

List
- Za istoimenu vrstu riba, pogledajte članak List (riba)



- Vrh
- Srednja (primarna) vena (nerv)
- Sekundarna vena
- Lamina
- Rub
- Peteljka
- Pupoljak
- Stabljika
List je veoma značajan organ fotosinteze svih biljnih vrsta među koje spada i drveće. Na njemu su glavni dijelovi: liska, lisna drška (peteljka) i zalisci. Zališće je kod nekih vrsta prilično trajno, dok kod većine vrsta opada još pri razvijanju pupoljaka.
Liska je glavni dio lista. Lisna drška ima važnu ulogu u funkciji lista. Lišče našeg drveća većinom ima peteljku. Ovim je izbjegnuta ukočenost krošnje, a koja bi bila štetna mehanički i fiziološki. Pokretljivošću lišća izbjegava se prejaki udar vjetra i vode. Uz to svaki pokret lista omogućava ulazak svjetlosti dublje u krošnju. Dužina lisne drške je promjenjiva i na istom izbojku, a pogotovo kod različitih vrsta. Nakon opadanja lista na grančici, ispod bočnog pupoljka, ostaje jasno ograničena mala površina, na kojoj je bila učvršćena lisna drška. Ona je određenog oblika, boje i udubljenja, što jasno ukazuje na vrstu drveta sa koje je lišće palo.
Položaj listova na izbojku – grančici je raznolik i karakterističan je za pojedine vrste, a može biti jedan, dva ili više.
List je dorziventuralno spljošten organ, koji obično raste iznad zemlje i specijaliziran je za fotosintezu. U većini listova primarno fotosintetsko tkivo, palisadni mezofil, nalazi se na gornjoj strani lista ili lamine (plojke), ali kod nekih vrsta, uključujući zrelo lišće roda Eucalyptus, palisadni mezofil prisutan je s obje strane, a za lišće se kaže da je izobilateralno. Većina listova ima izraženu gornju površinu (adaksijalnu ) i donju površinu (abaksijalnu) koje se razlikuju u boji, dlakavosti i broju stoma (pore koje unose i ispuštaju plinove), količinu i strukturu epikutikulskog voska i druge karakteristike. Listovi su uglavnom zelene boje. Ovo je zbog Ovo je zbog prisustvo sastojka zvanog hlorofil. On je neophodan za fotosintezu jer apsorbuje svjetlosnu energiju iz sunca. List s bijelim mrljama ili rubovima naziva se varijegatni list.
Listovi mogu imati mnogo različitih oblika, veličina i tekstura. Široki, ravni listovi sa složenim venacijom cvjetnica poznati su kao megafili , a vrste koje ih nose, većina, kao širokolistne ili megafilne biljke. U “klupskim mahovinama, različitog evolucijskog porijekla, listovi su jednostavni (sa samo jednom venom) i poznati su kao mikrofili. Neki listovi, poput krlušti lukovica, nisu iznad zemlje. U mnogih vodenih vrsta listovi su potopljeni u vodu. Sukulentne biljke često imaju debele sočne listove, ali neki listovi su bez većih fotosintetskih funkcija i mogu biti mrtvi u zrelosti, kao u nekim katafiliima i trnju, bodljama. Nadalje, nekoliko vrsta lisnatih struktura koje se nalaze u vaskularnim biljkama nisu s njima potpuno homologni. Primjeri uključuju spljoštene stabljike biljaka nazvane filokladi i kladode, i spljoštene stabljike listova nazvane filode koje se razlikuju od lists po strukturi i porijeklu. Neke strukture nevaskularnih biljaka izgledaju i djeluju poput lišća. Primjeri uključuju filide mahovins i jetrenjača.
Opća svojstva
[uredi | uredi izvor]Listovi su najvažniji organi većine vaskularnih biljaka. Zeleno bilje je autotrofno, što znači da ne dobijaju hranu od drugih živih bića, već umjesto toga stvaraju vlastitu hranu pomoću fotosinteze. Energiju hvataju u sunčevoj svjetlosti i koriste je za sintezu jednostavnih šećera, poput glukoze i saharoze, iz ugljik-dioksida i vode. Šećeri se zatim skladište kao skrob, koji se dalje hemijskom sintezom prerađuju u složenije organske molekule, kao što su proteini ili celuloza, osnovni strukturni materijali u zidovima biljnih ćelija ili se metaboliziraju pomoću ćelijskog disanj da bi se dobila hemijska energija za pokretanje ćelijskih procesa. Listovi izvlače vodu iz zemlje u struji transpiracije kroz vaskularni sistem za provođenje, poznat kao ksilem i dobijaju ugljik dioksid iz atmosphere difuzijom kroz otvore nazvane stome u spoljnjem prekrivajućem sloju lista (epiderme)], dok su listovi orijentirani tako da maksimalno izloženi prema sunčevoj svjetlosti.[1] Jednom kada je šećer sintetiziran, treba ga transportirati u područja aktivnog rasta poput izdanaka biljke i korijena. Vaskularne biljke prenose saharozu u posebnom tkivu zvanom floem. Floem i ksilem međusobno su paralelni, ali transport materija obično je u suprotnim smjerovima. Unutar lista ti se vaskularni sistemi granaju (razgranavaju) da bi formirali vene koje opskrbljuju što je moguće više lista, osiguravajući da ćelije koje obavljaju fotosintezu budu blizu transportnog sistema.[2]
Obično su listovi široki, ravni i tanki (dorziveralno spljošteni), čime se maksimizira površina koja je izravno izložena svjetlosti i omogućava svjetlu da prodre u tkiva i dospije do hloroplasta, čime se promovira fotosinteza. Oni su raspoređeni na biljci tako da svoje površine izlažu svjetlu što je moguće efikasnije, a da se pritom ne zasijenjuju; postoje, međutim, mnogi izuzeci i komplikacije. Naprimjer, biljke prilagođene vjetrovitim uvjetima mogu imati dodatni list, kao što je to slučaj u mnogim vrbama i eukaliptusima. Ravan ili laminarni oblik također maksimizira toplinski kontakt s okolnim zrakom, potičući hlađenje. Funkcionalno, osim što provodi fotosintezu, list je glavno mjesto transpiracije, pružajući energiju potrebnu za izvlačenje transpiracijske struje iz korijena i gutacijom.
