


Int. J. Mol. Sci., Volume 16, Issue 9 (September 2015) – 165 articles , Pages 20100-23126
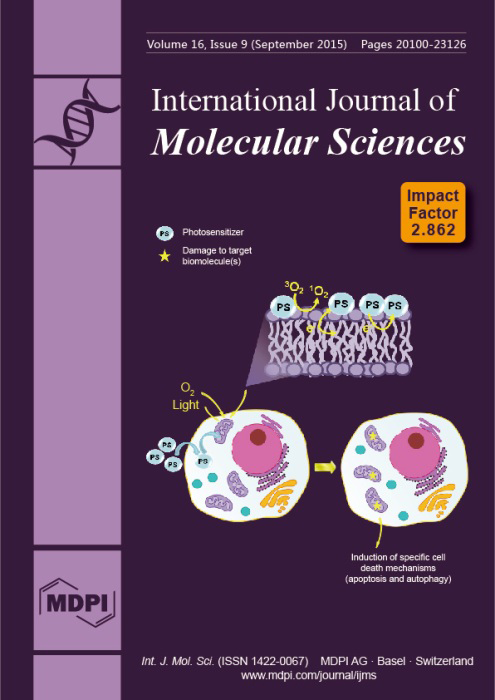
Cover Story:
Photosensitizers (PS) are the main agents for Photodynamic therapy (PDT). Still, photosensitizers are usually considered simple singlet oxygen generators. By analyzing the molecular and biological processes taking place during and after photosensitization, we aim to suggest alternatives for achieving high-efficiency PDT protocols. We submit that PSs should be designed to induce specific mechanisms of cell death and researchers should first consider tissue and intracellular localization, instead of trying to maximize the generation of reactive species. View the article.
- Issues are regarded as officially published after their release is announced to the table of contents alert mailing list.
- You may sign up for e-mail alerts to receive table of contents of newly released issues.
- PDF is the official format for papers published in both, html and pdf forms. To view the papers in pdf format, click on the "PDF Full-text" link, and use the free Adobe Reader to open them.
Previous Issue
Next Issue