


Cancers, Volume 11, Issue 12 (December 2019) – 223 articles
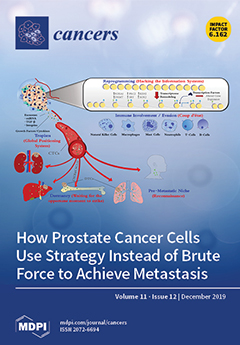
Cover Story (view full-size image):
Despite being an extremely inefficient process, the high burden of lethality in metastatic prostate cancer patients can be attributed to a plethora of strategies utilized by the cancer cells that have escaped the primary tumor. Striking similarities exist between the means used to achieve metastasis and military combat, where oftentimes, the modus operandi involves the creative use of tactics to overcome the host systems. These strategies include priming the metastatic sites ahead of colonization, targeting specific organs, outsmarting the host immune surveillance, lying in a dormant state at the metastatic site for prolonged periods, and widespread reprogramming of gene expression. This review provides a perspective for understanding metastasis from the standpoint of military combat, wherein strategic maneuvering instead of brute force often plays a decisive role in the outcome. View this paper
- Issues are regarded as officially published after their release is announced to the table of contents alert mailing list.
- You may sign up for e-mail alerts to receive table of contents of newly released issues.
- PDF is the official format for papers published in both, html and pdf forms. To view the papers in pdf format, click on the "PDF Full-text" link, and use the free Adobe Reader to open them.
Previous Issue
Next Issue