Mnoge gimnosperme imaju tanke igličaste ili slične listove koji mogu biti povoljni u hladnom podneblju sa čestim snijegom i mrazom.[3] Ovo se tumači kao smanjenje od megafilnih listića njihovih devonskih predaka[4]Neki oblici listova prilagođeni su da moduliraju količinu svjetlosti koju apsorbuju, kako bi izbjegli ili ublažili prekomjernu toplinu, ultraljubičastog oštećenja ili isušivanje ili žrtvovali efikasnost apsorpcije svjetlosti za pogodnost zaštite od biljojeda. Za kserofite glavno ograničenje nije svjetlost fluks ili intenzitet, već suša. .[5] Neke biljke prozora, kao što su vrste roda Fenestraria i neke roda Haworthia poput Haworthia tesselata i Haworthia truncata tipski primjeri kserofita.[6] a Bulbine mezembriantemoida.
Listovi također funkcioniraju za skladištenje hemijske energije i vode (posebno u sokovima i mogu postati specijalizirani organi koji služe drugim funkcijama, poput vitica graška i drugih mahunarki, zaštitnih bodlji kaktusa i zamke za insekte u mesožderkim biljkama poput rodova Nepenthes i Sarracenia. Listovi su temeljne strukturne jedinice od kojih su zgrađene šišarke u golosjemenjača (svaka skala konusa je modificirani list megafil, poznat kao sporofil)[4]:408 i od kojih su izragrađeni cvjetovi u cvjetnica.[4]:445

Unutrašnja organizacija većine vrsta lišća razvila se tako da maksimizira izlaganje fotosintetskih organela, hloroplasta, svjetlosti i da poveća apsorpciju ugljik-dioksida, dok se istovremeno kontrolira gubitka vode. Njihove površine su vodootporne, sa biljnom kutikulom, a izmjena plinova između ćelija mezofila i atmosfera se kontrolira minutnim (dužinom i širinom izmjerenim u desetinama µm) otvorima zvanim stome, koje se otvaraju ili zatvaraju za regulaciju izmjena brzine ugljik-dioksida, kisika i vodene pare u unutrašnji i međućelijski sistem Stomini otvori kontroliraju turgorov pritisak u paru ćelija zaštitnica koje okružuju stomin otvor. Na bilo kojem kvadratnom centimetru biljnog lista može biti od 1.000 do 100.000 stoma.[7]

Oblik i struktura lišća znatno se razlikuju od vrste do vrste biljaka, ovisno o njihovoj prilagodbi klimi i dostupnoj svjetlosti, ali i drugim faktorima poput ispaše životinja (poput jelena), raspoloživim hranjivim tvarima i ekološkoj konkurenciji drugih biljaka. Značajne promjene u obliku lišća događaju se i u vrstama, naprimjer kako biljka sazrijeva; kao što je slučaj vrsta roda „Eukaliptus“ koje obično imaju izobilateralne, viseće listove kada sazrijevaju i dominiraju nad svojim susjedima. Međutim, takva stabla kao mladice imaju uspravno ili vodoravno dorziventralno lišće, kada je njihov rast ograničen dostupnom svetlošću.[8] Ostali faktori uključuju potrebu za uravnoteženjem gubitka vode pri visokoj temperaturi i niskoj vlažnosti zraka, suprotno potrebi apsorpcije atmosferskog ugljik-dioksida. U većini biljaka listovi su također primarni organi odgovorni za transpiraciju i gutaciju (izlučivanje kuglica tečnosti koje se formiraju na rubu listova).
Listovi takođe mogu skladištiti hranu i vodu, te su prema tome modificirani kako bi ispunili ove funkcije; naprimjer u lišću sočnih biljaka i u ljuskama lukovica. Koncentracija fotosintetskih struktura u lišću zahtijeva da su bogatija proteinma, mineralima i šećerima od, recimo, drvenastih matičnih tkiva. Prema tome, lišće je prominentno u ishrani mnogih životinja.

U skladu s tim, lišće predstavlja velika ulaganja za biljke koje ih imaju, a njihovo zadržavanje ili zbrinjavanje predmet su razrađenih strategija za suzbijanje pritiska štetočina, sezonskih uvjeta i zaštitnih mjera poput rasta trnova, proizvodnje fitolita, lignina, tanin i otrova.
Listopadne biljke u hladnim ili umjereno hladnim regijama obično odbcuju lišće u jesen, dok u područjima koja imaju ozbiljnu sušnu sezonu, neke biljke ne mogu zgubitii lišće sve dok se sušna sezona ne završi. U oba slučaja može se očekivati da listovi, zadržanim hranjivim tvarima, doprinose tlu gdje padaju. Suprotno tome, mnoge druge nesezonske biljke, poput palmi i četinjara, zadržavaju listove dugo vremena; Welwitschia zadržava dva glavna lista tokom cijelog životnog vijeka, koji može prelaziti i hiljadu godina.
Listoliki organi kod briofita (npr. mahovina i Marchantiophyta, poznati kao filidi, morfološki se razlikuju od listovai vaskularnih biljaka, jer kod njih nedostaje vaskularno tkivo; obično su samo jednoćelijske debljine i nemaju kutikule stome ili unutrašnji sistem međućelijskih prostora.
Jednostavni, vaskularizirani listovi (mikrofilidi) prvo su se razvili kao enacije (ekstenzije stabljike), kod klupskih mahovina, poput roda Baragwanathia, tokom silura. Pravi listovi ili eufili veće veličine i složenije nervature nisu postali česti u drugim skupinama sve do devonskog razdoblja, do kada je koncentracija ugljik-dioksida u atmosferi značajno pala. To se dogodilo neovisno u nekoliko zasebnih linija vaskularnih biljaka, u progimnospermi poput roda Archaeopteris, u Sphenopsida, paprati, a kasnije u gimnospermama i angiospermama. Eufili se nazivaju i makrofilima ili megafilima (veliki listovi).[4]
Morfologija
[uredi | uredi izvor]Strukturno potpuni list angiospermi sastoji se od peteljke (stabljika lista), lamine ili liske (lisnasta lopatica), i stipule (male strukture smještene na obje strane dna peteljke). Ne formira se svaka vrsta listova sa svim tim strukturnim komponentama. Stipule mogu biti upadljive (npr. grah i ruža); uskoro opadaju ili na drugi način nisu očigledne kao u’’ Moraceae’’ ili su uopšte odsutne kao u Magnoliaceae. Peteljka može biti odsutna ili liska ne može biti laminarna (spljoštena). Ogromna raznolikost prikazana u obliku lišća (anatomije) od vrste do vrste detaljno je prikazana u nastavku. Peteljka mehanički povezuje list sa osttkom biljke i daje put za prenošenje vode i šećera u list i iz njega. Lamina je obično mjesto glavnine fotosinteze. Gornji adaksijalni ugao između lista i stabljike poznat je pod nazivom aksil’’ lista. Često je lokacija pupoljka. Strukture koje se tamo nalaze nazivaju se "aksilarne" (pazušne).

