Mitose
A mitose ou división do núcleo é o proceso polo cal os cromosomas duplicados se distribúen equitativamente entre as células fillas. É un proceso continuo que, para facilitar o seu estudo, se divide en cinco fases: profase, prometafase, metafase, anafase e telofase.


Profase
editarComeza no momento do ciclo celular no que os cromosomas condensados empezan a ser visíbeis nun xeito de filamentos no interior do núcleo. Ao longo da profase segue a condensación dos cromosomas, que se fan cada vez máis curtos e grosos.
Cada cromosoma aparece constituído por dúas cromátides irmás idénticas, que están unidas polos seus centrómeros; en cada un deles desenvólvese un cinetocoro. Ao mesmo tempo que os cromosomas se condensan, o nucléolo comeza a se descondensar e desaparece progresivamente.
Mentres que no núcleo teñen lugar estes cambios, no citoplasma comeza a se formar o fuso mitótico, que é unha estrutura bipolar composta de microtúbulos. Nas células animais, a formación do fuso está relacionada co centrosoma, que consta de dous pares de centríolos, que se duplicaron durante o período S da interfase. Durante a profase o centrosoma divídese e cada centrosoma fillo diríxese cara a un pólo da célula, organizándose entre eles un feixe de microtúbulos que constitúen o fuso mitótico. Os microtúbulos do fuso denomínanse polares e os exteriores ao fuso, microtúbulos astrais ou áster.
Nas células dos vexetais superiores, que non teñen centriolos, o fuso mitótico fórmase a partir dunha zona difusa, desprovista de orgánulos, situada arredor do núcleo; os microtúbulos aparecen nesa zona, e despois se converte nos pólos do fuso.
Prometafase
editarComeza cando se rompe a envoltura nuclear en pequenas vesículas ou fragmentos de membrana. Ditas vesículas fican durante a mitose nas proximidades do fuso e a partir delas formaranse as novas envolturas nucleares das células fillas.
A rotura da envoltura nuclear permite que os microtúbulos do fuso interaccionen cos cromosomas grazas ao cinetocoro. Os microtúbulos polares capturados polos cinetocoros pasan a se chamar cinetocóricos. En cada cromosoma os microtúbulos cinetocóricos esténdense en direccións opostas dende cada cromátide a un pólo do fuso, e orientan aos cromosomas facendo que se vaian concentrando no plano ecuatorial da célula.
Metafase
editarTodos os cromosomas sitúanse no plano ecuatorial do fuso, constituíndo a placa metafásica. Cada cromosoma mantense nesa posición grazas ás forzas xeradas polos seus microtúbulos cinetocóricos, que están ancorados aos pólos opostos da célula.
É nesta fase cando os cromosomas están máis condensados e por iso son máis visíbeis.
Anafase
editarComeza coa separación simultánea de cada cromosoma nas súas cromátides irmás, que se desprazan cara aos pólos opostos do fuso.
O desprazamento das cromátides prodúcese pola desensamblaxe e acurtamento dos microtúbulos cinetocóricos. Ademais, ao mesmo tempo os dous pólos do fuso mitótico se separan entre si, debido a que os microtúbulos polares se alongan pola ensamblaxe de moléculas de tubulina entre os seus extremos.
Cara ao final da anafase, os cromosomas separáronse en dous grupos iguais, cada un dos cales se atopa nun dos pólos do fuso e fará parte do núcleo dunha nova célula.
Telofase
editar- Artigo principal: Telofase.
Caracterízase pola reconstrución dos núcleos das células fillas. Para isto, fórmase unha nova envoltura nuclear arredor de cada grupo de cromosomas separados. Os cromosomas descondénsanse e os núcleos voltan a aparecer. Unha vez formado o núcleo vólvese á síntese de ARN.
A nova envoltura nuclear de cada célula filla fórmase grazas á fusión das vesículas ou fragmentos de membrana, que se sitúan sobre a superficie dos cromosomas agrupados. A lámina nuclear xoga un papel moi importante neste fenómeno.
