μ链的抗体浓度在0.002mg/ml以下,处于背景水平。该结果表明,在由同时保持人2号和14号染色体部分片段的ES细胞而来的嵌合体小鼠中,人抗体κ链基因和人抗体μ链基因能同时发挥功能,在一部分B细胞中能产生重链和轻链均为人源性的完整抗体。
表15嵌合体小鼠中人抗体的浓度

(实施例39)保持人2号染色体部分片段的小鼠ES细胞株(TT2F,X0)的制备
为了制备保持人2号染色体部分片段的小鼠ES细胞,用(实施例16)中制备的PG1细胞株作染色体供给细胞。用TT2F细胞(购自LifetechOriental公司)作染色体受体细胞,据报道该细胞株具有(39,X0)的染色体组型,在嵌合体小鼠中能有效地分化成卵母细胞(相尺慎一,生物手册,8,基因打靶,羊土社,1995)。微细胞融合及嘌呤霉素耐药株的选择除嘌呤霉素的浓度为0.75μg/ml外,其余用(实施例9)中G418耐药株的选择相同。结果嘌呤霉素耐药株的出现频率为每107个TT2F细胞出现5个。这些嘌呤霉素耐药株的冻存,基因组DNA的制备与(实施例2)相同。根据下面的PCR分析来证实耐药株P-20、P-21中保持有人2号染色体。以耐药株基因组DNA为模板,用存在于人2号染色体上的基因(基因图谱,同前)中,在(实施例1;A9/#2W23)中证实存在的引物Cκ、FABP1、Vκ1-2的3种进行PCR扩增,结果在2克隆株中,所有的三种引物均产生所期待的扩增产物。
以上实验表明,所得嘌呤霉素耐药株ES细胞株(TT2F,XO)保持有人2号染色体部分片段。
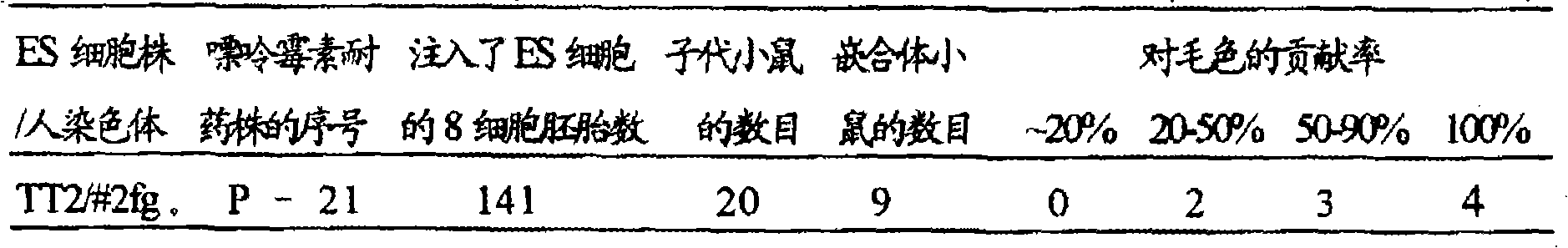
(实施例40)用保持人2号染色体部分片段的小鼠ES细胞株(TT2F,XO)制备嵌合体小鼠
将(实施例39)中制备的,证实含有人2号染色体部分片段的嘌呤霉素耐药性TT2细胞株P-21的冻存株融化,注入到ICR或MCH(ICR)(日本CREA)雄雌小鼠交配而得到的8细胞期胚胎中,每个胚胎注入10~12个细胞。用(实施例9)中ES细胞用培养基培养1晚上,形成胚泡后,移植到经伪孕处理2.5天后的代孕母亲ICR小鼠(日本CREA)的子宫中,每侧子宫注入大约10个注入胚。
其移植了141个注入胚,结果产生20只小鼠。可根据毛色来判断嵌合体,在来源于宿主胚(ICR)的白色中有可识别的来源于TT2细胞的野鼠色(深棕)。结果如(表16)所示。在所生的20只小鼠中毛色中有明显野鼠色部分,也就是说可以识别由ES细胞所贡献的个体有9只。其中有4只毛色完全呈野鼠色(ES细胞来源)。
该结果表明,保持人2号染色体部分片段的ES细胞株(P-21)具有形成嵌合体的能力,即具有分化成小鼠正常组织的能力。
表16用保持人2号染色体部分片段的TT2细胞株制备嵌合体小鼠
(实施例41)由保持人2号染色体部分片段的TT2F而来的嵌合体小鼠血清中人抗体κ链的检测及定量
从(实施例40)中出生后约1个月的嵌合体小鼠(来源于P-21嵌合率为100%,K2-1F~4F)采血,用与(实施例20)相同的ELISA方法对人抗体κ链的浓度进行定量。
结果如表17所示。结果表明当用TT2F ES细胞株时,人抗体κ链基因在嵌合体小鼠中能发挥作用。
表17嵌合体小鼠中人抗体κ链的浓度


(实施例42)在由保持人2号染色体部分片段的小鼠ES细胞(TT2F,X0)而来的嵌合体小鼠的后代中检测人染色体的保持
将(实施例40)中所得的雌性嵌合体小鼠中的K2-1F、K2-4F(二者毛色的嵌合率为100%)与雄性ICR小鼠进行交配,检测能否得到来源于ES细胞的后代,交配过程中用ICR雄性小鼠(白化,隐性)的精子使嵌合体小鼠的卵子受精,若卵子来源于TT2F细胞(野鼠色,显性)则产生野鼠色小鼠,若卵子来源于ICR则产生白化小鼠。分别经1次交配所得到的可生存子代小鼠(K2-1F:10只,K2-4F:5只)全部表现来源于ES细胞的野鼠色。从这些子代小鼠的尾部制备基因组DNA,通过PCR来检测人染色体片段的保持。用P-21(实施例39)中证实存在的3种引物进行PCR扩增,结果证实10只中有4只(K2-1F)、5只中有2只(K2-4F)存在P-21中检测出的3种引物。15只子鼠的PCR结果如(图21)所示。图中右侧为标记物(φX174,HaeIII片段,日本Gene)和主要带的分子量,左侧用箭头表示的是各个引物预期的扩增产物的长度。右侧也表示用阳性对照亲代嵌合体K2-1F、K2-4F尾部DNA的结果。这些结果表示保持人2号染色体部分片段的TT2F细胞株P-21在嵌合体小鼠中能分化成功能性卵子,并且将人2号染色体部分片段传递给由其卵子而来的后代。
(实施例43)在由保持人2号染色体部分片段的小鼠ES细胞株(TT2,XY)而来的嵌合体小鼠后代中检测人染色体的保持
将(实施例13)中得到的嵌合体小鼠中的K2-18(雄性,嵌合率70%)、K2-19L(雌性,嵌合率60%)及同胞的非嵌合体雌鼠进行混合交配,检测能否产生来源于ES细胞的后代。因为TT2细胞具有(40,XY)的染色体组型,所以在雄性嵌合体K2-18中有分化成功能性精子的可能。这种情况下嵌合体小鼠中来源于TT2细胞(野鼠色,显性)的精子使ICR(白色:隐性)来源的卵子受精可产生野鼠色子鼠。交配而产生的可生存的子代小鼠共110只,其中有10只显示来源于ES细胞的野鼠色。从这10只野鼠色子鼠中选7只,用其尾部制备基因组DNA,用PCR检测人染色体片段的保持。用证实在5-1株(TT2/#2fg.,实施例12)中存在的2种引物(Cκ,FABP1)和在(实施例1)中显示的Vκ1-2引物进行PCR扩增,结果证实7只中有2只全存在这3种标记。该结果显示,保持人2号染色体部分片段的TT2细胞株5-1在嵌合体小鼠中能分化成功能性的精子,并且可将人2号染色体部分片段传递给由其精子而来的后代。
(实施例44)嵌合体小鼠后代的血清中人抗体κ链的检测和定量
用ELISA方法对(实施例42)的嵌合体小鼠后代K2-1F-1~10F及K2-4F-1~5血清中人抗体κ链浓度进行定量。从出生后约4~6周龄的小鼠采血,用与(实施例20)相同的ELISA方法对血清中人抗体κ链进行检测。结果与(实施例42)中所得保持染色体的结果一起用(表18)表示。该结果证实嵌合体小鼠所生后代中的人抗体κ链基因也能发挥作用。
表18嵌合体小鼠后代中人抗体κ链的浓度

(实施例45)对导入了人14号染色体部分片段的嵌合体小鼠脾细胞的分析
参照下列文献所描述的方法用流式细胞术进行分析。日本生化学会编,新生化学实验讲座12,分子免疫学I-免疫细胞。细胞因子,1989,东京化学同仁;东京大学医科学研究所,制癌研究部编,细胞工程分册第8卷,新细胞工学实验方案,1991,秀润社;A.Doyle and J.B.Griffiths,“细胞和组织培养:实验室方法”John Wiley&Sons Ltd.出版,1996。从(实施例19)出生后约6个月龄的嵌合体小鼠(KPG06;来源于PG 16,嵌合率为30%)取脾,经氯化铵水溶液处理后,用含1%大鼠血清的PBS中异硫氰酸荧光素(FITC)标记的抗小鼠CD 45R(B220)抗体(Pharmingen,01124A)进行染色。洗涤后,与含5%小鼠血清的PBS中生物素标记的抗人IgM抗体(Pharmigen,08072D)或作为对照的生物素标记的抗人λ链抗体(Pharmingen,08152D)进行反应,之后用链霉亲和素藻红蛋白(Pharmigen,13025D)染色,用流式细胞仪(BD,FACSort)进行分析。结果如图22所示。图中横轴为人IgM,纵轴为CD45R(B220)。B细胞标记~CD45R阳性(FITC)且人IgM阳性(PE)的细胞群增加了4%,这证实在嵌合体小鼠中存在细胞表面表达了人抗体μ链的细胞。
(实施例46)用分别表达人抗体重链、κ链、λ链的嵌合体小鼠脾脏cDNA克隆人抗体基因的可变区并测定碱基序列
从在(实施例29)、(实施例23)、(实施例32)中证实分别表达人抗体重链、κ链、λ链的嵌合体小鼠K15A(来源于1-4株,按实施例10的方法制作)、K2-8(实施例13中制作)、KPG22-2(实施例31中制作)的脾脏提取RNA,用与(实施例5)相同的方法合成cDNA,用下面的引物进行PCR,分别扩增人抗体基因的可变区。从非嵌合体小鼠ICR取脾制备cDNA,用它作阴性对照。无参考文献的引物的制作是以GenBank等数据库中碱基序列为基础的。
K15A(重链)
恒定区用:HIGMEX 1-2:5′-CCAAGCTTCAGGAGAAAGTGATGGAGTC(序列号53)
HIGMEX 1-1:5′-CCAAGCTTAGGCAGCCAACGGCCACGCT(VH3BACK的第2轮PCR使用)(序列号54)可变区用:VH1/5BACK(59℃,35个循环,Marks等,欧洲免疫学杂志,21,985-,1991),VH4BACK(59℃,35个循环,Marks等,同前),VH3BACK(第1次PCR:59℃,35个循环,第2次PCR:59℃,35个循环,Marks等,同前)。
·K2-8(轻链κ)
恒定区用:KC2H:5′-CCAAGCTTCAGAGGCAGTTCCAGATTTC(序列号55)
可变区用:Vκ1/4BACK(55℃,40个循环,Marks等,欧洲免疫学杂志,21,985-,1991),Vκ2BACK(55℃,40个循环,Marks等,同前),Vκ3BACK(55℃,40个循环,Marks等,同前)。
·KPG22-2(轻链λ)
恒定区用:CλMIX(以下3种引物等摩尔比混合)
IGL 1-CR:5′-GGGAATTCGGGTAGAAGTCACTGATCAG(序列号56)
IGL2-CR:5′-GGGAATTCGGGTAGAAGTCACTTATGAG(序列号57)
IGL7-CR:5′-GGGAATTCGGGTAGAAGTCACTTACGAG(序列号58)
可变区用:Vλ1LEA1(55℃,40个循环,Williams等,欧洲免疫学杂志,23,1456-,1993),Vλ2MIX(55℃,40个循环,如前面Williams等的报道中,将Vλ2LEA1、Vλ2JLEAD等摩尔比混合。Vλ3MIX(55℃,40个循环,如前面Williams等的报道,将Vλ3LEA1、Vλ3JLEAD、Vλ3BACK4等摩尔比混合)。
PCR是这样进行的:各个恒定区域与可变区域(重链3种、κ链3种、λ链3种)引物的组合在94℃,15秒,各个可变区引物的退火温度15秒,72℃,20秒用各个可变区引物的循环数(使用Perkin-Elmer,热循环仪9600)。VH3BACK的第2轮PCR是将第1轮PCR的扩增产物与(HIGMEX1-1×VH3BACK)的引物组合进行再度扩增。全部的扩增产物经1.5%琼脂糖电泳后,用溴化乙锭染色进行检测。结果在所有的组合中检测出所期待的长度(重链,约470bp,轻链κ:400bp,轻链λ:约510bp)的扩增产物。除在阴性对照中,在全部相同位置粉末检测出特异的扩增产物。用Prep.A.gene(Bio-Rad)将所有的扩增产物从琼脂糖凝胶提取出,然后用限制性酶处理(重链:HindIII-Pst I,重链κ:Hind III-Pvull,轻链λ:Hind III-EcoR I),克隆到pUC 119(泵酒透)载体的Hind III,PstI部位(重链),Hind III,Hincll部位(κ链),Hind III,EcoRI部位(λ链)。用下面的引物(右侧表示克隆数)进行扩增所得到的扩增产物克隆到质粒中,用自动荧光测序仪(Applyed Bio System)测定扩增产物的碱基序列。
·HIGMEX1-2×VH1/5BACK:10克隆
·HIGMEX1-2×VH4BACK:8克隆
·HIGMEX1-2(2nd PCR,HIGMEX1-1)×VH3BACK:5克隆
·KC2H×Vκ1/4BACK:6克隆
·KC2H×Vκ2BACK:7克隆
·KC2H×Vκ3BACK:4克隆
·CλMIX×Vλ1LEA1:5克隆
·CλMIX×Vλ2MI X:6克隆
·CλMIX×Vλ3MI X:5克隆
用DNASIS(日立软件工程公司)分析所得碱基序列,结果表明所有的序列来自人,它们是从起始密码子到恒定区域间不包含终止密码子的功能序列,所有的κ链、λ链、23种重链中的21种中均如此。在所确定的序列中去除同一序列后,可确定各不相同的可变区序列:重链为17种,κ链11种,λ链12种。
(实施例47)从分别表达人抗体重链,κ链、λ链的嵌合体小鼠脾脏制备cDNA,用它分析人抗体基因可变区的碱基序列
在以下几个方面分析(实施例46)中所确定的碱基序列(重链17克隆、κ链11克隆、λ链12克隆)。
1.鉴定各可变区序列中使用的已知胚系V基因区段
2.鉴定各可变区序列中使用的已知胚系J基因区段
3.鉴定重链可变区中使用的已知胚系D区段
4.在1、2、3结果的基础上鉴定重链可变区中N领域的付加
5.确定从各可变区序列推断出的氨基酸序列
结果如(表19)所示。1、2的鉴定是通过用DNASIS检素Genbank中登记的同源胚系V及J区段。分别参照(Cook等,Nature genetics7,162-1994),(Klein等,欧洲免疫学杂志,23,3248-,1993)、(Williams等,同前)所描述的方法,VH区段、V κ区段。Vλ区段和各V区段家族的名称一起列在表中。在3中,用DNASIS检素(Ichinhara等,The EMBO J.,7,13,4141-,1988)报道中的同源胚系D区段,以连续8bp以上的一放性为标准来确定,结果如表中所示。认为DN*是(Green等,Nature Genetics,7,13-1994)报道中的新DN*家族。4中,在1(V)、2(J)、3(D)结果的基础上,将在任何胚系区段中都不出现的碱基序列确定为N区。结果在确定为D区的13种顺序列中的11种中有N区,其平均长度为8.7bp。5中用DNASIS将各碱基序列变换为用单符号表示的氨基酸序列。表中只表示了CD3区。表中右侧为克隆各可变区时使用的引物名称及各克隆名称。
表19
(实施例48)制备用于敲除掉TT2(或TT2F)的细胞的抗体基因(重链、软链κ)靶向载体
小鼠抗体基因(重链、软链κ)破坏后,可将G418耐药基因标记的人14号染色体片段(实施例9)和嘌呤霉素耐药基因标记的人2号(实施例18)或22号染色体(实施例35)导入TT2(或TT2F)细胞。这样参照(主链+κ链:实施例19,主链+λ链:实施例36)的方法,应用导入的人14号+2号或人14号+22号染色体、小鼠抗体基因(主链、转链κ)基因破坏了的TT2(或TT2F)的细胞,可制成嵌合体小鼠,期望它能产生重链和轻链主要来源于人的抗体。以下表示图23~27中限制性酶的简称。
限制性酶:Kp:KpnI、B:BamHI、RI:EcoRI、RV:EcoRV、N:NotI、SII:ScaII、Sca:ScaI、Sfi:SfiI、Sm:SmaI、X:XhoI、SI:SaII、dKp:KpnI的删除、(X):λ载体的XhoI酶切位点。
点线部分:pBluescript SKII(+)质粒DNA。
1.制备G418耐药基因两侧插入了LoxP序列的LoxP-pstNEO质粒
TT2(或TT2F)细胞的抗体基因被除掉以后,为了去除G418耐药基因,有必要在G418耐药基因(实施例1)的两端同向插入Cre重组酶(Sauer等,美国国家科学院院刊,85,5166-,1988)的识别序列LoxP序列(Sauer等,同前)。用限制酶XhoI把pstNEO基因从pSTneoB质粒DNA(实施例1)切出,用琼脂糖凝胶电泳纯化DNA片段,用T4DNA聚合酶(Takara社制Blunting end kit)将两端平端化。含LoxP序列的质粒DNA pBS246(质粒pBS246,LoxP2盒式载体,U.S.专利4959317)购自GIBCO BRL公司。在该质粒的EcoRI和SpeI切断位点插入XhoI接头DNA。变成XhoI的识别序列。将上述pstNEO DNA片段插入这一改变的pBS246的EcoRV酶切位点,得到质粒LoxP-pstNEO(图23)。
