

Luân phiên thế hệ

Luân phiên thế hệ (còn gọi là xen kẽ thế hệ, metagenesis hay heterogenesis)[1] là kiểu vòng đời của tất cả thực vật trên cạn và nhiều loài tảo; miêu tả sự tồn tại của hai giai đoạn riêng biệt hữu tính đơn bội và vô tính lưỡng bội. Sự luân phiên được nhắc đến là sự luân phiên giữa hai dạng sống: thể giao tử (n) và thể bào tử (2n). Trong đó n và 2n thể hiện mức bội thể.
Có thể tóm tắt quá trình đó như sau: thể bào tử (2n) → bào tử (n) → thể giao tử (n) → giao tử (n) → thể bào tử (2n). Trong đó, những dạng sống có chữ "thể" là đa bào, không có chữ "thể" là đơn bào và là đơn vị sinh sản của dạng sống có chữ "thể".
Quá trình 2n --> n là giảm phân, quá trình n --> n và 2n --> 2n là nguyên phân. Tế bào n không thể tự thân tạo thành tế bào 2n mà phải cộng với một tế bào n khác, gọi là quá trình dung hợp. Nguyên phân là sự phát triển từ đơn vị sinh sản thành cơ thể.
Tương quan giữa thể bào tử và thể giao tử không giống nhau ở tất cả thực vật.
- Tảo: Thể bào tử và thể giao tử bình đẳng và sống tách biệt hẳn khỏi nhau; ngoại hình của hai thế này có thể giống nhau hoặc không.
- Rêu (rêu, rêu sừng và rêu tản): Thể giao tử là dạng sống cơ bản, thể bào tử kém phát triển hơn và sống phụ thuộc vào thể giao tử. Dù thể bào tử của rêu và rêu sừng vẫn chưa mất khả năng quang hợp, chúng vẫn cần được thể giao tử bổ sung chất dinh dưỡng, nước, khoáng và nitơ mới có thể phát triển và tạo bào tử.[2][3]
- Quyết: Thể bào tử là dạng sống cơ bản, thể giao tử chỉ còn là một nguyên tản tự dưỡng. Nguyên tản này phụ thuộc vào thể bào tử nhưng không hoàn toàn.
- Thực vật hạt kín: Thể bào tử là dạng sống cơ bản, sự thoái hoá của thể giao tử cực kì dữ dội: chúng chỉ còn là một vài tế bào phát triển hoàn toàn bên trong thể thể bào tử.
Động vật thì sinh sản theo một cách khác hoàn toàn. "Thể bào tử" của chúng ta trực tiếp tạo ra giao tử đơn bội, không hình thành bào tử đơn bội nên không có pha đơn bội đa bào. Nói cách khác, khả năng nguyên phân không hiện diện ở những tế bào đơn bội được tạo ra trong quá trình sinh sản của động vật. (Một số côn trùng có hệ thống xác định giới tính trong đó trứng chưa thụ tinh sẽ nở thành con đực đơn bội, còn trứng đã thụ tinh nở thành con cái lưỡng bội).
Chu kì sống của thực vật và tảo có sự luân phiên giữa kì đơn bội đa bào và lưỡng bội đa bào được gọi là chu kì lưỡng-đơn bội. Chu kì sống không có sự luân phiên đó mà chỉ có một kì lưỡng bội đa bào (ví dụ như ở động vật) được gọi là chu kì lưỡng bội. Chu kì sống chỉ có một kì đơn bội đa bào được cọi là chu kì đơn bội.