Variacións
editarFormas de mitose
editarO proceso da mitose nas células dos organismos eucariotas segue un esquema similar, pero existen variacións en tres importants detalles. Pode distinguirse a mitose "pechada" da "aberta" dependendo de se a envoltura nuclear permanece intacta ou se desfai. Unha forma intermdia con degradación parcial da envoltura nuclear chámase mitose "semiaberta". Con respecto á simetría do aparato do fuso durante a metafase, se o fuso ten unha forma aproximadamente simétrica axialmente (centrada) denomínase "ortomitose", o que se distingue dos fusos excéntricos propios da "pleuromitose", na cal o aparato mitótico ten unha simetría bilateral. Finalmente, un terceiro criterio é a localización do fuso central no caso da pleuromitose pechada, que pode ser: "extranuclear" (fuso localizado no citoplasma) ou "intranuclear" (no núcleo).[1]
-
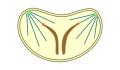 Pleuromitose
Pleuromitose
pechada
intranuclear -
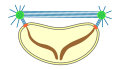 Pleuromitose
Pleuromitose
pechada
extranuclear -
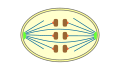 Ortomitose
Ortomitose
pechada -
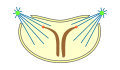 Pleuromitose
Pleuromitose
semiaberta -
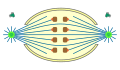 Ortomitose
Ortomitose
semiaberta -
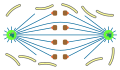 Ortomitose
Ortomitose
aberta






A división nuclear mitótica ten lugar só en células de organismos do dominio eucariota, xa que as bacterias e arqueas non teñen núcleo e divídense doutra maneira. Dentro de cada supergrupo de eucariotas, pode encontrarse a mitose de tipo aberto, pero tamén a mitose pechada, excepto nos Excavata, que presentan exclusivamente mitose pechada.[2] Velaquí as formas de mitose e a súa distribución entre os eucariotas:[1] [3]
- Pleuromitose pechada intranuclear, típica de Foraminifera, algúns Prasinomonadida, algúns Kinetoplastida, os Oxymonadida, os Haplosporidia, moitos fungos (quitridios, oomicetos, cigomicetos, ascomicetos), e algúns Radiolaria (Spumellaria e Acantharia); parece ser o tipo de mitose máis primitivo.
- Pleuromitose pechada extranuclear, que se dá en Trichomonadida e Dinoflagellata.
- Ortomitose pechada, que se encontra en diatomeas, ciliados, algúns microsporidios, lévedos unicelulares e algúns fungos multicelulares.
- Pleurmitose semiaberta, típica da maioría dos Apicomplexa.
- Ortomitose semiaberta, presente con diferentes variantes nalgunhas amebas (Lobosa) e nalgúns flaxelados (por exemplo en Rafidófitas ou Volvox).
- Ortomitose aberta, característica de mamíferos e outros metazoos e das plantas terrestres; pero tamén aparece nalgúns protistas.
Erros e outras variacións
editar
Durante a mitose poden ocorrer erros, especialmente durante o desenvolvemento embrionario inicial humano.[4] Durante cada etapa da mitose, hai normalmente puntos de control que controlan o resultado normal da mitose.[5] Pero de cando en vez poden producirse erros. Os erros mitóticos poden crear células aneuploides que teñen menos ou máis cromosomas do normal, o cal afecta a un par cromosómico ou a varios, unha condición asociada co cancro.[6] [7] As células dos embrións humanos temperáns, as células cancerosas e as células infectadas ou intoxicadas poden tamén sufrir unha división patolóxica dando tres ou máis células fillas (mitose tripolar ou multipolar), o que ten como resultado graves erros nos seus complementos cromosómicos normais.[8]
Na non disxunción, as cromátides irmás non conseguen separarse durante a anafase.[9] Unha célula filla recibe ambas as cromátides irmás do cromosoma non disxunto e a outra célula non recibe ningunha. Como resultado, a primeira célula vai ter tres copias do cromosoma, unha condición chamada trisomía, e a segunda célula terá só unha copia, unha condición chamada monosomía. En ocasións, cando as células experimentan non disxunción, non conseguen completar a citocinese e reteñen ambos os núcleos na mesma célula, resultando células binucleadas.[10]
Retardo da anafase (anaphase lag). Ocorre cando está impedido o movemento dunha cromátide durante a anafase.[9] Isto pode ser causado por un fallo no fuso mitótico á hora de adherirse correctamente ao cromosoma. A cromátida afectada é excluída de ambos os núcleos e pérdese. Por tanto, unha das células fillas será monosómica para ese cromosoma.
A endorreplicación ocore cando os cromosomas se duplican pero a célula non se divide seguidamente. Isto orixina células poliploides ou, se o cromosoma se duplica repetidamente, cromosomas politénicos.[9][11] A endorreplicación atópase en moitas especies e parece ser unha parte normal do seu desenvolvemento.[11] A endomitose é unha variante de endorreplicación na cal as células replican os seus cromosomas durante a fase S e entran na mitose, pero termínana prematuramente. En vez de repartírense nos dous novos núcleos fillos, os cromosomas replicados son retidos dento do núcleo orixinal.[12][13] Entón, as células volven entrar nas fases G1 e S e replican os seus cromosomas de novo.[13] Isto pode acontecer moitas veces, incrementando o número de cromosomas en cada rolda de replicación e endomitose. Os megacariocitos produtores de plaquetas experimentan endomitose durante a diferenciación celular.[14][15]
A amitose en ciliados e en tecidos placentarios animais dá lugar a unha distribución aleatoria dos alelos parentais.