2.包含来源于C57BL/6的抗体重链Cμ(IgM恒定区)、轻链Jκ-Cκ(Igκ连结区域和恒定区)的基因组DNA克隆的分离
由于TT2(或TT2G细胞)来源于C57BL/6小鼠和CBA小鼠的F1代小鼠,本发明者打算用来源于C57BL/6小鼠的基因组DNA克隆制备敲除掉抗体基因用的载体。用来源于Clontech公司成熟C57BL/6N雄性小鼠肝脏的基因组DNA文库作基因组DNA库。用以下的合成DNA序列(60聚体)作筛选用的探针。
重链Cμ探针:5′-ACC TTC ATC GTC CTC TTC CTC CTG AGC CTCTTC TAC AGC ACC ACC GTC ACC CTG TTC AAG-3′(序列号59)轻链κ探针:5′-TGA TGC TGC ACC AAC TGT ATC CAT CTT CCCACC ATC CAG TGA GCA GTT AAC ATC TGG AGG-3′(序列号60)。
分析分离出的λ克隆,将含有重链Cμ或轻链Jκ-Cκ的DNA片段亚克隆到pBluescript SKII(+)质粒(Stratagene公司)中(重链Cμ:图24;轻链Jκ-Cκ:图25)。用这些DNA片段制备破坏以下TT2(或TT2F)ES细胞中的小鼠抗体基因的靶向载体。
3.制备破坏小鼠抗体重链基因用的载体-质粒
在编码含有2中制备的小鼠抗体重链恒定区的基因组DNA Cμ的区域内,用1中制备的LoxP-pstNEO基因替换含有第2~第4外显子的DNA片段(BamHI~XhoI)(图26)。pstNEO的转录方向与抗体基因的转录方向相反。用大肠杆菌JM109扩增该质粒DNA,通过氯化铯平衡离心进行纯化(细胞工学实验操作入门,1992年讲谈社刊)。用限制性酶SacII在一个位点酶切纯化的质粒DNA,用于TT2(或TT2F)ES细胞的转染。为了从转化体TT2(或TT2F)ES细胞检测抗体重链部分与靶向载体间同源重组而形成的克隆,用Cμ上游存在的转换区DNA片段(约500对碱基)作转化体基因组DNA Southern印迹用的探针。该DNA片段是在以下条件用PCR扩增129小鼠基因组DNA而得到的。
有义引物:5′-CTG GGG TGA GCC GGA TGT TTT G-3′(序列号61)
反义引物:5′-CCA ACC CAG CTC AGC CCA GTT C-3′(序列号62)
模板DNA:EcoRI消化的129小鼠基因组DNA 1μg
反应缓冲液:脱氧核苷混合物、Taq DNA聚合酶,均为Takara公司产品。
反应条件:94℃,3分,1次→94℃,1分;55℃,2分;72℃,2分;3次→94℃,45秒;55℃,1分;72℃,1分;36次。
证实所扩增的DNA能按照Genbank的数据库,用H限制性酶HindIII在一位点酶切后,将它亚克隆到pBluescript质粒的EcoRV酶切部位。用限制性酶BamHI和XhoI酶切该质粒DNA(S8),用琼脂糖凝胶电泳纯化PCR片段(约550碱基对),用来作探针。用EcoRI和XohI消化靶向载体转化的TT2(或TT2F)ES细胞的基因组DNA,经琼脂糖凝胶电泳后,分离,进行Southern印迹,用上述探针进行检测。4.制备破坏小鼠抗体基因轻链κ用的载体
将2中制备的含小鼠抗体轻链κJ区及恒定区的基因组DNA的含J区(J1-J5)的DNA片段(EcoRI~SacII)用1中制备的LoxP-pstNEO基因替换(图27)。pst NEO的转录方向与抗体基因的转录方向相同。用大肠杆菌JM 109扩增该质粒DNA,通过氯化铯平衡离心进行纯化。用限制性酶KpnI在一个位点酶切纯化的质粒DNA,用以转染TT2(或TT2F)ES细胞。为了从转化体TT2(或TT2F)ES细胞检测抗体重链部分与靶向载体进行同源重组而形成的克隆而进行转化体基因组DNA Southern印迹分析时用轻链Jκ-Cκ基因组DNA(参照图25)的3′端DNA片段(XhoI~EcoRI;约1.4k bp)作探针。用EcoRI和NotI消化靶向载体转化的TT2(或TT2F)ES细胞的基因组DNA,经琼脂糖凝胶电泳分离后,进Southern印迹分析,有上述的探针检测。
(实施例49)获取抗体基因破坏了的小鼠ES细胞株
为了获得抗体重链基因的同源重组体,用限制酶SacII(宝酒造)使(实施例48)中制备的抗体重链靶向载体线性化,参照(相尺慎一、生物学手册,8,基因打靶,羊土社,1995)的方法,导入小鼠ES细胞TT2F。用胰酶处理TT2F细胞,以2.5×107/ml的浓度悬浮到HBS中,加入5μg DNA,用基因脉冲仪(Bio-Rad,未连接电阻器装置)进行电穿孔。用4mm距离的电穿孔细胞,在960μF、250V的条件下于室温进行。将经过电穿孔的细胞悬浮到20ml的ES培养基中,接种到2块预先铺有饲养细胞的100mm组织培养用的塑料平皿中(Corning)。同样用10,15μgDNA进行实验。1天后换成含300μg/ml的G418(GENETICIN,Sigma)的培养基。7-9天后,挑选生成的176个克隆,分别在12孔培养板中进行增殖至汇合,将其4/5悬浮到0.2ml的保存用培养基(ES培养基+10%DMSO(Sigma))中,-80℃冻存。将剩余的1/5接种到12孔明胶板中,培养2天,用(实施例2)的方法提取基因组DNA。用限制性酶EcoRI和XhoI(宝酒造)消化这些G418耐药性TT2F细胞的基因组DNA,经琼脂糖凝胶电泳分离后,进行Southern印迹分析,用(实施例48)-3中所示的探针检测同源重组体。结果176株中得到3株同源重组体。野生型TT2F细胞和同源重组体#131、#141的Southern印迹分析结果如(图28)左侧第3列所示。用EcoRI、XhoI消化野生型TT2F细胞可检测出2条带(a、b),预期在同源重组体中这2条带均消失,出现新的下部的带(c)。结果图中#131、#141中a带消失,出现新的c带。图中左侧表示DNA的大小。结果表明这些克隆中抗体重链基因的一个等位基因被同源重组所破坏。
(实施例50)用抗体重链同源重组体ES细胞制备嵌合体小鼠
将(实施例49)中所得到的抗体重链同源重组体TT2F细胞株#131的冻存株融化,注入到ICR或MCH(ICR)(日本CREA)雄雌小鼠交配而得到的8细胞期胚胎中,每个胚胎注入10~12个。用ES细胞的培养基(实施例9)培养一晚上,形成胚泡后,移植到伪孕2.5天的代孕母亲ICR小鼠(日本CREA)的子宫中,每侧注入约10个注入胚。其移植了94个注入胚,结果产生了22只子鼠。根据毛色,来源于宿主胚(ICR)的白色中有可识别的来源于TT2F细胞的野鼠色(深棕),来判断嵌合体。所生22只子鼠中毛色中有明显呈野鼠色部分的,也就是说可识别由ES细胞所贡献的有18只。其中16个是毛色有80%以上野鼠色(来源于ES细胞)的雌性嵌合体小鼠。该结果表明,抗体重链同源重组体ES细胞株#131具有形成嵌合体的能力。由于所得嵌合体小鼠中贡献率非常多的个体多为雌性,表明ES细胞分化成机能性生殖细胞(卵子)的可能性较多。将嵌合体小鼠中贡献率为100%的2个雌性嵌合体,与MCH(ICR)雄性小鼠交配,结果所生子鼠全部为野鼠色。这些子鼠来自#131(参照实施例42),我们认为破坏的抗体重链等位基因以2∶1的比例进行遗传。
(实施例51)从抗体重链同源重组体制备双重破坏株
有这样的报道:因G418耐药基因的插入而破坏了一个等位基因的ES细胞株,在高浓度G418的培养液中进行培养,筛选高浓度G418耐药株,可得到2个等位基因同时破坏了的细胞株(相沢慎一等,生物学手册系列8,基因打靶,羊土社,1995)。基于此,为了得到TT2F抗体重链同源重组体#131、#141的2个等位基因破坏株,我们进行了以下实验。首先,为了测定#131、#141两株的G418致死浓度,以每个平皿约100个细胞的比例接种到10块35mm的平皿中(该例中使用未经丝裂霉素处理的G418耐性原代培养细胞作营养细胞,参照实施例9),在G418的浓度分别为0,0.5,1,2,3,5,8,10,15,20mg/ml(GENETICIN,Sigma)的ES培养基中培养10天。结果在3mg/ml时还能看到克隆,而到5mg/ml时则未见克隆形成。由此可确定最小致死浓度为5mg/ml,在浓度为4,5,6,7,8mg/ml的情况下筛选高浓度G418耐药株。将#131、#141分别以每块平皿约106个细胞的比例接种到10个100mm平皿中,在含有上述各浓度G418的ES培养基中进行培养(5个浓度,每个浓度两块平皿)。培养开始12天后在7mg,8mg/ml的平皿中挑选明确的克隆(#131:12株,#141:10株),用与(实施例49)相同的方法进行冻存、提取DNA。用限制性酶EcoRI和XhoI(宝酒造)消化这些高浓度G418耐药株的基因组DNA,经琼脂糖凝胶电泳分离后进行Southern印迹分析,用(实施例48)-3中所示的探针检测破坏了2个等位基因的克隆。结果从#131得到,株(#131-3)2个等位基因被破坏了的细胞株。对#131的6株进行Southern印迹分析的结果如(图28)所示。用EcoRI、XhoI进行消化,在野生型TT2F细胞中检测出2条野生型带(a、b)。在一个等位基因同源重组体(#131、#141)中上部的a带消失,出现新的c带(实施例49)。预期在2个等位基因被破坏的情况下,另一个野生型带b消失,只出现破坏型c带。在图中的3号(#131-3)克隆中可观察到这一带型。也就是说该克隆是抗体重链基因的2个等位基因均被破坏了的克隆。(实施例52)从抗体重链缺陷的纯合子TT2F细胞株中除去G418耐性标记基因
通过以下步骤除去(实施例51)中获得的抗体重链2个等位基因均被破坏了的细胞株(高浓度G418耐药株,#131-3)中G418耐药性标记基因,参照(相沢慎一,生物学手册系列8,基因打靶,羊土社,1995;高津圣志等,实验医学分册,免疫研究的基础技术,p255-,1995,羊土社)的方法,将含有能在G418耐药标记基因两端插入的LoxP序列(实施例48-1)间引起位点特异性重组的Cre重组酶基因的表达载体-PBS 185(BRL)导入#131-3。用胰酶处理#131-3细胞,以2.5×107/ml的浓度悬浮到HBS中,加入30μg的pBS 185DNA,用基因脉冲仪(Bio-Rad,未接电阻器装置)进行电穿孔。用4mm距离电穿孔细胞(实施例1),在960μF、250V的情况下进行。将经过电穿孔的细胞悬浮到5ml ES培养基中,接种到1块预先铺有饲养细胞的、60mm组织培养用塑料培养瓶中(Corning)。2天后用胰酶处理细胞,再次接种到预先铺有饲养细胞、100mm的培养瓶中,每个接种100、200、300个细胞,共接种3瓶。基因脉冲仪的设定条件只改变为(接连电阻器装置,电阻值无限大),其余同前。7天后,挑选长出的96个克隆,经胰酶处理后,分成2份,分别接种到2块铺有饲养细胞的48孔板中和只经凝胶处理的48孔板中。在含有300μ/ml G418(GENETIC、Sigma)的培养基中,将后者培养3天,根据其生存率来判断G418耐性。结果有6个克隆在G418存在的情况下死亡。将这些G418敏感株在35mm培养瓶中增殖至汇合,将其4/5悬浮到0.5ml保存时应用的培养基(ES培养基+10%DMSO<Sigma>)中,-80℃保存。将剩余的1/5接种到12孔凝胶铺过的板中,按(实施例2)的方法培养2天,制备基因组DNA。用限制性酶EcoRI(宝酒造)消化这些G418敏感性TT2F细胞株的基因组DNA,经琼脂糖凝胶电泳分离后进行Southern印迹分析,用包含G418耐药基因、来源于pSTneoB的3.2kb XhoI片段(探针A)来证实G418耐药基因的去除。其结果在所有的敏感株中均未检测出在#131-3中所观察到的与探针A杂交的带。由此可证实,在所得G418敏感株中,G418耐性标记基因确实被去除。此外,用同样的方法,以EcoRI消化pBS 185DNA的探针B进行Southern印迹分析,结果在这些G418敏感株中没有检测到与探针B杂交而形成的特异性带,这表明含有Cre重组酶的pBS 185未插入到敏感株的染色体中。也就是说,这些敏感株可与实施例48-4中所示由于敲除抗体轻链的载体(G418耐药基因的两端存在LoxP序列)进行转化。
(实施例53)将人14号染色体(抗体重链)导入抗体重链缺陷的ES细胞株
按(实施例9)的方法,用微细胞法将G418耐药基因标记的人14号染色体(含有抗体重链基因)导入(实施例52)中所得的内源性抗体重链的缺陷小鼠ES细胞株中(来于TT2F,G418敏感性)。用PCR分析等方法(实施例9)证实在所得G418耐药株中保持有包含人抗体重链基因的人14号染色体(片段)。
(实施例54)将人2号染色体片段或人22号染色体导入保持人14号染色体(片段)、抗体重链缺陷的ES细胞株中
按照(实施例18、35),用微细胞法,将嘌呤霉素耐药基因标记的人2号染色体(含有人抗体轻链κ基因)或人22号染色体(含有人抗体轻链λ基因)导入在(实施例53)中得到的、保持人14号染色体片段、抗体重链缺陷的小鼠ES细胞株(G418耐性)中。通过PCR分析等(实施例18、35)来证实在所得嘌呤霉素、G418双重耐药株中保持有人14号染色体(片段)和2号染色体片段或22号染色体(片段)。
(实施例55)用内源性抗体重链缺陷,保持含人抗体重链基因的14号染色体(片段)的ES细胞制作嵌合体小鼠
按照(实施例10)中的方法,用(实施例53)中获得的缺乏内源性抗体重链、保持含人抗体重链基因的14号染色体(片段)的ES细胞制作嵌合体小鼠。用(实施例14)的方法在所得嵌合体小鼠中检测来源于ES细胞株的B细胞所产生的人抗体重链。由于来源于ES细胞的B细胞中的功能性抗体重链基因只有导入染色体上的人源基因,所以大多数来源于ES细胞株的B细胞产生人抗体重链。
(实施例56)用保持人14号+2号、14号+22号染色体(片段)、内源性抗体重链缺陷的ES细胞制备嵌合体小鼠
参照(实施例19、36)的方法,用在(实施例54)中获取的保持人14号+2号、14号+22号染色体(片段)、缺乏内源性抗体重链的小鼠ES细胞制备嵌合体小鼠。用(实施例14、23、32)中所示的方法在所得嵌合体小鼠中来源于ES细胞株的B细胞中检测人抗体重链和轻链κ或轻链λ。与(实施例55)相同由于来源于ES细胞的B细胞中的功能性抗体重链基因只有导入染色体土的人源基因,所以多数来源于ES细胞的B细胞产生人抗体重链。另外,按如(实施例37、38)所示的方法,也检测出重链、轻链均来源于人的完整人抗体分子。
(实施例57)用保持人14号+2号、14号+22号染色体(片段)、内源性抗体重链缺陷的小鼠ES细胞来源的嵌合体小鼠制备产生人抗体的杂交瘤
与(实施例15、25、34)相同,用目的抗原免疫(实施例56)中制备的嵌合体小鼠,取脾,与骨髓瘤细胞进行细胞融合,制成杂交瘤。用ELISA法分析1~3周间的培养上清。按(实施例14、15、21、24、25、33、34、37、38)中所示的方法进行ELISA,可得到人抗体阳性以及人抗体阳性且对免疫抗原特异的克隆。
(实施例58)用抗体重链缺陷的纯合子小鼠ES细胞制备破坏了抗体轻链基因的细胞株