Vanjske karakteristike lišća, poput oblika, ruba, dlaka, peteljki i prisutnosti pršljenova i žlijezda često su važne za identificiranje biljaka prema razini porodice, roda ili vrsta, a botaničari su razvili bogatu terminologiju za opisivanje karakteristika lista. Listovi gotovo uvijek imaju određeni rast. Narastu do određenog obrasca i oblika, a zatim stanu. Ostali dijelovi biljke poput stabljike ili korijena imaju neodređeni rast i obično će nastaviti rasti sve dok za to imaju resurse.
Svojstva lista su obično karakteristična za vrstu (monomorfna), mada neke vrste proizvode više vrsta lista (dimorfne ili polimorfne]). Najduži listovi su listovi Raffia palma, R. regalis koji mogu biti dugi i do|25 m, a široki I do 3 m.[10] Terminologija povezana s opisom morfologije lista predstavljena je, u ilustriranom obliku, na Wikiknjigama Wikibooks.

Osnovni tipovi lista
[uredi | uredi izvor]
- Paprati imaju frondne listove;
- Lišće četinara obično je igličasto ili u obliku šilice ili u obliku razmjernika;
- Listovi angiospermi (cvjetnice): standardni oblik uključuje lisku, peteljku, laminu:
- Lycophytes ima listove zvane mikrofili.
- Monokotiledone imaju posebne listove koje se nalaei u većini travs i mnogim drugim monokotiledonama:
- Ostali specijalizirani listovi, poput onih iz roda Nepenthes ("biljni vrč")
Raspored na stablu
[uredi | uredi izvor]Za opis rasporeda lišća na stabljici obično se upotrebljavaju različiti izrazi (filotaksis):

- Alternativno
- Jedan list, grana ili cvjetni dio pričvršćuje se u svakoj tački ili čvoru na stabljici i ima, u većem ili manjem stupnju, alternativni smjer duž stabljike.
- Bazni
- Nastaju iz baze stabljike.
- Kaulini
- Nastaju iz zračne stabljike.
- Nasuprotni
- dva lista, grane ili dijelovi cvijeta pričvršćuju se u svakoj tački ili čvoru na stabljici. Prilozi listova su upareni na svakom čvoru i dekusatni ako se, što je tipski, svaki uzastopni par rotira za 90°, napredujući duž stabljike.
- Vrtložni ili verticilatni
- tri ili više listova, grana ili cvjetnih dijelova pričvršćeni su u svakoj tački ili čvoru na stabljici. Kao i kod suprotnih listova, uzastopne vijuge mogu ali ne moraju biti odsječene, zakrenute za pola ugla između listovs u vrtlogu (tj. uzastopne vijuge od tri rotacije za 60°, vijuga od četiri rotacije za 45° itd.). Nasuprotni listovi mogu se pojaviti nakrivljeni blizu vrha stabljike. Pseudoverticillatnost’’ opisuje aranžman koji samo izgleda zgužvano, ali zapravo nije tako.
- Rozulat
- Listovi formiraju rozete.
- Redovi
- Termin "distihozan", doslovno znači "dva reda". Lišće u ovom rasporedu može biti naizmjenično ili suprotno u svom pričvršćivanju. Termin “2-rangirani", je ekvivalentan. Ponekad se susreću i pojmovi „tristihozan“ i „tetrastihozan“. Naprimjer, "listovi" (zapravo mikrofili) većine vrsta roda Selaginella tetrastihozni su, ali nisu dekusatni.
Kako „stabljika“ raste, lišće izgleda raspoređeno oko nje na način koji optimizira iskorištenje svjetlosti. U osnovi, lišće tvori heliks obrazac, usredotočen oko stabljike ili u smjeru kazaljke na satu ili obrnuto ili u smjeru suprotnom od kazaljke na satu, s istim uglom odstupanja (ovisno o vrsti). Postoji pravilnost u tim uglovima i oni slijede brojeve u Fibonaccijevog niza: 1/2, 2/3, 3/5, 5/8, 8/13, 13/21, 21/34, 34/55, 55/89. Ova serija teži zlatnom uglu, što je otprilike 360° × 34/89 ≈ 137.52° ≈ 137° 30′. U seriji, brojnik označava broj potpunih zavoja ili "đira" dok list ne dođe u početni položaj, a nazivnik označava broj listova u rasporedu. To se može pokazati sljedećim:
- Naizmjenični listovi imaju ugao od 180° (ili 1/2)
- 120° (ili 1/3): 3 lista u jednom krugu;
- 144° (ili 2/5): 5 listova u 2 kruga
- 135° (ili 3/8): 8 listova u 3 đira.
Oblici liske (plojke)
[uredi | uredi izvor]
Prema obrascu podijeljenosi liske (lamine, plojke) mogu se opisati dva osnovna oblika listova.
- Jednostavni list ima nepodijeljenu lisku. Međutim, list može biti seciran tako da formira režnjeve, ali praznine između njih ne dosežu do glavne nervature.
- Složeni list ima potpuno podeljenu lisku, a svaki listić" plojke (liske) je odvojen duž glavnog ili sekundarnog nerva. Listovi mogu imati drškice i štikle, ekvivalente peteljki i listiće listova. Kako se svaki list može činiti jednostavnim listom, važno je prepoznati gdje se nalazi peteljka, kako bi se identificirao složeni list. Složeni listovi su karakteristični za neke porodice viših biljaka, poput ‘’Fabaceae’’. Srednja vena složenih listova ili frond, kada postoji, naziva se rahis.
- Prstasto složeni listovi imaju listiće radijalno, od kraja drškice, kao prste na dlanu šake; naprimjer, Cannabis (konoplja) i Aesculus (kesten).
- Perasto složeni
- listovi sa listićima koji se nalaze pored glavne ili srednje vene. Sa terminalnom listićem; naprimjer, Fraxinus (jasen).
- Biperasto složeni
- Listovi su dvostruko podijeljeni: listići su raspoređeni duž sekundarne vene koja je jedan od nekoliko ogranka koji se odvajaju od rašnica. Svaki se listić naziva pinula (perce). Skupina pinula na svakoj sekundarnoj veni formira „pinu“; naprimjer, Albizia (svileno drvo).
- Trolisni (ili trifolioliatni, trifolatni)
- sa listićima u obliku pera; naprimjer, Trifolium (djetelina), Laburnum (laburnum) i neke vrste roda Toxicodendron '(naprimjer, Toksicodendron radicans).