A cariocinese sen citocinese orixina células multinucleadas chamadas cenocitos.
Notas
editar- ↑ 1,0 1,1 Raikov IB (1994). "The diversity of forms of mitosis in protozoa: A comparative review". European Journal of Protistology 30 (3): 253–69. doi:10.1016/S0932-4739(11)80072-6.
- ↑ Boettcher B, Barral Y (2013). "The cell biology of open and closed mitosis". Nucleus 4 (3): 160–5. PMC 3720745. PMID 23644379. doi:10.4161/nucl.24676.
- ↑ R. Desalle, B. Schierwater: Key Transitions in Animal Evolution. CRC Press, 2010, p. 12, link Arquivado 2019-01-02 en Wayback Machine..
- ↑ Mantikou E, Wong KM, Repping S, Mastenbroek S (decembro de 2012). "Molecular origin of mitotic aneuploidies in preimplantation embryos". Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 1822 (12): 1921–30. PMID 22771499. doi:10.1016/j.bbadis.2012.06.013.
- ↑ Wassmann, Katja; Benezra, Robert (2001-02-01). "Mitotic checkpoints: from yeast to cancer". Current Opinion in Genetics & Development (en inglés) 11 (1): 83–90. ISSN 0959-437X. PMID 11163156. doi:10.1016/S0959-437X(00)00161-1.
- ↑ Draviam VM, Xie S, Sorger PK (abril de 2004). "Chromosome segregation and genomic stability". Current Opinion in Genetics & Development 14 (2): 120–5. PMID 15196457. doi:10.1016/j.gde.2004.02.007.
- ↑ Santaguida S, Amon A (agosto de 2015). "Short- and long-term effects of chromosome mis-segregation and aneuploidy". Nature Reviews. Molecular Cell Biology 16 (8): 473–85. PMID 26204159. doi:10.1038/nrm4025. hdl:1721.1/117201.
- ↑ Kalatova B, Jesenska R, Hlinka D, Dudas M (xeneiro de 2015). "Tripolar mitosis in human cells and embryos: occurrence, pathophysiology and medical implications". Acta Histochemica 117 (1): 111–25. PMID 25554607. doi:10.1016/j.acthis.2014.11.009.
- ↑ 9,0 9,1 9,2 Iourov IY, Vorsanova SG, Yurov YB (2006). "Chromosomal Variations in Mammalian Neuronal Cells: Known Facts and Attractive Hypotheses". En Jeon KJ. International Review Of Cytology: A Survey of Cell Biology 249. Waltham, MA: Academic Press. p. 146. ISBN 9780080463506.
- ↑ Shi Q, King RW (outubro de 2005). "Chromosome nondisjunction yields tetraploid rather than aneuploid cells in human cell lines". Nature 437 (7061): 1038–42. Bibcode:2005Natur.437.1038S. PMID 16222248. doi:10.1038/nature03958.
- ↑ 11,0 11,1 Edgar BA, Orr-Weaver TL (maio de 2001). "Endoreplication cell cycles: more for less". Cell 105 (3): 297–306. PMID 11348589. doi:10.1016/S0092-8674(01)00334-8.
- ↑ Lilly MA, Duronio RJ (abril de 2005). "New insights into cell cycle control from the Drosophila endocycle". Oncogene 24 (17): 2765–75. PMID 15838513. doi:10.1038/sj.onc.1208610.
- ↑ 13,0 13,1 Lee HO, Davidson JM, Duronio RJ (novembro de 2009). "Endoreplication: polyploidy with purpose". Genes & Development 23 (21): 2461–77. PMC 2779750. PMID 19884253. doi:10.1101/gad.1829209.
- ↑ Italiano JE, Shivdasani RA (xuño de 2003). "Megakaryocytes and beyond: the birth of platelets". Journal of Thrombosis and Haemostasis 1 (6): 1174–82. PMID 12871316. doi:10.1046/j.1538-7836.2003.00290.x.
- ↑ Vitrat N, Cohen-Solal K, Pique C, Le Couedic JP, Norol F, Larsen AK, Katz A, Vainchenker W, Debili N (maio de 1998). "Endomitosis of human megakaryocytes are due to abortive mitosis". Blood 91 (10): 3711–23. PMID 9573008. doi:10.1182/blood.V91.10.3711.