通过以下步骤制备同源重组体,该同源重组体是在(实施例52)中获得的抗体重链缺陷纯分子TT2F细胞株(G418敏感性)中又破坏了抗体轻链基因。用限制性酶KpnI(宝酒造)将(实施例48-4)中制备的抗体轻链靶向载体线性化,参照(相沢慎一、生物学手册系列8,基因打靶,羊土社,1995)的方法,导入到上述TT2F细胞株(G418敏感性)。7~9天后挑选所生克隆,按(实施例49)的方法冻存、提取基因组DNA。用限制性酶EcoRI和NotI(宝酒造)消化G418耐药株的基因组DNA,经琼脂糖凝胶电泳分离后进行Southern印迹分析,用(实施例48-4)中的探针检测同源重组体。
(实施例59)从抗体轻链同源重组体制备双重破坏株
依照下面程序,从(实施例58)的TT2F抗体轻链同源重组体(并且抗体重链缺陷的纯合子)制备破坏了轻链两侧等位基因的克隆。用(实施例51)的方法制备高浓度G418耐药株,冻存、提取DNA。用限制性酶EcoRI和NotI(宝酒造)消化高浓度G418耐药株的基因组DNA,经琼脂糖凝胶电泳分离后进行Southern印迹分析,用(实施例48-4)中的探针检测破坏了两侧等位基因的细胞株。
(实施例60)从抗体轻链缺陷的纯合子(并且抗体重链缺陷的纯合子)TT2F细胞株除去G418耐性标记基因
按(实施例52)中的步骤除去(实施例59)中抗体轻链两侧等位基因破坏株(高浓度G418耐药株)的G418耐性标记基因。将含有Cre重组酶基因的表达载体pBS 185(BRL)按(实施例52)的方法导入上述细胞株,其中Cre重组酶基因能在G418耐性标记基因两侧插入的LoxP序列(实施例48-1)间引起位点特异性重组。与(实施例52)相同,将所得G418敏感株在35mm培养瓶中进行增殖至汇合,将其4/5悬浮到0.5ml保存用培养基(ES培养基+10%DMSO<Sigmn>)中,-80℃冻存。将剩余的1/5接种到12孔铺有凝胶的板中,培养2天,按(实施例2)的方法提取基因组DNA。用限制性酶EcoRI(宝酒造)消化这些G418敏感性TT2F细胞的基因组DNA,经琼脂糖凝胶电泳分离后进行Southern印迹分析,用含有G418耐性基因、来源于pSTneoB的3.2kb XhoI片段作探针证实G418耐性基因的去除。
(实施例61)将人14号染色体(抗体重链)导入抗体重链、轻链缺陷的ES细胞株
参照(实施例9),用微细胞法将G418耐性基因标记的人14号染色体(含有人抗体重链基因)导入(实施例60)中内源性抗体重链和轻链缺陷的小鼠ES细胞株(来于TT2F,G418敏感性),用PCR分析等方法(实施例19)来证实在所得G418耐药株中保持有含人抗体重链基因的人14号染色体(片段)。
(实施例62)将人2号染色体(轻链κ)导入抗体重链、轻链缺陷且保持人14号染色体(抗体重链)的ES细胞株中
参照(实施例18),用微细胞法将嘌呤霉素耐性基因标记的人2号染色体(含人抗体轻链κ基因)部分片段导入(实施例61)中内源性抗体重链和轻链缺陷,且保持人14号染色体的小鼠ES细胞株(来源于TT2F,G418耐性)中。用PCR分析等方法(实施例18)来证实在G418双重耐药株中保持有人14号染色体(片段)和人2号染色体片段。
(实施例63)将人22号染色体(轻链λ)导入抗体重链、轻链且保持人14号染色体(抗体重链)的ES细胞株
参照(实施例35),用微细胞法将嘌呤霉素耐性基因标记的人22号染色体(含人抗体轻链λ基因)导入在(实施例61)中得到的内源性抗体重链和轻链缺陷且保持人14号染色体的小鼠ES细胞株中(来自TT2F、G418耐性)。通过PCR分析等方法(实施例35)在所得嘌呤霉素、G418双重耐药株中证实人14号染色体(片段)和人22号染色体(片段)的保持。
(实施例64)制备同时保持人2号(抗体轻链κ)、14号(抗体重链)、22号(抗体λ链)染色体或其片段、内源性抗体重链、轻链缺陷的小鼠ES细胞
为了得到同时保持3种人染色体的小鼠ES细胞,将杀稻瘟素耐性(Izumi等,实验细胞研究,197,229,1991)、潮霉素耐性(Wind等,细胞,82,321-,1995)等标记基因插入到人2号或22号染色体进行标记。按照(实施例16、26)的方法进行。用与(实施例9)相同的方法将杀稻瘟素耐性或潮霉素耐性标记的人22号染色体(含有人抗体轻链λ基因)导入在(实施例62)中得到的、内源性抗体重链和轻链缺陷且保持人14号染色体(片段)及2号染色体部分片段的小鼠ES细胞株中(来自TT2F,G418耐性、嘌呤霉素耐性)。使用分别与选择标记相适应的饲养细胞用于ES细胞的培养。使用潮霉素耐性标记的时候,选用保持、表达该标记的转基因小鼠的原代培养纤维母细胞(Johnson等,核酸研究,23卷,7期,1273-,1995)。用PCR分析等方法(实施例9,18,35)来证实所得G418、嘌呤霉素、潮霉素(或杀稻瘟素三重耐药株是否保持上述3种人染色体(片段)。用同样的方法将潮霉素或杀稻瘟素耐性基因标记的人2号染色体导入(实施例63)中所得的、内源性抗体重链和轻链缺陷且保持人14号染色体(片段)及22号染色体部分片段的小鼠ES细胞株(来自TT2F,G418耐性、嘌呤霉素耐性)。
(实施例65)从保持含人抗体基因(重链+轻链)的染色体(片段)、内源性抗体重链、轻链缺陷基因的小鼠ES细胞制备嵌合体小鼠
按照(实施例10)的方法,用(实施例61、62、63、64)中得到的保持有含人抗体基因的染色体(片段)、内源性抗体重链、轻链基因缺陷的小鼠ES细胞株制作嵌合体小鼠。用(实施例14、23、32)的方法检测所得嵌合体小鼠中,来自宿主胚的B细胞产生的小鼠抗体和来自ES细胞株的B细胞所产生的人抗体。由于来源于ES细胞的B细胞中,功能性抗体重链及轻链κ基因只有存在于导入染色体上的人源基因,所以来源于ES细胞的B细胞多数产生人抗体重链和轻链κ(Lonberg等,自然,368,856-,1994)。此外,用(实施例37、38)的方法检测重链、轻链均为人源性的完整人抗体分子。(实施例66)用来源于小鼠ES细胞的嵌合体小鼠制备产生人抗体的杂交瘤,其中ES细胞保持有含人抗体基因(重链+轻链)的染色体(片段)且内源性抗体重链、轻链基因缺陷。
与(实施例25)相同,用目的抗原免疫(实施例65)中得到的嵌合体小鼠,取其脾,与骨髓瘤细胞进行细胞融合,制作杂交瘤。用ELISA方法分析1-3周间进行培养的培养上清。ELISA方法按(实施例14、15、21、22、23、24、25、33、34、37、38)的方法进行,可得到人抗体阳性以及人抗体阳性且对免疫抗原特异的克隆。
(实施例67)用重链基因缺陷的宿主胚制备嵌合体小鼠
来源于(实施例49)中内源性抗体重链缺陷一侧等位基因的TT2F细胞株的嵌合体小鼠的后代中,呈野鼠色的个体,通过Southern印迹分析或PCR(实施例49)等方法选拔保持等位基因缺陷的个体(预期可能性为1/2)。抗体重链缺陷的杂合子雌雄个体进行交配产生子代小鼠,用Southern印迹分析(参照实施例49)来分析其淋巴细胞表示μ链的表达(Kitamura等,自然,350,423-,1991),可得到双侧等位基因缺陷、基本不能产生自身功能性抗体的抗体重链缺陷的纯合子(预期出现率为1/4,在膜型μ链缺陷小鼠中的结果:参照Kitamura等,自然,350,423-,1991)。在清洁环境中饲养的纯合体雌雄个体进行交配,由此而得到胚胎,可用这胚胎作制备嵌合体小鼠时的宿主。这种情况下制备的嵌合体小鼠中,功能性B细胞基本上来源于注入的ES细胞。RAG-2缺陷小鼠(Shinkai等,细胞,68,855-,1992)等不能制备自身功能性B细胞的其它小鼠品系也可用于此目的。在该系统中应用(实施例62,63,64)中得到的内源性抗体重链和轻链缺陷且保持人14号+2号、或14号+22号或14号+2号+22号染色体(片段)的小鼠ES细胞株,参照(实施例10)的方法制作嵌合体小鼠。所得嵌合体小鼠中,由来源于ES细胞的B细胞中功能性人抗体重链(14号染色体上)、轻链κ(2号染色体上)、轻链λ(22号染色体上)基因的表达而产生主要由人重链和人轻链组成的抗体。
(实施例68)导入了人14号染色体(片段)的ES细胞来源的嵌合体小鼠后代中人染色体的保持
按照(实施例9)的方法以及将(实施例9)中的小鼠ES细胞替换为TT2F(39,XO,实施例39)的方法得到保持人14号染色体(片段)的嵌合体小鼠,将它们与野生型ICR小鼠(白色,日本CREA)混合,交配。从所生野鼠色子鼠的尾部制备基因组DNA,通过PCR来检测人14号染色体片段的保持(实施例9、42、43)。如(实施例42)和(实施例43)所示,保持人14号染色体(片段)的小鼠ES细胞株在嵌合体小鼠中能分化成功能性卵子或精子,人14号染色体(片段)能遗传给其后代。也就是说可建立一种小鼠品系,它保持有含人抗体重链基因的人14号染色体(片段),并可遗传给后代。在本实施例以及下面的实施例69-74中,人14号染色体(片段)指含有人抗体重链基因的染色体(片段),人2号染色体(片段)指含有人轻链κ基因,人22号染色体(片段)指含有人抗体λ链基因的染色体(片段)。
(实施例69)导入了人22号染色体(片段)的ES细胞来源的嵌合体小鼠后代中人染色体的保持
用与(实施例30)相同的方法或稍作调整将(实施例30)的小鼠ES细胞替换为TT2F(39,XO)得到保持人22号染色体(片段)的嵌合体小鼠,将其与ICR小鼠混合,交配。从所生野鼠色子代鼠的尾部制备基因组DNA,用PCR法检测其中人22号染色体片段的保持(参照实施例42、43、30)。如(实施例42)和(实施例43)所示,保持人22号染色体或其部分片段的小鼠ES细胞株在嵌合体小鼠中能分化成功能性卵子或精子,并可将人22号染色体(片段)遗传给其后代。也就是说可建立一种小鼠品系,它保持有含人抗体轻链λ基因的22号染色体(片段),并能遗传给后代。
(实施例70)通过交配制备同时保持人2号染色体片段和14号染色体(片段)的小鼠
将(实施例42)或(实施例43)中得到的保持人2号染色体片段的小鼠品系与(实施例68)中得到的保持人14号染色体(片段)的小鼠品系进行交配,从所生子代小鼠的尾部制备基因组DNA,通过PCR方法等(实施例9、42、43)进行分析,得到同时保持人2号染色体部分片段和人14号染色体(片段)的小鼠。
(实施例71)通过交配制备同时保持人22号染色体(片段)和14号染色体(片段)的小鼠。
将(实施例69)中得到的保持人22号染色体(片段)的小鼠品系与(实施例68)中得到的保持人14号染色体(片段)的小鼠品系进行交配,从所生子代小鼠的尾部制备基因组DNA,通过PCR方法等(实施例30、42、43)进行分析,得到同时保持人22号染色体(片段)和人14号染色体(片段)的小鼠。
(实施例72)通过交配制备同耐保持人2号染色体片段,14号染色体(片段)和22号染色体(片段)的小鼠。
将(实施例71)中得到的保持人22号和14号染色体(片段)的小鼠品系与(实施例42、43)中得到的保持人2号染色体片段的小鼠品系进行交配,从所生小鼠的尾部制备基因组DNA,用PCR等方法(实施例9、30、42、43)进行分析,得到同时保持人22号和14号染色体(片段)及人2号染色体部分片段三种染色体的小鼠。或者将(实施例70)中所得保持人2号染色体(片段)和人14号染色体(片段)的小鼠品系与(实施例69)中所得保持人22号染色体片段的小鼠品系进行交配,同样可得到同时保持3种人染色体的小鼠。
(实施例73)通过交配制备能主要产生完整人抗体的小鼠品系
将保持人2号+14号(实施例70)、14号+22号(实施例71)、2号+14号+22号(实施例72)染色体的小鼠品系与内源性抗体重链缺陷(实施例67;Kitamura等,自然,350,423-,1991)、轻链κ(Zou等,EMBO.J.12,811-,1993;Chem等,EMBO.J.,12,821-,1993)基因缺陷的小鼠品系进行反复交配,通过PCR等分析(实施例9、30、42、43)筛选保持人2号+14号、14号+22号、或2号+14号+22号染色体的小鼠,建立主要产生完整的人抗体的小鼠系统(Green等,Nature Genetics,7,13-,1994;Lonberg等,自然,368,856-,1994)。
(实施例74)从交配得到的保持含人抗体基因的人染色体的小鼠品系制备产生人抗体的杂交瘤
与(实施例25)相同,用目的抗原免疫(实施例42、43、68、69、70、71、72、73)中得到的、保持有含人抗体基因的人染色体的小鼠,取脾,与骨髓瘤细胞融合制备杂交瘤。用ELISA方法分析1~3周培养期间的培养上清。按(实施例14、15、21、22、25、33、34、37、38)的方法进行ELISA,得到人抗体阳性及人抗体阳性且对免疫抗原特异的克隆。
(实施例75)小鼠抗体重链两侧等位基因均破坏了的TT2F细胞株来源的嵌合体小鼠血清中小鼠IgM的检测及定量。
按照(实施例40)的方法,从(实施例51)中获得的小鼠抗体重链两个等位基因均破坏了的TT2F细胞株(#131-3)所产生的子代小鼠中分别得到嵌合率为0%、50%、99%的3个小鼠,对其血清中小鼠IgM进行检测和定量。从出生后2周龄的嵌合体小鼠采血,用(实施例14)的ELISA方法对血清中小鼠IgM的浓度进行定量。固定PBS稀释的抗小鼠IgM抗体(Kirkegaard&Perry Laboratories Inc.,01-18-03),然后加入血清样品,血清样品用添加了5%FBS的PBS稀释。加入过氧化物酶标记的抗小鼠IgM抗体(Kirkegaard&PerryLaboratories Inc.,074-1803),以TMBZ作底物,测定450nm的吸光度。以纯化的小鼠IgM(Phamningn,03081D)作标准,用补充有FBS的PBS进行梯度稀释。结果如表20所示,该结果显示由小鼠抗体重链两侧等位基因破坏了的TT2F细胞而来的嵌合体小鼠中,嵌合率为99%的小鼠中小鼠IgM浓度低,这表明ES细胞的小鼠重链基因基本上没有功能。
表20嵌合体小鼠中小鼠IgM的浓度(ELISA)
产业上利用的可能性
本发明提供了一种嵌合体非人动物,它保持有单一或多个外源染色体或其片段,并能表达该染色体或其片段上的基因。利用本发明的嵌合体非人动物,可制备生物活性物质。
本发明提供具有分化多能性的细胞,它保持有单一或多个外源染色体或其片段,并能表达该外源染色体或其片段上的基因。利用该细胞,通过骨髓移植等方法可治疗遗传病。
序列表
序列号:1
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TGGAAGGTGG ATAACGCCCT
序列号:2
序列长度:22
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TCATTCTCCT CCAACATTAG CA
序列号:3
序列长度:21
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸 合成DNA
序列
TGTAGGGGAC CTGGAGCCTT G
序列号:4
序列长度:21
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TTGACAACTC ACCTGGACTA G
序列号:5
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CTCTCCTGCA GGGCCAGTCA
序列号:6
序列长度:22
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TGCTGATGGT GAGAGTGAAC TC
序列号:7
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
AGTCAGGGCA TTAGCAGTGC
序列号:8
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GCTGCTGATG GTGAGAGTGA