- Pinatifidni
- Perasto seciran na središnjem nervu, ali listići nisu potpuno odvojeni; naprimjer, rod Polypodium, neki predstavnici roda Sorbus (bijeli snopovi). U peasto žilastim listovima središnji nerv poznat jeu kao "srednji vijenac" (midrib).
Svojstva petiole (peteljkice)
[uredi | uredi izvor]
Petiolirani listići imaju petiole (lisne drške), a za njih se kaže da su petiolatne. Sesilni ili sjedeći (epetiolatni) listovi nemaju peteljke i liska se pričvršćuje direktno na stabljiku. Subpetiolatni listovi su gotovo petiolatni ili imaju izrazito kratku peteljku i mogu izgledati kao da sjede.
U listovima stezanja ili dekurentnih liski, one djelomično okružuje stabljiku.
Kad baza lišća potpuno okruži stabljiku, za listove se kaže da su perfoliatni, kao što je primjerice u Eupatorium perfoliatum.
U štitastim (peltatnim) listovima, peteljke se vežu za lisku unutar njene margine.
Kod nekih vrsta roda Acacia, kao što je drvo koa (Acacia koa), peteljke se proširuju ili šire i funkcioniraju poput lisnih listića; nazivaju se filode. Na vrhu filode mogu postojati normalni perasti listovi.
Sporedni listićI ili stipule, prisutni su na listovima mnogih dikotiledona; oni su dodatak na svakoj strani na dnu peteljke, nalik malim listovima. Stipule mogu biti trajne i ne mogu se skinuti (narezani listovi, kao što su kod ruža) i graha, ili otpadaju dok se list širi, ostavljajući izražen ožiljak na grančici (list izdvajanja ). Stanje, uređenje i struktura stipula naziva se "stipulacija".
- Slobodni, bočni (lateralni)
- naprimjer Hibiscus.
- Prirasli
- Spojeni na bazi peteljk, kao što je u rodu Rosa.
- Ohreatni
- opskrbljena sa ohreom ili oblogama u obliku omotača, kao u porodici Polygonaceae; npr. rabarbara.
- Opkoljavajuća baze peteljki
- Između peteljki dva suprotna lista, kao u porodici ‘’Rubiaceae ili Između peteljke i stabljike, kao u Malpighiaceae.
Nervatura ili venacija
[uredi | uredi izvor]


Vene ili nervi čine jednu od vidljivijih osobina listova. Predstavljaju vaskularnu strukturu organa, koja se proteže u listu preko petiola) i omogućavaju transport vode i hranjivih sastojaka između lista i stabljike i imaju presudnu ulogu u održavanju stanje vode listova i fotosintetskog kapaciteta. .[11][12] Oni također igraju ulogu u mehaničkom učvrćivanju lista. U okviru lamine lista, dok neke vaskularne biljke posjeduju samo jednu venu, u većini se ta vaskulatura uglavnom dijeli (razgranava) prema različitim obrascima (venacija) i oblikuje cilindrične snopove, obično ležeći u srednjoj ravnini mezofila, između dva sloja epiderme. Ovaj obrazac je često specifičan za taksone, a od kojih kod angiospermi postoje dva glavna obrasca paralelni i mrežasti ( retickulatni)]. Općenito, paralelna venacija je tipska za monokotiledone, dok je mrežasta više tipska za eudikotiledonee i magnolidne ("dikotiledone"), iako postoje i mnogi izuzeci.
Vena ili vene koje uđu u list iz peteljke nazivaju se primarne ili vene prvog reda. Vene koje se granaju od ovih su vene sekundarnog ili drugog reda. Te se primarne i sekundarne vene smatraju glavnim venama ili venama nižeg reda, mada neki autori uključuju I treći red. Svako naredno grananje se redno broji, a to su vene višeg reda, a svaka grana je povezana s užim promjerom vena. [13] U paralelnim venastim listovima, primarne vene teku paralelno i podjednako jednake drugoj većini dužine lista, a zatim se konvergiraju ili spajaju (anastomozma) prema vrhu. Obično mnogo manjih malih vena povezuje ove primarne vene, ali mogu završiti I vrlo finim venskim završecima u mezofilu. Male vene su tipičnije za angiosperme koje mogu imati čak četiri viša reda.[14] } Nasuprot tome, kod listova s mrežastom venacijom postoji jednostruka (ponekad i više njih) primarna vena u sredini lista, koja se naziva srednje rebro ili skelet, a kontinuirana je sa vaskulaturom bliže peteljke. Srednjinerv se potom grana na niz manjih sekundarnih, poznatih i kao vene drugog reda, koje se protežu prema ivicama listova. One se često završavaju u hidatodama, sekretornim organima, na margini. Zauzvrat, manje vene granaju se iz sekundarnih vena, poznatih kao vene trećeg (ili višeg reda), tvoreći gusti mrežasti obrazac. Područja ili otoci mezofila koji leže između vena višeg reda nazivaju se areole. Neke od najmanjih vena (veneleta = venica) mogu imati završetke u areolama, što je proces poznat kao areolacija.[13] Ove manje vene djeluju kao mjesta razmjene između mezofila i vaskularnog sistema biljke..[12] Tako male vene sakupljaju proizvode fotosinteze (fotosintate) iz ćelija u kojima se odvija, dok su glavne vene odgovorne za njihov transport van lista. Istovremeno se voda prnosii u suprotnom smjeru.[15][16][17]
Broj završetaka vena vrlo je promjenjiv, kao i da li se vene drugog reda završavaju na margini ili se vraćaju na ostale vene. Postoje mnoge složene varijacije uzoraka koje oblikuju lisne vene i ove imaju funkcionalne implikacije. Od njih imaju najveću raznolikost štitarice. U njima glavne vene djeluju kao mreža za podršku i distribuciju lišća i povezane su s oblikom lišća. Naprimjer, paralelna venacija koja se nalazi u većini monokotiledona korelira sa izduženim oblikom lišća i širokom lisnom osnovom, dok se mrežasta venacija vidi u jednostavnim cijelim listovima, a prstasti listovi obično imaju venaciju u kojoj se tri ili više primarnih vena radijalno razilaze od jedne tačke.