Định nghĩa
[sửa | sửa mã nguồn]Luân phiên thế hệ được định nghĩa là sự luân phiên giữa thể lưỡng bội và thể đơn bội trong một chu kì sống, hai thể đó có thể sống phụ thuộc hoặc độc lập khỏi nhau.[4] Trong một số loài, ví dụ như loài tảo Ulva lactuca, thể đơn và lưỡng bội có thể sống hoàn toàn tách biệt khỏi nhau, và vì chúng có vẻ ngoài giống hệt nhau nên được gọi là đẳng hình (hoặc đồng hình). Hai giao tử đơn bội sẽ hợp với nhau tạo thành hợp tử, hợp tử nảy mầm thành thể lưỡng bội đa bào. Thể lưỡng bội giảm phân tạo bào tử đơn bội, bào tử đơn bội nảy mầm thành thể giao tử đơn bội.[5]
Tuy nhiên, trong một số nhóm khác, một trong hai thể bị thoái hoá và phải sống phụ thuộc vào thể kia. Ví dụ, tất cả loài rêu đều có thể giao tử là trội, tức là thể bào tử bị sống phụ thuộc. Ngược lại, tất cả động vật có mạch có thể bào tử là trội và thể giao tử sống phụ thuộc, nhưng những bằng chứng hoá thạch cho thấy chúng tiên hoá từ tổ tiên có hai thể đồng hình.[6] Ở thực vật có hạt, thể giao tử phát triển bên trong cơ thể của thể bào tử, giúp bảo vệ và nuôi dưỡng nó cũng như sản phẩm của nó (hợp tử) tốt hơn. Hạt phấn là thể giao tử đực thoái hoá chỉ còn một vài tế bào (trong nhiều trường hợp là 3). Ở những loài này, sự tách biệt giữa hai thể hệ không rõ, như Bateman & Dimichele nói "có thể coi rằng thể bào tử và thể giao tử hoạt động như một sinh vật duy nhất".[7] Thuật ngữ "Luân phiên các pha" sẽ mô tả hiện tượng này chính xác hơn.[8]
Lịch sử
[sửa | sửa mã nguồn]Các cuộc tranh luận về sự luân phiên thế hệ vào đầu thế kỉ XX khá rối rắm vì có rất nhiều hệ thống chia "thế hệ" cùng tồn tại (hữu tính và vô tính, thể giao tử và thể bào tử, thể đơn bội và thể lưỡng bội,...).[9]
Ban đầu, khi nghiên cứu về sinh sản của phân ngành Sống đuôi, ngành Thích ty bào và lớp Trematoda, Chamisso và Steenstrup dùng thuật ngữ "luân phiên thế hệ" để miêu tả sự kế thừa giữa các thế hệ động vật (vô tính và hữu tính). Hiện tượng này ngày nay được gọi là dị giao, còn thuật ngữ "luân phiên thế hệ" chỉ còn được dùng cho thực vật, cụ thể là sự luân phiên giữa thể giao tử đơn bội và thể bào tử lưỡng bội.[9]
Wilhelm Hofmeister đã chứng minh có sự khác biệt về hình thái trong sự luân phiên thế hệ ở thực vật,[10] giữa một thế hệ mang bào tử (thể bào tử) và một thế hệ mang giao tử (thể giao tử).[11][12] Trong thời gian đó, đã có một cuộc tranh luận giữa hai luồng ý kiến, sự luân phiên thế hệ xen kẽ (Čelakovský, 1874) và tương đồng (Pringsheim, 1876).[9] Čelakovský cũng là người tạo ra từ "thể bào tử" và "thể giao tử".[cần dẫn nguồn]
Eduard Strasburger (1874) đã phát hiện ra sự luân phiên giữa thế hệ lưỡng bội và đơn bội,[9] còn được gọi là sự luân phiên tế bào học của pha nhân.[13] Dù nhiều khi chúng đồng bộ nhau, đôi khi sự luân phiên về hình thái không diễn ra cùng lúc với sự luân phiên về nhân. Trong nhiều loài tảo đỏ, một pha nhân có thể chứa hai pha hình thái.[13] Còn trong nhiều loài dương xỉ, cụ thể là những loài không có sinh sản hữu tính, sự thay đổi về pha nhân không diễn ra, nhưng vẫn có sự thay đổi pha hình thái nên vẫn có sự luân phiên thế hệ.[14]
Sự luân phiên thế hệ ở thực vật
[sửa | sửa mã nguồn]Các yếu tố cơ bản
[sửa | sửa mã nguồn]Lược đồ ở đầu trang miêu tả các yêu tố cơ bản của sự luân phiên thế hệ ở thực vật. Những biến thể từ các yếu tố cơ bản này sẽ được mô tả kĩ càng hơn ở các mục sau. Bắt đầu từ phía bên phải, quá trình đó diễn ra như sau:[15]

- Hai giao tử đơn bào đều có bộ nhiễm sắc thể n dung hợp để tạo thành một hợp tử đơn bào lưỡng bội, có bộ nhiễm sắc thể 2n (có n cặp nhiễm sắc thể)
- Hợp tử đơn bào này nảy mầm, nguyên phân để giữ bộ nhiễm sắc thể 2n. Tạo ra một thể bào tử đa bào.