序列号:9
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TGGTGGCTGA AAGCTAAGAA
序列号:10
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CCAGAAGAAT GGTGTCATTA
序列号:11
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TCCAGGTTCT GCAGAGCAAG
序列号:12
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TGTAGTTGGA GGCCATGTCC
序列号:13
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CCCCACCCAT GATCCAGTAC
序列号:14
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GCCCTCAGAA GACGAAGCAG
序列号:15
序列长度:22
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GAGAGTTGCA GAAGGGGTGA CT
序列号:16
序列长度:22
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸 合成DNA
序列
GGAGACCACC AAACCCTCCA AA
序列号:17
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GGCTATGGGG ACCTGGGCTG
序列号:18
序列长度:22
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CAGAGACACA GGCACGTAGA AG
序列号:19
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TTAAGGGTCA CCCAGAGACT
序列号:20
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TGTAGTTGGA GGCCATGTCC
序列号:21
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸 合成DNA
序列
CAAAAAGTCC AACCCTATCA
序列号:22
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GCCCTCAGAA GACGAAGCAG
序列号:23
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TCGTTCCTGT CGAGGATGAA
序列号:24
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TCACTCCGAA GCTGCCTTTC
序列号:25
序列长度:21
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸 合成DNA
序列
ATGTACAGGA TGCAACTCCT G
序列号:26
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TCATCTGTAA ATCCAGCAGT
序列号:27
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GATCCCATCG CAGCTACCGC
序列号:28
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TTCGCCGAGT AGTCGCACGG
序列号:29
序列长度:22
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GATGAACTAG TCCAGGTGAG TT
序列号:30
序列长度:22
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CCTTTTGGCT TCTACTCCTT CA
序列号:31
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
ATAGAGGGTA CCCACTCTGG
序列号:32
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
AACCAGGTAG GTTGATATGG
序列号:33
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
AAGTTCCTGT GATGTCAAGC
序列号:34
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TCATGAGCAG ATTAAACCCG
序列号:35
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TGTGAAGGAG GACCAGGTGT
序列号:36
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TGTAGGGGTT GACAGTGACA
序列号:37
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CTGAGAGATG CCTCTGGTGC
序列号:38
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GGCGGTTAGT GGGGTCTTCA
序列号:39
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GGTGTCGTGG AACTCAGGCG
序列号:40
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CTGGTGCAGG ACGGTGAGGA
序列号:41
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GCATCCTGAC CGTGTCCGAA
序列号:42
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GGGTCAGTAG CAGGTGCCAG
序列号:43
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
AGTGAGATAA GCAGTGGATG
序列号:44
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GTTGTGCTAC TCCCATCACT
序列号:45
序列长度:21
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TTGTATTTCC AGGAGAAAGT G
序列号:46
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GGAGACGAGG GGGAAAAGGG
序列号:47
序列长度:27
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
ATGGACTGGA CCTGGAGGRT CYTCTKC
序列号:48
序列长度:27
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
ATGGAGYTTG GGCTGASCTG GSTTTYT
序列号:49
序列长度:27
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
ATGRAMMWAC TKTGKWBCWY SCTYCTG
序列号:50
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CAGAGGCAGT TCCAGATTTC
序列号:51
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TGGGATAGAA GTTATTCAGC
序列号:52
序列长度:20
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
ATGGACATGR RRDYCCHVGY KCASCTT
序列号:53
序列长度:28
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CCAAGCTTCA GGAGAAAGTG ATGGAGTC
序列号:54
序列长度:28
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CCAAGCTTAG GCAGCCAACG GCCACGCT
序列号:55
序列长度:28
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CCAAGCTTCA GAGGCAGTTC CAGATTTC
序列号:56
序列长度:28
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GGGAATTCGG GTAGAAGTCA CTGATCAG
序列号:57
序列长度:28
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GGGAATTCGG GTAGAAGTCA CTTATGAG
序列号:58
序列长度:28
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
GGGAATTCGG GTAGAAGTCA CTTACGAG
序列号:59
序列长度:60
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
ACCTTCATCG TCCTCTTCCT CCTGAGCCTC TTCTACAGCA
CCACCGTCAC CCTGTTCAAG
序列号:60
序列长度:60
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
TGATGCTGCA CCAACTGTAT CCATCTTCCC ACCATCCAGT
GAGCAGTTAA CATCTGGAGG
序列号:61
序列长度:22
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CTGGGGTGAG CCGGATGTTT TG
序列号:62
序列长度:22
序列类型:核酸
链数:单链
拓朴学:线性
序列的种类:其它核苷酸合成DNA
序列
CCAACCCAGC TCAGCCCAGT TC
Claims (23)
1.制备能表达人抗体基因的嵌合非人动物或其子代的方法,其包括下述步骤:制备包含至少一个下述的人染色体片段的微细胞,所述的染色体片段选自碱基长度不小于1Mb、包含编码人抗体的基因或其大片段功能序列部分的染色体2,14和22的片段;通过与所述的微细胞融合将所述的染色体片段转化到非人动物多能细胞中,由此制备保有所述的至少一个人染色体片段的多能细胞;由所述的多能细胞制备所述的嵌合非人动物;可选择地,由所述的嵌合非人动物生产其子代。
2.如权利要求1所述的方法,其中,所述的非人动物、其子代或多能细胞保有包含人抗体重链基因的人染色体14的片段。
3.如权利要求1所述的方法,其中,所述的非人动物、其子代或多能细胞保有包含人抗体轻链κ基因的人染色体2的片段。
4.如权利要求1所述的方法,其中,所述的非人动物、其子代或多能细胞保有包含人抗体轻链λ基因的人染色体22的片段。
5.如权利要求1所述的方法,其中,所述的非人动物、其子代或多能细胞保有包含人抗体轻链κ基因的人染色体2的片段和包含人抗体轻链λ基因的人染色体22的片段。
6.如权利要求1所述的方法,其中,所述的非人动物、其子代或多能细胞保有包含人抗体重链基因的人染色体14的片段和包含人抗体轻链κ基因的人染色体2的片段。
7.如权利要求1所述的方法,其中,所述的非人动物、其子代或多能细胞保有包含人抗体重链基因的人染色体14的片段和包含人抗体轻链λ基因的人染色体22的片段。
8.如权利要求1所述的方法,其中,所述的含有至少一个染色体片段的微细胞是通过诱导下述杂合细胞得来的,所述杂合细胞是由衍生所述染色体片段的细胞与具备有效形成微细胞能力的细胞融合形成的。
9.如权利要求8所述的方法,其中,所述的含有至少一个染色体片段的微细胞是通过诱导下述细胞得来的,所述细胞是将诱导上述杂合细胞所得的微细胞与具备有效形成微细胞能力的细胞进一步融合所得的。
10.如权利要求8所述的方法,其中,作为所述至少一个染色体片段的来源的细胞是人正常二倍体细胞。
11.如权利要求8所述的方法,其中,具备有效形成微细胞能力的细胞是小鼠A9细胞。
12.如权利要求1所述的方法,其中,所述的多能细胞是胚胎干细胞。
13.如权利要求1所述的方法,其中,所述的ES细胞来源于小鼠。
14.如权利要求1所述的方法,其中,所述的多能细胞具有经破坏的、与所述染色体片段上的编码人抗体的基因或其大片段功能序列同一或同源的基因。
15.如权利要求1所述的方法,其中,所述的嵌合非人动物细胞抗原能够将所述的至少一个染色体片段传递到其子代中。
16.如权利要求1所述的方法,其中,所述的嵌合非人动物是哺乳动物。
17.如权利要求16所述的方法,其中,所述的哺乳动物是小鼠。
18.制备人抗体的方法,其包括在由权利要求1的方法制备的除小鼠之外的、嵌合非人动物、其子代、其组织或其细胞中表达至少一个染色体片段上的人抗体基因,并收集人抗体。
19.如权利要求18所述的方法,其中,所述的嵌合非人动物或其子代的细胞是B细胞。
20.如权利要求19所述的方法,其中,所述的B细胞通过与骨髓瘤细胞融合无限增殖化。
21.制备多克隆人抗体的方法,其包括用抗原免疫由权利要求1的方法制备的除小鼠之外的、嵌合非人动物或其子代,从所述动物的血液或腹水中收集抗血清。
22.制备单克隆人抗体的方法,其包括用抗原免疫由权利要求1的方法制备的除小鼠之外的、嵌合非人动物或其子代,从来源于所述动物或其子代的杂交瘤收集针对所述抗原的单克隆抗体。
23.制备人抗体的方法,其包括用抗原免疫由权利要求1的方法制备的除小鼠之外的、嵌合非人动物或其子代,从经免疫的动物中提取人抗体基因或其大片段功能序列;将所述的人抗体基因或其大片段功能序列转化到细胞中;培养所述的细胞;收集针对所述抗原的抗体。
Applications Claiming Priority (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP24234095 | 1995-08-29 | ||
| JP242340/95 | 1995-08-29 | ||
| JP2794096 | 1996-02-15 | ||
| JP27940/96 | 1996-02-15 |