U evolucijskom smislu, rane nastajući rani taksone imali su tendenciju da kasnije nastaju dihotomna grananja sa mrežicama. U žilama su se pojavile vene u permskom period (prije 299–252 miliona godina), prvo kod angiospermi u trijasu (prije 252–201 miliona godina), tokom kojeg venska hijerarhija izgleda da omogućava veće funkcije, većih dimenzija lista i prilagođavanje raznim klimatskim uslovima.[14] Iako je to složeniji obrazac, čini se da su granaste vene pleziomorfne i u nekom obliku bile prisutne u drevnim sjemenjačama još prije 250 miliona godina. Pseudomrežasta venacija koja je zapravo vrlo modificirana perastoparalelna je autapomorfija kod nekih pripadnika porodice ‘’ Melanthiaceae’’, koji su monokotiledonr; npr. Paris quadrifolia. Na lišću s mrežaste nervature formiraju matricu skela koja listovima daje mehaničku čvrstinu.[18]
Morfološke promijene unutar jedne biljke
[uredi | uredi izvor]- Homoblastija
- Karakteristika u kojoj biljka ima male promjeneu veličini, obliku i obrazaca rasta listova između juvenil nihi odraslih stadija, za razliku od pojave zvane:
- Heteroblastija
- u kojoj biljka ima značajne promjene u veličini, obliku i a rasta i načina rasta među stadijima juvenila i odraslih.
Anatomija (srednje i male skale)
[uredi | uredi izvor]Obilježja srednje skale
[uredi | uredi izvor]Listovi su obično jako vaskularizirani i obično imaju mreže vaskularnih snopova koji sadrže ksplem, koji vodom opskrbljuje fotosintezu, i floem, koji prenosi šećere proizvedene tokom fotosinteze. Mnogi su listovi prekriveni trihomima (malim dlačicama) različitih struktura i funkcija.
Obilježja srednje skale
[uredi | uredi izvor]Listovi su obično jako vaskularizirani i obično imaju mreže vaskularnih snopova koji sadrže ksilem, koji opskrbljuje vodom fotosinteza, i floem, koji prenosi šećere proizvedene fotosintezom. Mnogi su listovi prekriveni trihomima (malim dlačicama) različitih struktura i funkcija.

Obilježja male skale
[uredi | uredi izvor]Glavni prisutni tkivni sistemi su:
- epiderma, koji pokriva gornju i donju površinu lista;
- Mezofilno tkivo (mezofil) unutar lista, koji je bogat hloroplastima (koji se nazivaju i hlorenhim)
- Raspored vena/nervature (vaskularno tkivo)
Ova tri tkivna sistema obično formiraju redovnu organizaciju na ćelijskoj skali. Specijalizirane ćelije koje se značajno razlikuju od okolnih i koje često sintetiziraju specijalizirane proizvode poput kristala, nazivaju se idioblasti.[19]

Glavna tkiva lista
[uredi | uredi izvor]-
 Poprečni presjek lista
Poprečni presjek lista -
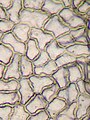 Ćelije epiderme
Ćelije epiderme -
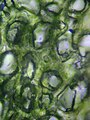 Spužvaste mezofine ćelije
Spužvaste mezofine ćelije



Epiderma
[uredi | uredi izvor]
epiderma je vanjski sloj ćelija koji pokriva list. Prekriven je voštanim kutikulom koja je nepropusna za tekuću vodu i vodenu paru i tvori granicu koja razdvaja unutarnje biljne ćelije od vanjskog svijeta. Kutikula je u nekim slučajevima tanja na donjoj epidermi nego na gornjoj, i obično je gušća na lišću iz suhog podneblja u usporedbi s onima iz vlažnih klimata.[20] Epiderma ima nekoliko funkcija: zaštita od gubitka vode putem transpiracije, regulacija razmjene plinova i izlučivanje metaboličkih spojeva. Većina listova pokazuje dorzoventralnu anatomiju: Gornja (adaksijalna) i donja (abaksijalna) površina imaju nešto drugačiju konstrukciju i mogu služiti različitim funkcijama.
Tkivo epiderme uključuje nekoliko diferenciranih tipova ćelija; ćelije epiderme, epidermne dlake (trihomi), ćelije u stomimog komplekau; čuvarske ćelije i pomoćne ćelije. Ćelije epiderme su najbrojnije, najveće i najmanje specijalizirane i čine njenu većinu. U listovima monokotiledona obično su više izduženi nego u listovima dikotiledona.
U ćelijama epiderme, hloroplasti su uglavnom odsutni, izuzev su zaštitnih ćelija stoma. Stomne pore perforiraju epidermu i okružene su sa svake strane zaštitnim ćelijma koje sadrže hloroplaste, te dvije do četiri pomoćne ćelije kojima nedostaju hloroplasti, tvoreći specijaliziranu ćelijsku skupinu poznatu kao stomni kompleks. Otvaranje i zatvaranje stomnog otvora kontrolira stomani kompleks i regulira razmjenu plinova i vodene pare između vanjskog zraka i unutrašnjosti lista. Stome zato imaju važnu ulogu u omogućavanju fotosinteze, a da se list ne osuši. U tipskom listu, stome su mnogobrojnije u abaksialnoj (donjoj) epidermi od adaksialne (gornje) i mnogo su brojniji u biljkama iz hladnijih klimata.
Mezofill
[uredi | uredi izvor]Veći dio unutrašnjosti lista između gornjeg i donjeg sloja epiderme je parenhim (temeljno tkivo) ili hlorenhim, tkivo koje se naziva mezofil (grčki naziv za "srednji list"). Ovo asimilacijsko tkivo je primarna lokacija fotosinteze u biljci. Proizvodi fotosinteze nazivaju se "asimilati".
U paprati i većini cvjetnica, mezofil je podijeljen u dva sloja:
- Gornji palisadni sloj čine vertikalno izdužene ćelije, debljine jedna do dvije ćelije, neposredno ispod adaksijalne epiderme, s međućelijskim zračnim prostorima između njih. Njegove ćelije sadrže mnogo više hloroplasta od sunđerastog sloja. Ove duge cilindrične ćelije su redovno raspoređene u jednom do pet redova. Cilindrične ćelije, sa „hloroplastima“ blizu ćelijskih zidova, mogu iskoristiti optimalnu prednost svetlosti. Lagano odvajanje ćelija omogućava maksimalnu apsorpciju ugljik-dioksida. Osunčano lišće ima višeslojni palisadni sloj, dok su sjenoviti ili stariji listovi bliže tlu jednoslojni.
- Ispod palisadnog, nalazi se spužvasti sloj. Ćelije spužvastog sloja su više razgranate i nisu toliko nabijene, tako da između njih postoje veliki međućelijski zračni prostori za kisik i ugljik-dioksid koji difundiraju u i iz njih tokom disanja i fotosinteze. Ove ćelije sadrže manje hloroplasta nego one palisadnog sloja. Pore ili „stome“ epiderme otvaraju se u substomne komore, koje su povezane u međućelijske zračne prostore između spužvastih i palisadnih mezofilnih ćelija.