- Khi thể bào tử này trưởng thành, nó tạo ra một hoặc nhiều túi bào tử - một cơ quan có khả năng tạo ra các tế bào mẹ lưỡng bội (gọi là tế bào nguyên bào tử). Những tế bào này giảm phân (phân chia để lượng nhiễm sắc thế giảm còn một nửa, từ 2n xuống n). Tạo thành 4 bào tử đơn bội đơn bào.
- Bào tử đơn bội này nảy mầm trong điều kiện thích hợp, nguyên phân tạo thành một sinh vật đa bào đơn bội, gọi là thể giao tử (vì nó có khả năng tạo ra giao tử)
- Khi thể giao tử này trưởng thành, nó tạo ra một hoặc nhiều túi giao tử - một cơ quan có khả năng tạo ra các giao tử đơn bội. Ít nhất một loại giao tử có khả năng tiếp cận một giao tử khác để dung hợp.
Ta có thể thấy sự "luân phiên thế hệ" trong chu kì sống của thực vật là sự luân phiên giữa thế hệ lưỡng bội (2n, thể bào tử) và thế hệ đơn bội (n, thể giao tử).
Sự luân phiên này không diễn ra ở động vật. Quá trình cơ bản ở động vật là một thể lưỡng bội trục tiếp tạo ra giao tử đơn bội bằng hình thức giảm phân. Ở động vật, "bào tử" (tế bào đơn bội có khả năng nguyên phân) không được tạo ra, cho nên không xuất hiện thế hệ đa bào, vô tính. Một số côn trùng có cơ chế xác định giới tính bằng mức bội thể; theo đó, thể lưỡng bội (nở ra từ trứng đã thụ tinh) là con cái và thể đơn bội (nở ra từ trứng chưa thụ tinh) là con đực.
Các biến thể
[sửa | sửa mã nguồn]Giản đồ ở đầu trang thể hiện tốt vòng đời của một số tảo đa bào (ví dụ như chi Cladophora), các tảo này có thể bào tử và thể giao tử giống hệt nhau về hình dáng; bào tử và giao tử cũng giống hệt nhau về gen.[16]
Tuy nhiên, mỗi yếu tố cơ bản của sự luân phiên thế hệ có nhiều biến thể. Mỗi biến thể lại có thể diễn ra tách biệt hoặc cùng nhau, nên có rất nhiều kiểu vòng đời khác nhau. Và vì thế những thuật ngữ mà các nhà thực vật học dùng để miêu tả các kiểu vòng đời này cũng rất gây bối rối. Như Bateman và Dimichele đã nói "sự luân phiên thế hệ đã trở thành một bãi lầy thuật ngữ; trong đó một từ có thể mang nhiều nghĩa và nhiều từ có thể mang cùng nghĩa"[17]


Các biến thể đó bao gồm:
- Về tầm quan trọng của thế bào tử và thể giao tử
- Bằng nhau (đồng hình hoặc đẳng hình).
Tảo sợi thuộc chi Cladophora, thường gặp ở vùng nước ngọt, có thể bào tử và thể giao tử gần như giống hệt nhau về hình dáng bên ngoài.[18] Không có thực vật trên cạn nào có thể giao tử và thể bào tử quan trọng bằng nhau, mặc dù một số thuyết cho rằng điều này đã diễn ra với một số thực vật trên cạn cổ đại. - Khác nhau (dị hình hoặc lệch hình)
- Thể giao tử trội.
Ở các loài rêu (rêu sừng, rêu tản và rêu thật sự), Thể trội hơn là thể giao tử, còn thể bào tử sống phụ thuộc vào thể giao tử mẹ của nó về dinh dưỡng và nước, chúng không có chất diệp lục khi trưởng thành.[19] - Thể bào tử trội.
Ở các loài quyết (bào gồm dương xỉ), cả thể bào tử và thể giao tử đều có thể sống độc lập, nhưng thể trội hơn là thể bào tử. Thể giao tử bị thoái hóa, trở nên nhỏ hơn và đơn giản hơn rất nhiều về cấu tạo.