Related Parent Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CNB961977876A Division CN100379857C (zh) | 1995-08-29 | 1996-08-29 | 嵌合体动物及其制备方法 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN101333516A true CN101333516A (zh) | 2008-12-31 |
Family
ID=26365944
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CNA2008100099774A Pending CN101333516A (zh) | 1995-08-29 | 1996-08-29 | 嵌合体动物及其制备方法 |
| CNB961977876A Expired - Lifetime CN100379857C (zh) | 1995-08-29 | 1996-08-29 | 嵌合体动物及其制备方法 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CNB961977876A Expired - Lifetime CN100379857C (zh) | 1995-08-29 | 1996-08-29 | 嵌合体动物及其制备方法 |
Country Status (11)
| Country | Link |
|---|---|
| EP (3) | EP0773288A3 (zh) |
| JP (1) | JP4318736B2 (zh) |
| KR (1) | KR100308764B1 (zh) |
| CN (2) | CN101333516A (zh) |
| AT (1) | ATE352613T1 (zh) |
| AU (1) | AU718138B2 (zh) |
| CA (1) | CA2230759C (zh) |
| DE (1) | DE69636868T2 (zh) |
| DK (1) | DK0843961T3 (zh) |
| TW (1) | TW448032B (zh) |
| WO (1) | WO1997007671A1 (zh) |
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2018233608A1 (en) * | 2017-06-19 | 2018-12-27 | Beijing Biocytogen Co., Ltd | Genetically modified non-human animal with human or chimeric cd28 |
| CN109837307A (zh) * | 2017-11-24 | 2019-06-04 | 中国科学院上海生命科学研究院 | 建立含外源染色体的胚胎干细胞的新方法 |
| CN109906272A (zh) * | 2016-10-31 | 2019-06-18 | 国立大学法人鸟取大学 | 产生人抗体的非人动物和使用该非人动物的人抗体制作方法 |
| US11350614B2 (en) | 2017-06-19 | 2022-06-07 | Biocytogen Pharmaceuticals (Beijing) Co., Ltd. | Genetically modified non-human animal with human or chimeric CD28 |
Families Citing this family (170)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB8908127D0 (en) * | 1989-04-11 | 1989-05-24 | Shaw Francis & Co Ltd | Mixer and a method of a mixer control |
| US7084260B1 (en) | 1996-10-10 | 2006-08-01 | Genpharm International, Inc. | High affinity human antibodies and human antibodies against human antigens |
| US5770429A (en) * | 1990-08-29 | 1998-06-23 | Genpharm International, Inc. | Transgenic non-human animals capable of producing heterologous antibodies |
| EP1707051B1 (en) * | 1997-02-28 | 2010-01-13 | Kyowa Hakko Kirin Co., Ltd. | Human chromosome fragment, and vector containing it |
| AU7057398A (en) * | 1997-04-11 | 1998-11-11 | Imperial College Of Science, Technology And Medicine | Chromosome transfer (xmmct) to es cells, induced during the exposure of microcells to radiation |
| TWI255853B (en) * | 1998-08-21 | 2006-06-01 | Kirin Brewery | Method for modifying chromosomes |
| JP4689781B2 (ja) * | 1998-09-03 | 2011-05-25 | 独立行政法人科学技術振興機構 | アミノ酸輸送蛋白及びその遺伝子 |
| CA2375827C (en) | 1999-06-01 | 2017-01-10 | Biogen, Inc. | A blocking monoclonal antibody to vla-1 and its use for the treatment of inflammatory disorders |
| US6833268B1 (en) | 1999-06-10 | 2004-12-21 | Abgenix, Inc. | Transgenic animals for producing specific isotypes of human antibodies via non-cognate switch regions |
| WO2001011951A1 (fr) * | 1999-08-13 | 2001-02-22 | Kirin Beer Kabushiki Kaisha | Souris renfermant un cytochrome p450 humain |
| CA2589418A1 (en) * | 1999-08-24 | 2001-03-01 | Medarex, Inc. | Human ctla-4 antibodies and their uses |
| US7605238B2 (en) | 1999-08-24 | 2009-10-20 | Medarex, Inc. | Human CTLA-4 antibodies and their uses |
| US6835370B2 (en) * | 1999-11-08 | 2004-12-28 | Rhode Island Hospital | Diagnosis and treatment of malignant neoplasms |
| US7820878B2 (en) | 1999-11-19 | 2010-10-26 | Kyowa Hakko Kirin Co., Ltd. | Production of ungulates, preferably bovines that produce human immunoglobulins |
| US7074983B2 (en) | 1999-11-19 | 2006-07-11 | Kirin Beer Kabushiki Kaisha | Transgenic bovine comprising human immunoglobulin loci and producing human immunoglobulin |
| US7414170B2 (en) | 1999-11-19 | 2008-08-19 | Kirin Beer Kabushiki Kaisha | Transgenic bovines capable of human antibody production |
| AU2001236807A1 (en) * | 2000-02-10 | 2001-08-20 | Abbott Laboratories | Antibodies that bind human interleukin-18 and methods of making and using |
| JP2003530113A (ja) * | 2000-04-07 | 2003-10-14 | フラームス・インテルウニフェルシタイル・インステイチュート・フォール・ビオテヒノロヒー・ヴェーゼットウェー | 新規な染色体性ベクターおよびその用途 |
| DE60144305D1 (de) | 2000-11-17 | 2011-05-05 | Kyowa Hakko Kirin Co Ltd | Expression von xenogenen (humanen) Immunglobulinen in klonierten, transgenen Huftieren |
| IL155977A0 (en) | 2000-11-30 | 2003-12-23 | Medarex Inc | Transgenic transchromosomal rodents for making human antibodies |
| WO2002059263A2 (en) * | 2000-12-19 | 2002-08-01 | Sunol Molecular Corporation | Transgenic animals comprising a humanized immune system |
| HUP0402250A3 (en) | 2001-04-13 | 2007-06-28 | Biogen Idec Inc | Antibodies to vla-1 |
| JP2002316944A (ja) * | 2001-04-17 | 2002-10-31 | Chemo Sero Therapeut Res Inst | ヒトポリクローナル抗体組成物 |
| DE60227067D1 (de) * | 2001-05-11 | 2008-07-24 | Kirin Pharma Kk | Künstliches menschliches chromosom mit dem gen für die lambda-leichte kette menschlicher antikörper |
| EP1452093A4 (en) * | 2001-11-15 | 2007-08-15 | Kirin Brewery | CHIMESE NON-MENTAL ANIMAL |
| US7303896B2 (en) | 2002-02-25 | 2007-12-04 | Genentech, Inc. | Nucleic acid encoding novel type-1 cytokine receptor GLM-R |
| CN102659946A (zh) | 2002-03-13 | 2012-09-12 | 拜奥根Idec马萨诸塞公司 | 抗αvβ6抗体 |
| SI1503794T1 (sl) | 2002-04-12 | 2012-09-28 | Medarex Inc | Postopek zdravljenja z uporabo ctla-4 antiteles |
| IL165088A0 (en) * | 2002-05-17 | 2005-12-18 | Kirin Brewery | Transgenic ungulates capable of human antibody production |
| EP2243493A1 (en) | 2002-07-03 | 2010-10-27 | Ono Pharmaceutical Co., Ltd. | Immunopotentiative composition |
| KR101317045B1 (ko) * | 2002-09-06 | 2013-10-16 | 암젠 인코포레이티드 | 치료학적 인체 항-il-1r1 모노클로날 항체 |
| EP1563084A4 (en) | 2002-11-08 | 2006-03-01 | Hematech Llc | TRANSGENE UNGULATES WITH REDUCED PRION PROTEIN ACTIVITY AND USES THEREOF |
| EP1578799B8 (en) | 2002-12-02 | 2011-03-23 | Amgen Fremont Inc. | Antibodies directed to tumor necrosis factor and uses thereof |
| CA2528551A1 (en) | 2003-06-13 | 2005-01-13 | Biogen Idec Ma Inc. | Aglycosyl anti-cd154 (cd40 ligand) antibodies and uses thereof |
| KR101435167B1 (ko) | 2003-06-27 | 2014-11-04 | 암젠 프레몬트 인코포레이티드 | 상피 성장 인자 수용체의 결실 돌연변이체 지향 항체 및 그용도 |
| BR0318454A (pt) | 2003-08-08 | 2006-09-12 | Abgenix Inc | anticorpos dirigidos a hormÈnio da paratireóide (pth) e seus usos |
| EP1533617A1 (en) | 2003-11-19 | 2005-05-25 | RMF Dictagene S.A. | Angiogenesis inhibiting molecules, their selection, production and their use in the treatment and diagnosis of cancer |
| US7642341B2 (en) | 2003-12-18 | 2010-01-05 | Merck Serono S.A. | Angiogenesis inhibiting molecules, their selection, production and their use in the treatment of cancer |
| PT2177537E (pt) | 2004-01-09 | 2011-12-13 | Pfizer | Anticorpos contra madcam |