Listovi su obično zeleni, zbog hlorofila u hloroplastims u mezofilnim ćelijama. Biljke kojima nedostaje hlorofil ne mogu obavljati fotosintezu.
Razvoj lista
[uredi | uredi izvor]Prema teoriji Agnes Arber delimičnih izdanaka lišća, listovi su i definirani kao djelimični izdanaci i potiču iz listova primordijalnog vrha izdanka.[21]
Složeni listovi bliži su izdancima nego jednostavni. Istraživanja razvoja pokazala su da se složeni listovi, poput izdanka, mogu granati u tri dimenzije.[22][23] Na osnovu molekularne genetike, Eckardt i Baum (2010) su zaključili da je "sada opšte prihvaćeno da složeni listovi ispoljavaju i i lisna i izdanačka svojstva."[24]
Ekologija
[uredi | uredi izvor]Biomehanika
[uredi | uredi izvor]Biljke reagiraju i prilagođavaju se faktorima okoliša, kao što su svjetlo i mehanički stres od vjetra. Listovi trebaju podržati vlastitu masu i uskladiti se na takav način da optimiziraju izlaganje suncu, uglavnom manje ili više vodoravno. Međutim, horizontalno poravnanje maksimizira izlaganje silama savijanja i vodi neuspjehu uslijed naprezanja tokom vjetra, snijega, tuče, padajućih krhotina, životinja i abrazije sa okolnog lišća i biljnih struktura. Listovi svih tipova su relativno lepršavi, s obzirom na ostale biljne strukture poput stabljika, grana i korijena.[25] Plojka lista i struktura peteljki utiču na reakciju lista na sile poput vjetra, dopuštajući stupanj premještanja da bi se minimiziralo povlačenje] i oštećenja, za razliku od otpora. Pomicanje listova poput ovog također može povećati turbulenciju zraka blizu površine lista, koji razrjeđuje granični sloj zraka odmah pokraj površine, povećavajući kapacitet za izmjenu plina i topline, kao i fotosinteze. Snažne sile vjetra mogu rezultirati smanjenim brojem listova i površine lista, što uz smanjenje pokretljivost, uključuje odstupanje i smanjuje intenzite fotosinteze. Stoga dizajn listova može uključivati kompromis između povećanja dotoka ugljika, termoregulacije i gubitka vode s jedne strane, i troškova održavanja statičkih i dinamičkih opterećenja. U vaskularnim biljkama, okomite sile se šire na većem području i relativno su fleksibilne pri savijanju i torziji], omogućujući elastično deformiranje bez oštećenja.[25]
Mnogi se listovi oslanjaju na hidrostatičke nosače smještene oko skeleta vaskularnog tkiva zbog svoje snage, što ovisi o održavanju stanja vode u listovima. I mehanika i arhitektura lista odražavaju potrebu za transportom i podrškom. Read i Stokes (2006) uzimaju u obzir dva osnovna modela, "hidrostatičkog" i oblik "čvrstog snopa" (vidi sliku).[25] Hidrostatički listovi kao u Prostanthera lasianthos su krupni i tanki i mogu uključivati potrebu za višestrukim lišćem, a ne samo jednim velikim listom, zbog količine vena potrebnih za podržavanje periferije velikih listova. Ali velike dimenzije listovaa pogoduju efikasnosti u fotosintezi i očuvanju vode, što uključuje daljnje kompromise. S druge strane, listovi kratkog snopa, poput onog kod vrste Banksia marginata, uključuju specijalizirane strukture ukrućivanja. Ove I-zrake nastaju od proširenja omotača sklerenhimskog snopa, koji se uklapa u ukrućene subepidermne slojeve. To prebacuje ravnotežu sa oslanjanja na hidrostski pritisak na strukturnu potporu, što je očita prednost tamo gdje je količina vode relativno mala.{{sfn|Read|Stokes|2006}][25]
Interakcije sa ostalim organizmims
[uredi | uredi izvor]
Iako nije tako hranjivo kao i drugi organi kao što su plodovi, lišće pruža izvor hrane za mnoge organizme. List je biljni vitalni izvor proizvodnje energije, a biljke su razvile zaštitu od životinja koje ga konzumiraju, poput tanina, hemikalija koje ometaju varenje proteina i imaju neugodan okus. Životinje koje su specijalizirane za jelo lišća su poznate kao folivori.
vrste imaju prilagodbe pomoću kojih koriste lišće u izbjegavanju predatora. Naprimjer, gusjenica porodice Tortricidae , od uvaljanog lišća stvorit će malu kućicu, presavijajući je nad sobom. Neke Pamphiliidae na sličan način prevrću lišće biljaka i hrane se lišćrm, savijenim u tube. Ženke Attelabidae, takozvanih valjaoničara njime, polažu jaja u lišće koje potom uvaljaju, kao zaštitno sredstvo. Ostali biljojedi i njihovi grabežljivci oponašaju izgled lista. Gmizavci, poput nekih kameleona i insekti slični nekim katidama također oponašaju oscilirajuće pokrete lišća na vjetru, krećući se sa strane naprijed ili naprijed i naprijed, izbjegavajući moguću prijetnju.
Sezonsko gubljene listova
[uredi | uredi izvor]
Lšće u umjerenom, borealnom ekosistemu i sezonski suhim zonama može biti sezonsko (listopadno; opadati ili umirati tokom nepovoljne sezone za vegetaciju). Ovaj mehanizam za odbacivanje lišća naziva se apscizija. Kada se list prorijedi, na grančici ostaje njihov ožiljak. U hladnim jesenima se ponekad promijene boju i pretvore u žutu, svijetle, kao što su narandžasta], ili crvena, kao različiti dodatni pigmenti karotenoidi i ksantofili ispoljavajuju se kada drvo reaguje na hladnoću i smanjuje sunčeva svjetlost] ograničavanjem proizvodnje hlorofila. Smatra se u listu dok ugiba stvaraju crveni antocijaninski pigmenti, moguće da prikrivaju žutu nijansu preostalu kada se izgubi hlorofil, pa izgleda da žuto lišće privlači biljojede poput lisnih ušiju.[26] Optičkim maskiranjem hlorofila antocijanima, smanjuje se rizik od foto-oksidativnog oštećenja lisnih ćelija dok stare, što u suprotnom može smanjiti efikasnost pronalaženja hranjivih sastojaka iz jesenskog lišća koje ostarijava[27]
Evolucijske adaptacije
[uredi | uredi izvor]
U toku evolucije listovi su se prilagodili različitim biofizičkim okruženjima na sljedeće načine:
- Epikutikulski vosak: voštani mikro- i nanostrukture na površini lista smanjuju vlaženje kišom i prijanjanje onečišćenja ( Vidi Lotus efekt ).