Ở thực vật có hạt, thể giao tử càng thoái hóa hơn nữa (tối thiểu chỉ bao gồm 3 tế bào), phụ thuộc hoàn toàn vào thể bào tử về dinh dưỡng. Sự thoái hóa cực độ và sự phụ thuộc hoàn toàn của thể giao tử khiến cụm "luân phiên thế hệ" hơi không phù hợp cho thực vật có hạt: "thể bào tử và thể giao tử hoạt động như một sinh vật duy nhất".[7] Vì lí do đó, một số tác giả thích dùng cụm "luân phiên giai đoạn" hơn.[8]
Gametophyte of Pellia epiphylla with sporophytes growing from the remains of archegonia.
- Thể giao tử trội.
- Bằng nhau (đồng hình hoặc đẳng hình).
- Về sự phân hóa của giao tử
- Hai giao tử giống nhau (đẳng giao).
Như những loài khác của chi Cladophora, tất cả giao tử của loài C. callicoma đều có đuôi và đều có khả năng di chuyển.[18] - Hai giao tử có kích thước khác nhau (dị giao hoặc lệch giao)
- Cả hai đều có khả năng di chuyển.
Ở những loài của chi Tảo diếp, tất cả giao tử đều có đuôi và có thẻ di chuyển. Tuy nhiên, chúng có hai kích thước: giao tử 'cái' lớn hơn và giao tử 'đực' nhỏ hơn.[20] - Một lớn và không có đuôi, một nhỏ và có thể di chuyển (tức có đuôi) (noãn giao)
Giao tử lớn và không có đuôi là trứng (hoặc noãn), giao tử bé và có thể di chuyển là tinh trùng. Khả năng di chuyển của tinh trùng có thể rất hạn chế (ví dụ như trường hợp của thực vật có hoa) nhưng tất cả đều có khả năng chủ động tiếp cận trứng, theo cách này hoặc cách khác. Khi trứng và tinh trùng được tạo ra ở hai túi giao tử khác nhau, túi giao tử sản xuất ra tinh trùng được gọi là túi đực và túi sản xuất ra trứng gọi là túi cái.- Túi đực và túi cái tồn tại trên cùng cơ thể (đồng chu).[21][22] Loài rêu tản Pellia epiphylla có thể giao tử là thế hệ trội, nó cũng là loài đồng chu: túi đực nằm rải rác trên gân giữa của lá còn túi cái nằm gần đỉnh phân chia của cây.[23]
- Túi đực và túi cái tồn tại trên hai cơ thể khác nhau (biệt chu) Loài rêu Mnium hornum có thể giao tử là thể trội, nhưng nó là loài biệt chu: cây rêu đực chỉ có túi đực trên những thể hoa thị ở đỉnh, còn rêu cái chỉ có túi cái mang hình những cái nang có thân.[24] Thể giao tử của thực vật có hạt cũng biệt chu, nhưng thể bào tử của loài có thể biệt chu hoặc đồng chu. Thể giao tử của thực vật có hạt cực kì thoái hóa; túi cái của nó chỉ chứa một ít tế bào và toàn bộ thể giao tử đực của nó có thể chỉ có hai tế bào.[25]
- Cả hai đều có khả năng di chuyển.
- Hai giao tử giống nhau (đẳng giao).
- Về sự phân hóa của bào tử.
- Hai bào tử giống nhau (đẳng bào hay đồng hình bào tử).
Tất cả bào tử của dương xỉ đuôi ngựa đều có cùng kích thước.[26] - Hai bào tử có kích thước khác nhau (dị bào, lệch bào hay dị hình bào tử): Khi có hai loại bào tử sinh ra từ nhiều loại túi bào tử khác nhau, chúng sẽ được gọi là đại bào tử nang và tiểu bào tử nang. Một đại bào tử thường được sinh ra từ giảm phân cùng với ba thể cực, nhưng ba thể cực này "hi sinh" tiêu biến để dành dinh dưỡng cho chỉ một đại bào tử này.
- Đại bào tử nang và tiểu bào tử nang tồn tại trên cũng thể bào tử (đồng chu) Hầu hết thực vật hạt kín thuộc loại này. Ví dụ: hoa loa kèn có sáu nhị (tiểu bào tử nang) có khả năng tạo hạt phấn (tiểu thể bào tử); và ba nhụy chứa các noãn (đại bào tử nang) có khả năng tạo đại bào tử, từ đó tạo đại thể bào tử. Với một số thực vật khác, ví dụ như cây Phỉ, mỗi hoa chỉ có nhị hoặc nhụy, nhưng mỗi cây (một thể bào tử) đều có đủ hai loại hoa, cho nên chúng đồng chu.