| AU2005227313A1 (en) | 2004-03-19 | 2005-10-06 | Amgen Inc. | Reducing the risk of human and anti-human antibodies through V gene manipulation |
| CA2563064A1 (en) | 2004-04-22 | 2005-11-10 | Kirin Beer Kabushiki Kaisha | Transgenic animals and uses thereof |
| JP5102028B2 (ja) | 2004-07-26 | 2012-12-19 | バイオジェン・アイデック・エムエイ・インコーポレイテッド | 抗cd154抗体 |
| EP1811832A4 (en) | 2004-10-22 | 2009-12-30 | Revivicor Inc | NATIVES WITH GENETICALLY MODIFIED IMMUNE SYSTEMS |
| US20080026457A1 (en) | 2004-10-22 | 2008-01-31 | Kevin Wells | Ungulates with genetically modified immune systems |
| MX2007005866A (es) | 2004-11-17 | 2007-11-12 | Amgen Inc | Anticuerpos monoclonales totalmente humanos para il-13. |
| TW200635946A (en) | 2004-12-20 | 2006-10-16 | Abgenix Inc | Binding proteins specific for human matriptase |
| NZ556029A (en) | 2004-12-21 | 2010-04-30 | Astrazeneca Ab | Antibodies directed to angiopoietin-2 and uses thereof |
| EP2361933A3 (en) | 2005-01-26 | 2012-05-02 | Amgen Fremont Inc. | Antibodies against interleukin-1 beta |
| MX2007013978A (es) | 2005-05-09 | 2008-02-22 | Ono Pharmaceutical Co | Anticuerpos monoclonales humanos a muerte programada 1 (pd-1) y metodos para tratamiento de cancer utilizando anticuerpos anti-pd-1 solos o en combinacion con otros inmunoterapeuticos. |
| CN105330741B (zh) | 2005-07-01 | 2023-01-31 | E.R.施贵宝&圣斯有限责任公司 | 抗程序性死亡配体1(pd-l1)的人单克隆抗体 |
| CN102875681A (zh) | 2005-07-08 | 2013-01-16 | 拜奥根Idec马萨诸塞公司 | 抗-αvβ6抗体及其用途 |
| CA2619695A1 (en) | 2005-08-18 | 2007-02-22 | Genmab A/S | Therapy with cd4 binding peptides and radiation |
| UA92504C2 (en) | 2005-10-12 | 2010-11-10 | Эли Лилли Энд Компани | Anti-myostatin monoclonal antibody |
| NZ568016A (en) | 2005-12-07 | 2011-12-22 | Medarex Inc | CTLA-4 antibody dosage escalation regimens |
| US7838638B2 (en) | 2005-12-13 | 2010-11-23 | Eli Lilly And Company | Anti-IL-17 antibodies |
| PL1979001T3 (pl) | 2005-12-13 | 2012-09-28 | Medimmune Ltd | Białka wiążące specyficzne dla insulinopodobnych czynników wzrostu i ich zastosowania |
| CA2633211A1 (en) | 2005-12-15 | 2007-06-21 | Astrazeneca Ab | Combination of angiopoietin-2 antagonist and of vegf-a, kdr and/or flt1 antagonist for treating cancer |
| AR056857A1 (es) | 2005-12-30 | 2007-10-24 | U3 Pharma Ag | Anticuerpos dirigidos hacia her-3 (receptor del factor de crecimiento epidérmico humano-3) y sus usos |
| KR20080105111A (ko) * | 2006-02-28 | 2008-12-03 | 온코세라피 사이언스 가부시키가이샤 | 항 EphA4 항체의 효과기 기능을 이용한 세포 손상 방법 |
| TW200813091A (en) | 2006-04-10 | 2008-03-16 | Amgen Fremont Inc | Targeted binding agents directed to uPAR and uses thereof |
| JP5496658B2 (ja) | 2006-05-25 | 2014-05-21 | バイオジェン・アイデック・エムエイ・インコーポレイテッド | 脳卒中を処置する方法 |
| JP5345391B2 (ja) | 2006-07-07 | 2013-11-20 | 協和発酵キリン株式会社 | ヒト人工染色体(hac)ベクター及びヒト人工染色体(hac)ベクターを有するヒト細胞医薬 |
| JP2009542810A (ja) | 2006-07-10 | 2009-12-03 | バイオジェン・アイデック・エムエイ・インコーポレイテッド | Smad4欠損癌の増殖を阻害するための組成物および方法 |
| TW200815474A (en) | 2006-08-03 | 2008-04-01 | Astrazeneca Ab | Antibodies alphaVbeta6 and uses thereof |
| CL2007002225A1 (es) | 2006-08-03 | 2008-04-18 | Astrazeneca Ab | Agente de union especifico para un receptor del factor de crecimiento derivado de plaquetas (pdgfr-alfa); molecula de acido nucleico que lo codifica; vector y celula huesped que la comprenden; conjugado que comprende al agente; y uso del agente de un |
| WO2008038127A2 (en) | 2006-09-28 | 2008-04-03 | Merck Serono S.A. | Junctional adhesion molecule-c (jam-c) binding compounds and methods of their use |
| RU2448979C2 (ru) | 2006-12-14 | 2012-04-27 | Ридженерон Фармасьютикалз, Инк. | Антитела человека к дельта-подобному лиганду-4 человека |
| CA2680549A1 (en) | 2007-03-12 | 2008-09-18 | Alan D. D'andrea | Prognostic, diagnostic, and cancer therapeutic uses of fanci and fanci modulating agents |
| BRPI0809042B1 (pt) | 2007-03-22 | 2021-08-31 | Biogen Ma Inc. | Proteína de ligação a cd154 isolada, seu uso, e composição |
| EP2006304A1 (en) * | 2007-06-22 | 2008-12-24 | Stiftung Für Diagnostische Forschung | Anti-HPA-5b monoclonal antibody |
| EP2185719B1 (en) | 2007-08-02 | 2013-11-13 | NovImmune SA | Anti-rantes antibodies and methods of use thereof |
| EP4248976A3 (en) | 2007-08-23 | 2024-04-10 | Amgen Inc. | Antigen binding proteins to proprotein convertase subtilisin kexin type 9 (pcsk9) |
| JOP20080381B1 (ar) | 2007-08-23 | 2023-03-28 | Amgen Inc | بروتينات مرتبطة بمولدات مضادات تتفاعل مع بروبروتين كونفيرتاز سيتيليزين ككسين من النوع 9 (pcsk9) |
| CA2700723A1 (en) | 2007-09-26 | 2009-04-02 | Amgen Inc. | Heparin-binding epidermal growth factor-like growth factor antigen binding proteins |
| CA2700860C (en) * | 2007-10-01 | 2016-07-19 | Jonathan A. Terrett | Human antibodies that bind mesothelin, and uses thereof |
| GB0720552D0 (en) * | 2007-10-19 | 2007-11-28 | Iti Scotland Ltd | Multiple cluster knockout |
| MY155621A (en) | 2007-11-12 | 2015-11-13 | U3 Pharma Gmbh | Axl antibodies |
| JP5778577B2 (ja) | 2008-09-19 | 2015-09-16 | メディミューン,エルエルシー | Dll4に対する抗体およびその使用 |
| EP2348827B1 (en) | 2008-10-27 | 2015-07-01 | Revivicor, Inc. | Immunocompromised ungulates |
| CA2748158A1 (en) | 2008-12-23 | 2010-07-01 | Astrazeneca Ab | Targeted binding agents directed to .alpha.5.beta.1 and uses thereof |
| SG175276A1 (en) | 2009-05-05 | 2011-11-28 | Novimmune Sa | Anti-il-17f antibodies and methods of use thereof |
| ES2781299T3 (es) | 2009-11-13 | 2020-09-01 | Daiichi Sankyo Europe Gmbh | Materiales y métodos para tratar o prevenir las enfermedades asociadas a HER-3 |
| JPWO2011062206A1 (ja) * | 2009-11-17 | 2013-04-04 | 協和発酵キリン株式会社 | ヒト人工染色体ベクター |
| RS60033B1 (sr) | 2009-11-24 | 2020-04-30 | Medimmune Ltd | Ciljano vezujući agensi usmereni na b7-h1 |
| CA2815154A1 (en) | 2010-08-06 | 2012-02-09 | U3 Pharma Gmbh | Use of her3 binding agents in prostate treatment |
| SG192047A1 (en) | 2011-01-24 | 2013-08-30 | Univ Singapore | Pathogenic mycobacteria-derived mannose-capped lipoarabinomannan antigen binding proteins |
| SG194111A1 (en) | 2011-04-07 | 2013-11-29 | Amgen Inc | Novel egfr binding proteins |
| JOP20200043A1 (ar) | 2011-05-10 | 2017-06-16 | Amgen Inc | طرق معالجة أو منع الاضطرابات المختصة بالكوليسترول |
| CN107903325B (zh) | 2011-05-16 | 2021-10-29 | 埃泰美德(香港)有限公司 | 多特异性fab融合蛋白及其使用方法 |
| AU2012332590B2 (en) | 2011-11-01 | 2016-10-20 | Bionomics, Inc. | Anti-GPR49 antibodies |
| WO2013067060A1 (en) | 2011-11-01 | 2013-05-10 | Bionomics, Inc. | Anti-gpr49 antibodies |
| US10598653B2 (en) | 2011-11-01 | 2020-03-24 | Bionomics Inc. | Methods of blocking cancer stem cell growth |
| AU2012332587B2 (en) | 2011-11-01 | 2017-02-23 | Bionomics, Inc. | Antibodies and methods of treating cancer |
| TWI679212B (zh) | 2011-11-15 | 2019-12-11 | 美商安進股份有限公司 | 針對bcma之e3以及cd3的結合分子 |