- Podeljeni i složeni listovi smanjuju otpornost na vetar i podržavaju hlađenje.
- Dlake na površini listova zadržavaju vlažnost u suhim klimama i stvaraju granični sloj, smanjujući gubitak vode.
- Vosak biljne kutikule smanjuju gubitak vode.
- Velika površina pruža veliko područje za hvatanje sunčeve svjetlosti.
- U štetnim količinama sunčeve svjetlosti, specijalizirani listovi, neprozirni ili djelomično zakopani, propuštaju svjetlost kroz prozirni prozora listova za fotosintezu na unutrašnjim površinama listova (npr. Fenestraria ).
- Sukulente imaju skladište vode i organskih kiselina za upotrebu u fotosintezi CAM.
- Aromatsko ulje, otrovi ili feromoni proizvedeni od lisnih žlijezda koje odstranjuju biljojede (npr. Eucaliptus).
- Uključivanja kristalnih minerala odvraćaju biljojedi (npr. silikat, fitoliti trava, rafidi kod Araceae).
- Prašnici privlače oprašivače.
- Bodlje štite biljke od biljojeda (npr. kaktusi).
- Dlake služe za zaštitu od biljojeda, npr. u Urtica dioica i Dendrocnide moroides (Urticaceae).
- Posebni listovi mesožderki su prilagođeni za hranjenje hrane, uglavnom beskičmenjaka, iako neke vrste hvataju i male jedinke (vidi biljka mesožderka).
- Lukovica čuva hranu i vodu (npr. lukovi).
- Vitice omogućuje biljci da se uspne (npr. grašak).
- Brakteje i pseudanttiji (lažni cvjetovi) zamjenjuju normalne cvjetne strukture, kada se istinski cvjetovi uveliko smanje (npr. „mlijeko“ i smole u Araceae .
Terminologija
[uredi | uredi izvor]
Oblik
[uredi | uredi izvor]
Rubovi – Margine
[uredi | uredi izvor]| Slika | Termin | Latinski | Opis |
|---|---|---|---|
| Forma integra |
Čak; sa glatkom marginom; bez ozubljenja | ||
| Ciliatna | Cilliate | Ograničena dlačicama | |
| Krenatna | Crenate | Talasasta; ozubljena sa zaobljenim zupcima | |
| Dentatna | Dentata | Nazubljena,
može biti grubo udubljena, sa velikim zupcima ili sa zupcina sa žlijezdama | |
| Dentikulatna | Denticulata | Fino nazubljena | |
| Dvostruko nazubljena | Duplcato-dentata | Svaki zubac nosi manje zupce | |
| Nazubljeni | Seratni | Nazubljeni; sa asimetričnim zupcima okrenutim prema naprijed | |
| Sitno nazubljena | Serulatni | Fino nazubljeni | |
| Sinuatna | Sinuosa | Sa dubokim udubinama, nalik talasima; grubo krenatni | |
| Režnjevita | Lobata | Utisnuti, sa urezima da ne dosegnu središte | |
| Undulatna | Undulatna | S valovitim rubom, plitkije od sinutastog | |
| Trnasta ili oštrat | Spikulatna(bockasta) | Sa ukočenim, oštrim tačkama kao što je čičak |
Apeks – Vrh
[uredi | uredi izvor]| Slika | Termin | Latinski | Opis |
|---|---|---|---|
| Akuminatns | Acuminate | Dugošiljat, produžen u usku, suženu tačku na konkavan način | |
| Akutna | Acute | Završava u oštroj, ali ne produženoj tački | |
| Nazubljena | Cuspidate | S oštrim, izduženim, čvrstim vrhom; prekriven vrhom | |
| Urezana | Emarginate | Udubljen, s plitkim zarezom na vrhu | |
| Mukronatna | Mucronate | Naglo završava u malu, kratku tačku | |
| Mukronulatna | Mucronulate | Mukronatni, ali s primjetno umanjenom bodljom | |
| Srcasta | Obcordate | Obrnuto srcast | |
| Zatupljena | Obtusate | Zaobljen ili tup | |
| Skraćena | Truncate | Završava naglo ravnim krajem |
Baza
[uredi | uredi izvor]- Akuminatna
- Dolazi do oštre, uske, produžene tačke.
- Akutna
- Dolazi do oštre, ali ne produžene tačke.
- Aurikulatna
- U obliku uha.
- Kordatna
- Srcasta, sa zarezom prema stabljici.
- Kuneatna
- Klinasta
- Hastatna
- Oblikovana dlanoliko, sa baznim režnjem usmjerenim prema vani.
- Oblikusna
- Kosa.
- Reniformna
- Bubrežasta, ali zaobljena i šira od dugog.
- Zaobljena
- Zakrivljena
- Sagitatna
- Streličasta, s oštrim baznim kukama usmjerenim prema dolje.
- Trunkatna
- Naglo završava s ravnim krajem, koji izgleda kao odsječen.
Površina
[uredi | uredi izvor]- Koriacozna
- Kožasta, kruta i čvrsta, ali pomalo fleksibilna.
- Farinozna
- Nositelj farina; brašnasta, prekrivena voštanim, bjelkastim prahom.
- Glabrozna
- Glatka, ne dlakava.
- Glaukouzna
- S bjelkastim pokrovom; prekrivena vrlo finim, plavkasto-bijelim prahom.
- Ljepljiva
- Vidno ljepljiva.
- Lepidotna
- Obložena malim ljuskama (tako elepidote ', bez takvih vaga).
- Koriaceozna
- : Kožasta; kruta i čvrsta, ali pomalo fleksibilna.
- Farinozna
- Nositelj farina; brašnast, prekriven voštanim, bjelkastim prahom.
- Glabrozna
- Glatka, ne dlakava.
- Glaukozna
- S bjelkastim slojem; prekrivena vrlo finim, plavkasto-bijelim prahom.
- Ljepljiva
- Ljepljiva, vidljivo.
- Lepidotna
- Obložena malim ljuskama (elepidote, bez takvih ljuski).
- Makulatna
- Maljava, mrljast, uporedite imakulatna'.
- Papilatna ili papilozna
- nosi papile (male izbočine u obliku bradavice).
- Pubescentna
- Prekrivena uspravnim dlačicama (posebno mehkim i kratkim).