| US20140314667A1 (en) | 2011-11-16 | 2014-10-23 | Amgen Inc. | Methods of treating epidermal growth factor deletion mutant viii related disorders |
| RS59159B1 (sr) | 2012-02-06 | 2019-10-31 | Inhibrx Inc | Cd47 antitela i postupci za njihovu upotrebu |
| CA2864539C (en) | 2012-02-16 | 2022-06-07 | Santarus, Inc. | Antibody formulations |
| DK2825037T3 (da) | 2012-03-16 | 2019-07-29 | Regeneron Pharma | Gnavere, der udtrykker pH-sensitive immunoglobulinsekvenser |
| SMT201800229T1 (it) | 2012-03-16 | 2018-07-17 | Regeneron Pharma | Anticorpi a catena leggera ingegnerizzati con istidina e roditori modificati geneticamente per la generazione degli stessi |
| JP6133544B2 (ja) * | 2012-03-16 | 2017-05-24 | 康宏 香月 | ミクロセル保存方法 |
| EA039663B1 (ru) | 2012-05-03 | 2022-02-24 | Амген Инк. | Применение антитела против pcsk9 для снижения сывороточного холестерина лпнп и лечения связанных с холестерином расстройств |
| CN103387955B (zh) * | 2012-05-10 | 2015-04-29 | 上海南方模式生物科技发展有限公司 | 非人哺乳动物b淋巴细胞缺陷动物模型的制备方法及其用途 |
| JO3519B1 (ar) | 2013-01-25 | 2020-07-05 | Amgen Inc | تركيبات أجسام مضادة لأجل cdh19 و cd3 |
| US9920121B2 (en) | 2013-01-25 | 2018-03-20 | Amgen Inc. | Antibodies targeting CDH19 for melanoma |
| PE20151408A1 (es) | 2013-02-06 | 2015-10-15 | Inhibrx Llc | Anticuerpos cd47 que no agotan plaquetas ni globulos rojos y metodos de uso de los mismos |
| WO2014143739A2 (en) | 2013-03-15 | 2014-09-18 | Biogen Idec Ma Inc. | Anti-alpha v beta 6 antibodies and uses thereof |
| WO2014144466A1 (en) | 2013-03-15 | 2014-09-18 | Biogen Idec Ma Inc. | Anti-alpha v beta 6 antibodies and uses thereof |
| EP2970449B1 (en) | 2013-03-15 | 2019-09-25 | Amgen Research (Munich) GmbH | Single chain binding molecules comprising n-terminal abp |
| US9505849B2 (en) | 2013-03-15 | 2016-11-29 | Amgen Research (Munich) Gmbh | Antibody constructs for influenza M2 and CD3 |
| CA2916259C (en) | 2013-06-28 | 2024-02-20 | Amgen Inc. | Methods for treating homozygous familial hypercholesterolemia |
| EP3705494A3 (en) | 2013-08-14 | 2020-12-09 | Sachdev Sidhu | Antibodies against frizzled proteins and methods of use thereof |
| US10570204B2 (en) | 2013-09-26 | 2020-02-25 | The Medical College Of Wisconsin, Inc. | Methods for treating hematologic cancers |
| JOP20200094A1 (ar) | 2014-01-24 | 2017-06-16 | Dana Farber Cancer Inst Inc | جزيئات جسم مضاد لـ pd-1 واستخداماتها |
| JOP20200096A1 (ar) | 2014-01-31 | 2017-06-16 | Children’S Medical Center Corp | جزيئات جسم مضاد لـ tim-3 واستخداماتها |
| DK3138903T3 (da) | 2014-05-02 | 2022-04-19 | Chiome Bioscience Inc | Celler til produktion af humant antistof |
| TWI695011B (zh) | 2014-06-18 | 2020-06-01 | 美商梅爾莎納醫療公司 | 抗her2表位之單株抗體及其使用之方法 |
| TW201609811A (zh) | 2014-07-31 | 2016-03-16 | 安美基研究(慕尼黑)公司 | 具有增強之組織分布之雙特異性單鏈抗體構築體 |
| US11661462B2 (en) | 2014-07-31 | 2023-05-30 | Amgen Research (Munich) Gmbh | Optimized cross-species specific bispecific single chain antibody contructs |
| AR101669A1 (es) | 2014-07-31 | 2017-01-04 | Amgen Res Munich Gmbh | Constructos de anticuerpos para cdh19 y cd3 |
| CN107206071A (zh) | 2014-09-13 | 2017-09-26 | 诺华股份有限公司 | Alk抑制剂的联合疗法 |
| CA2982682A1 (en) | 2015-04-17 | 2016-10-20 | Amgen Research (Munich) Gmbh | Bispecific antibody constructs for cdh3 and cd3 |
| EA039859B1 (ru) | 2015-07-31 | 2022-03-21 | Эмджен Рисерч (Мюник) Гмбх | Биспецифические конструкты антител, связывающие egfrviii и cd3 |
| TWI744242B (zh) | 2015-07-31 | 2021-11-01 | 德商安美基研究(慕尼黑)公司 | Egfrviii及cd3抗體構築體 |
| TW202346349A (zh) | 2015-07-31 | 2023-12-01 | 德商安美基研究(慕尼黑)公司 | Dll3及cd3抗體構築體 |
| TWI796283B (zh) | 2015-07-31 | 2023-03-21 | 德商安美基研究(慕尼黑)公司 | Msln及cd3抗體構築體 |
| TWI717375B (zh) | 2015-07-31 | 2021-02-01 | 德商安美基研究(慕尼黑)公司 | Cd70及cd3抗體構築體 |
| TWI829617B (zh) | 2015-07-31 | 2024-01-21 | 德商安美基研究(慕尼黑)公司 | Flt3及cd3抗體構築體 |
| CN116063544A (zh) | 2016-02-03 | 2023-05-05 | 安进研发(慕尼黑)股份有限公司 | Bcma和cd3双特异性t细胞接合抗体构建体 |
| CA3011942A1 (en) | 2016-02-03 | 2017-08-10 | Amgen Research (Munich) Gmbh | Psma and cd3 bispecific t cell engaging antibody constructs |
| AU2017233121B2 (en) | 2016-03-15 | 2023-12-21 | Itabmed (Hk) Limited | Multispecific Fab fusion proteins and use thereof |
| WO2017160754A1 (en) | 2016-03-15 | 2017-09-21 | Mersana Therapeutics,Inc. | Napi2b-targeted antibody-drug conjugates and methods of use thereof |
| JP2019509322A (ja) | 2016-03-22 | 2019-04-04 | バイオノミクス リミテッド | 抗lgr5モノクローナル抗体の投与 |
| JOP20170091B1 (ar) | 2016-04-19 | 2021-08-17 | Amgen Res Munich Gmbh | إعطاء تركيبة ثنائية النوعية ترتبط بـ cd33 وcd3 للاستخدام في طريقة لعلاج اللوكيميا النخاعية |
| WO2018049261A1 (en) | 2016-09-09 | 2018-03-15 | Icellhealth Consulting Llc | Oncolytic virus expressing immune checkpoint modulators |
| JOP20190189A1 (ar) | 2017-02-02 | 2019-08-01 | Amgen Res Munich Gmbh | تركيبة صيدلانية ذات درجة حموضة منخفضة تتضمن بنيات جسم مضاد يستهدف الخلية t |
| UY37726A (es) | 2017-05-05 | 2018-11-30 | Amgen Inc | Composición farmacéutica que comprende constructos de anticuerpos biespecíficos para almacenamiento y administración mejorados |
| BR112020000698A2 (pt) | 2017-07-14 | 2020-07-14 | Pfizer Inc. | anticorpos contra madcam |
| CN111315780A (zh) | 2017-12-11 | 2020-06-19 | 安进公司 | 双特异性抗体产品的连续制造工艺 |
| UY38041A (es) | 2017-12-29 | 2019-06-28 | Amgen Inc | Construcción de anticuerpo biespecífico dirigida a muc17 y cd3 |
| AU2019313444A1 (en) | 2018-07-30 | 2021-02-18 | Amgen Inc. | Prolonged administration of a bispecific antibody construct binding to CD33 and CD3 |
| AR114541A1 (es) | 2018-08-03 | 2020-09-16 | Amgen Res Munich Gmbh | Constructos de anticuerpos para cldn18.2 y cd3 |
| WO2020043670A1 (en) | 2018-08-27 | 2020-03-05 | Affimed Gmbh | Cryopreserved nk cells preloaded with an antibody construct |
| CN112867735B (zh) | 2018-09-07 | 2025-05-09 | 埃泰美德(香港)有限公司 | 双特异性抗原结合蛋白及其用途 |
| CA3114802A1 (en) | 2018-10-11 | 2020-04-16 | Amgen Inc. | Downstream processing of bispecific antibody constructs |
| AU2020226445B2 (en) | 2019-02-18 | 2025-05-29 | Biocytogen Pharmaceuticals (Beijing) Co., Ltd. | Genetically modified non-human animals with humanized immunoglobulin locus |
| US20220259547A1 (en) | 2019-06-13 | 2022-08-18 | Amgeng Inc. | Automated biomass-based perfusion control in the manufacturing of biologics |
| JP2022542890A (ja) | 2019-07-26 | 2022-10-07 | アムジエン・インコーポレーテツド | 抗il13抗原結合タンパク質 |
| JP7686626B2 (ja) | 2019-09-10 | 2025-06-02 | アムジエン・インコーポレーテツド | 増強されたプロテインl捕捉動的結合容量を有する二重特異性抗原結合ポリペプチドの精製方法 |
| EP4058485A1 (en) | 2019-11-13 | 2022-09-21 | Amgen Inc. | Method for reduced aggregate formation in downstream processing of bispecific antigen-binding molecules |
| MX2022007688A (es) | 2019-12-20 | 2022-07-19 | Amgen Inc | Constructos de anticuerpo multiespecificos agonistas de cd40 dirigidos a mesotelina para el tratamiento de tumores solidos. |
| WO2021130383A1 (en) | 2019-12-27 | 2021-07-01 | Affimed Gmbh | Method for the production of bispecific fcyriii x cd30 antibody construct |
| WO2021150824A1 (en) | 2020-01-22 | 2021-07-29 | Amgen Research (Munich) Gmbh | Combinations of antibody constructs and inhibitors of cytokine release syndrome and uses thereof |
| US20230146593A1 (en) | 2020-03-12 | 2023-05-11 | Amgen Inc. | Method for treatment and prophylaxis of crs in patients comprising a combination of bispecific antibodies binding to cds x cancer cell and tnf alpha or il-6 inhibitor |
| AU2021240028A1 (en) | 2020-03-19 | 2022-09-15 | Amgen Inc. | Antibodies against mucin 17 and uses thereof |
| CA3183756A1 (en) | 2020-05-19 | 2021-11-25 | Amgen Inc. | Mageb2 binding constructs |
| AU2021283564B2 (en) | 2020-06-02 | 2025-05-29 | Biocytogen Pharmaceuticals (Beijing) Co., Ltd. | Genetically modified non-human animals with common light chain immunoglobulin locus |
| CN116368154A (zh) | 2020-10-08 | 2023-06-30 | 阿菲姆德股份有限公司 | 三特异性结合剂 |
| CN116437949A (zh) | 2020-11-06 | 2023-07-14 | 安进公司 | 剪切速率降低的抗原结合结构域 |
| CN116635421A (zh) | 2020-11-06 | 2023-08-22 | 安进公司 | 与cd3结合的多肽构建体 |
| WO2022096700A1 (en) | 2020-11-06 | 2022-05-12 | Amgen Research (Munich) Gmbh | Polypeptide constructs selectively binding to cldn6 and cd3 |
| CR20230229A (es) | 2020-11-06 | 2023-09-05 | Amgen Res Munich Gmbh | Moléculas de unión a antígeno biespecíficas con múltiples dianas de selectividad aumentada |
| CA3215594A1 (en) | 2021-04-02 | 2022-10-06 | Agnieszka KIELCZEWSKA | Mageb2 binding constructs |
| MX2023012931A (es) | 2021-05-06 | 2023-11-13 | Amgen Res Munich Gmbh | Moleculas de union a antigeno dirigidas a cd20 y cd22 para su uso en enfermedades proliferativas. |
| AU2022320948A1 (en) | 2021-07-30 | 2024-01-18 | Affimed Gmbh | Duplexbodies |
| WO2023078968A1 (en) | 2021-11-03 | 2023-05-11 | Affimed Gmbh | Bispecific cd16a binders |
| WO2023079493A1 (en) | 2021-11-03 | 2023-05-11 | Affimed Gmbh | Bispecific cd16a binders |
| EP4389895A4 (en) | 2021-11-16 | 2025-07-30 | Nat Univ Corp Tottori Univ | MAMMALIAN ARTIFICIAL CHROMOSOME VECTOR HAVING A HUMAN IMMUNOGLOBULIN HEAVY CHAIN LOCUS CONTAINING A MODIFIED D REGION, AND CELL OR NON-HUMAN ANIMAL CONTAINING SAID VECTOR |
| TW202346368A (zh) | 2022-05-12 | 2023-12-01 | 德商安美基研究(慕尼黑)公司 | 具有增加的選擇性的多鏈多靶向性雙特異性抗原結合分子 |
| JP2025533434A (ja) | 2022-09-14 | 2025-10-07 | アムジエン・インコーポレーテツド | 二重特異性分子安定化組成物 |
| UY40797A (es) | 2023-06-14 | 2024-12-31 | Amgen Inc | Moléculas captadoras de enmascaramiento de células t |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| ZA874410B (en) * | 1986-06-24 | 1988-04-27 | Merrell Dow Pharma | Fusion products |
-
1996
- 1996-08-29 CN CNA2008100099774A patent/CN101333516A/zh active Pending
- 1996-08-29 DE DE69636868T patent/DE69636868T2/de not_active Expired - Lifetime
- 1996-08-29 EP EP96113839A patent/EP0773288A3/en not_active Withdrawn
- 1996-08-29 AU AU68376/96A patent/AU718138B2/en not_active Expired
- 1996-08-29 EP EP96928694A patent/EP0843961B1/en not_active Expired - Lifetime
- 1996-08-29 WO PCT/JP1996/002427 patent/WO1997007671A1/ja not_active Ceased
- 1996-08-29 AT AT96928694T patent/ATE352613T1/de not_active IP Right Cessation
- 1996-08-29 KR KR1019980701537A patent/KR100308764B1/ko not_active Expired - Lifetime
- 1996-08-29 EP EP07001391A patent/EP1916300A1/en not_active Withdrawn
- 1996-08-29 DK DK96928694T patent/DK0843961T3/da active
- 1996-08-29 CA CA2230759A patent/CA2230759C/en not_active Expired - Lifetime
- 1996-08-29 CN CNB961977876A patent/CN100379857C/zh not_active Expired - Lifetime
- 1996-09-25 TW TW085111701A patent/TW448032B/zh not_active IP Right Cessation
-
2008
- 2008-03-10 JP JP2008060174A patent/JP4318736B2/ja not_active Expired - Lifetime
Cited By (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN109906272A (zh) * | 2016-10-31 | 2019-06-18 | 国立大学法人鸟取大学 | 产生人抗体的非人动物和使用该非人动物的人抗体制作方法 |
| US12063913B2 (en) | 2016-10-31 | 2024-08-20 | National University Corporation Tottori University | Human antibody-producing non-human animal and method for preparing human antibodies using same |
| WO2018233608A1 (en) * | 2017-06-19 | 2018-12-27 | Beijing Biocytogen Co., Ltd | Genetically modified non-human animal with human or chimeric cd28 |
| US11350614B2 (en) | 2017-06-19 | 2022-06-07 | Biocytogen Pharmaceuticals (Beijing) Co., Ltd. | Genetically modified non-human animal with human or chimeric CD28 |
| CN109837307A (zh) * | 2017-11-24 | 2019-06-04 | 中国科学院上海生命科学研究院 | 建立含外源染色体的胚胎干细胞的新方法 |
| CN109837307B (zh) * | 2017-11-24 | 2023-09-19 | 中国科学院脑科学与智能技术卓越创新中心 | 建立含外源染色体的胚胎干细胞的方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| DK0843961T3 (da) | 2007-05-21 |
| EP1916300A1 (en) | 2008-04-30 |
| AU718138B2 (en) | 2000-04-06 |
| CN1200014A (zh) | 1998-11-25 |
| TW448032B (en) | 2001-08-01 |
| AU6837696A (en) | 1997-03-19 |
| DE69636868T2 (de) | 2007-10-25 |
| KR100308764B1 (ko) | 2001-12-17 |
| ATE352613T1 (de) | 2007-02-15 |
| CA2230759C (en) | 2012-02-21 |
| EP0773288A3 (en) | 1997-07-09 |
| KR19990044300A (ko) | 1999-06-25 |
| EP0773288A2 (en) | 1997-05-14 |
| DE69636868D1 (de) | 2007-03-15 |
| HK1008472A1 (zh) | 1999-11-12 |
| EP0843961A1 (en) | 1998-05-27 |
| CN100379857C (zh) | 2008-04-09 |
| EP0843961B1 (en) | 2007-01-24 |
| JP2008200042A (ja) | 2008-09-04 |
| WO1997007671A1 (en) | 1997-03-06 |
| EP0843961A4 (en) | 2000-08-09 |
| CA2230759A1 (en) | 1997-03-06 |
| JP4318736B2 (ja) | 2009-08-26 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN100379857C (zh) | 嵌合体动物及其制备方法 | |
| US6632976B1 (en) | Chimeric mice that are produced by microcell mediated chromosome transfer and that retain a human antibody gene | |
| KR100882030B1 (ko) | 클로닝된 트랜스제닉 유제류에서 이종 (사람)면역글로블린의 발현 | |
| JP4082740B2 (ja) | 内因性遺伝子が破壊されている分化多能性保持細胞 | |
| JP3732407B2 (ja) | 染色体の改変方法 | |
| US7435871B2 (en) | Transgenic animals bearing human Igλ light chain genes | |
| JP5557217B2 (ja) | マウス人工染色体ベクター | |
| JPWO1998037757A1 (ja) | 内因性遺伝子が破壊されている分化多能性保持細胞 | |
| JP3030092B2 (ja) | キメラ動物およびその作製法 | |
| JP2007312792A (ja) | キメラ動物およびその作製法 | |
| JPWO1997007671A1 (ja) | キメラ動物およびその作製法 | |
| JPH11313576A (ja) | キメラ動物およびその作製法 | |
| TWI223585B (en) | Chimeric animal and method for producing the same | |
| HK1008472B (zh) | 嵌合型动物及其构建方法 | |
| HK1012413A (zh) | 嵌合动物及其制备方法 | |
| HK1157135A (zh) | 攜帶人IGλ輕鏈基因的轉基因動物 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| C06 | Publication | ||
| PB01 | Publication | ||
| C10 | Entry into substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| C02 | Deemed withdrawal of patent application after publication (patent law 2001) | ||
| WD01 | Invention patent application deemed withdrawn after publication |
Open date: 20081231 |