- Punktatna
- Označena tačkicama; isprekidana udubljenjima ili prozirnim žlijezdama ili obojenim tačkicama.
- Rugozna
- duboko naborana, s jasno visdljivom nervaturom.
- Perutava
- Prekrivena je sitnim česticama širokih razmjera.
- Tuberkulatna
- Prekrivena kvržicama bradavicama.
- Verukozna
- brušena, s bradavicama.
- Viskozna
- Prekrivena gustim, ljepljivim sekretima.
Površina lišća je također domaćin velikog broja mikroorganizama; u ovom kontekstu naziva se filosfera.
Dlakavost
[uredi | uredi izvor]

"Dlake" na biljkama pravilno se nazivaju trihomi. Listovi mogu ispoljavati nekoliko stupnjeva dlakavosti. Značenje nekoliko sljedećih izraza može se preklapati.
- Arahnoidne ili arahnozne
- S mnogo finih, zapletenih dlaka koje daju oblik paučine.
- Barbelatne
- sa sitno bodljikavim dlačicama (mrene).
- Bradaste
- Sa dugim ukočenim dlačicama.
- Čekinjaste
- Sa ukočenim bodljama nalik na kosu.
- Kanescentne
- osip sa gustom sivkasto-bijelom dlakavošću.
- Cilijatne
- Margine brubljene kratkim dlačicama (cilijams).
- Ciliolatne
- Minimalne cilije.
- Flocozne
- Sa grupama mekanih, vunenih dlaka koje imaju tendenciju odvajanja.
- Glabrescentne
- Gubitak dlaka s godinama.
- Glabrozne
- Nema dlačica bilo koje vrste.
- Žljezdanae
- Sa žlijezdom na vrhu dlake.
- Hirsutne
- S prilično hrapavim ili ukočenim dlačicama.
- Hispidne
- Sa krutim, čekinjastim dlačicama.
- Hispidulozne
- Minimalno hispidne.
- Hoarne
- S finom, tijesnom sivkasto-bijelom dlakom
- Lanatne, ili lanozne
- Sa vunastim dlačicama.
- Pilozne
- Sa mekim, jasno odvojenim dlačicama.
- Puberulentne
- s+Sa sitnim, malim dlačicama.
- Pubescentne
- sa mekim, kratkim i uspravnim dlačicama.
- Skabrozne ili škakljaste
- Grube na dodir.
- Sericeozne
- Svilenkastog izgleda, fine, ravne i utisnute (uske i ravne) dlake.
- Svilenkaste
- Sa utisnutim, mekim i ravnim dijelom dlakavosti.
- Zvjezdane ili stelliformne
- Sa dlakama u obliku zvijezde.
- Strigozne
- Sa ukočenim, oštrim, ravnim i uzdignutim dlačicama.
- Tomentozne
- Guste, sa matiranim, mekim bijelim dlakavim dlačicama. Između kanescentne i tomentozne
- Tomentulozne
- Minimalno ili samo blago tomentozne.
- Villozne
- Sa dugim i mekim dlačicama, obično zakrivljenim.
- Vunaste
- Sa dugim, mekim i kovrčavim ili matiranim dlačicama.
Vrijeme pojave
[uredi | uredi izvor]Provodno (vaskularno) tkivo
[uredi | uredi izvor]
Nervatura lista je vaskularno tkivo lista i locirana je u spužvastom sloju mezofila. Obrazac sistema vena naziva se venacija ili nervatura. U angiospermi venacija je tipski paralelna u monokotiledons, a tvori povezanu mržu u biljkama širokog lišća. Nekad se smatralo da su tipski primjeri formiranje obrasca putem ramifikacije, ali umjesto toga mogu se uočitii obrasci formirani u stresnom tenzorskom polju. [30][31][32] Vena je sastavljena od vaskularnog (provodnog) snopa. U jezgri svakog snopa su grozdovi od dvije različite vrste vodećih ćelija:
- Ksilem
- Ćelije koje donose vodu i minerale iz korijena u list.
- Floem
- ćelije koje se obično prenose, van lista sokove, sa rastvorenom saharozom (glukozom u saharozi) nastalom fotosintezom u listu.
Ksilem je obično na adaksiksijalnoj strani vaskularnog snopa, a floem obično na abaksiksalnoj strani. Oba su ugrađena u gusto tkivo parenhima, nazvano omotač, koje obično uključuje neko strukturno tkivo kolenhima.
Također pogledajte
[uredi | uredi izvor]Reference
[uredi | uredi izvor]- ^ Tsukaya 2013.
- ^ Feugier 2006.
- ^ Purcell 2016.
- ^ a b c d Stewart i Rothwell 1993.
- ^ Marloth 1913–1932.
- ^ Bayer 1982.
- ^ Krogh 2010.
- ^ James i Bell 2000.
- ^ Heywood et al 2007.
- ^ Hallé 1977.
- ^ Rolland-Lagan et al 2009.
- ^ a b Walls 2011.
- ^ a b Roth-Nebelsick et al 2001.
- ^ a b Sack i Scoffoni 2013.
- ^ Ueno et al 2006.
- ^ Rudall 2007.
- ^ Dickison 2000.
- ^ Bagchi, Debjani; Dasgupta, Avik; Gondaliya, Amit D.; Rajput, Kishore S. (2016). "Insights from the Plant World: A Fractal Analysis Approach to Tune Mechanical Rigidity of Scaffolding Matrix in Thin Films". Advanced Materials Research. 1141: 57–64. doi:10.4028/www.scientific.net/AMR.1141.57.
- ^ Cote 2009.
- ^ Clements, Edith Schwartz (1905). "The Relation of Leaf Structure to Physical Factors" (PDF). Transactions of the American Microscopical Society. 26: 19–98. doi:10.2307/3220956. hdl:2027/uc2.ark:/13960/t5v699n4k. JSTOR 3220956.
- ^ Simpson 2011, p. 356.
- ^ Rutishauser i Sattler 1997.
- ^ Lacroix et al 2003.
- ^ Eckardt i Baum 2010.
- ^ a b c d Read i Stokes 2006.
- ^ Doring et al 2009.
- ^ Feild et al 2001.
- ^ Kew Glossary: Hysteranthous
- ^ Kew Glossary: Synanthous
- ^ Couder et al 2002.
- ^ Corson et al 2009.
- ^ Laguna et al 2008.
Vanjski linkovi
[uredi | uredi izvor]| Discipline | |
|---|---|
| |
| Rast i habitus (izgled) | |
| Razmnožavanje | |
| Taksonomija biljaka | |
| Primjena | |
| |