- Đại bào tử nang và tiểu bào tử nang tồn tại trên các thể bào tử khác nhau (biệt chu) Một cá thể cây nhựa ruồi châu Âu có thể tạo ra hoa đực hoặc hoa cái, mỗi hoa chỉ có một loại bào tử nang. Hoa đực thì có nhị (tiểu bào tử nang) chỉ sản xuất tiểu bào tử có khả năng phát triển thành tiểu thể giao tử (hạt phấn); hoa cái thì có nhụy chứa đại bào tử nang (noãn) chứa đại bào tử có khả năng phát triển thành đại thể giao tử đa bào.
- Hai bào tử giống nhau (đẳng bào hay đồng hình bào tử).


Có một số sự liên quan giữa những biến thể này, nhưng chúng chỉ là liên quan, chứ không liên hệ tuyệt đối. Ví dụ, ở thực vật hạt kín, tiểu bào tử thường tạo ra tiểu giao tử (tinh) và đại bào tử tạo ra đại giao tử (noãn). Tuy nhiên, một số loài dương xỉ chỉ có một loại bào tử nhưng có nhiều loại thể giao tử. Ví dụ, loài Ceratopteris thalictroides có một loại bào tử duy nhất có mang nhiều kích thước khác nhau, thay đổi liên tục. Những bào tử nhỏ thường nảy mầm thành thể bào tử mang tính đực.[26]
Một chu kì sống phức tạp
[sửa | sửa mã nguồn]
Giản đồ bên dưới cho thấy sự luân phiên thế hệ của một loài dị hình, thể bào tử trội, noãn giao, biệt chu (cả về thể bào tử lẫn thể giao tử) và dị bào. Một ví dụ của loài như thế này là cây Liễu.[27] Từ giữa giản đồ, quá trình diễn ra như sau:
- Tinh trùng từ túi đực di chuyển đến trứng trong túi cái, hai giao tử này dung hợp tạo thành một hợp tử, hợp tử này là đực hoặc cái.
- Hợp tử đực nguyên phân thành tiểu thể bào tử có khả năng tạo tiểu bào tử nang khi trưởng thành. Tiểu bào tử nang sinh ra tiểu bào tử qua hình thức giảm phân.
- Ở cây Liễu, như các loài thực vật có hạt khác, hợp tử phát triển thành tiểu thể bào tử phôi bên trong noãn (một đại bào tử nang). Khi trưởng thành, cấu trúc này trở thành hạt của cây, hạt này sau đó được phát tán và nảy mầm thành một cây trưởng thành mới. Cây Liễu đực (tiểu bào tử thể) cho hoa chỉ có nhị, bao phấn ở đầu nhị là tiểu bào tử nang.
- Tiểu bào tử nảy mầm tạo thành tiểu giao tử thế; tiểu giao tử thể trưởng thành tạo ra túi đực, tạo ra tinh trùng.
- Ở cây Liễu, tiểu giao tử không được giải phóng khỏi bao phấn (tiểu bào tử nang) mà phát triển luôn thành hạt phấn (tiểu giao tử thể). Hạt phấn mới được phát tán (bằng côn trùng hay gió) đến noãn (đại giao tử thể), nơi tinh trùng được tạo ra và di chuyển theo ống phấn vào noãn cầu.
- Hợp tử cái nguyên phân thành đại bào tử thể có khả năng tạo tiểu bào tử nang khi trưởng thành. Đại bào tử nang sinh ra tiểu bào tử qua hình thức giảm phân; nhưng thường thì chỉ một trong bốn bào tử lấy phần nhiều tế bào chất, còn ba bào tử kia tiêu biến.
- Ở cây liễu, cây cái (đại bào tử thể) tạo hoa chứa lá noãn (lá biến dạng có chức năng giữ đại bào tử nang).
- Đại bào tử này mầm tạo ra đại bào tử thể; đại giao tử thể trưởng thành tạo ra túi cái, tạo ra noãn cầu. Trong mỗi noãn cầu là một đại bào tử nguyên phân tạo thành đại bào tử thể. Mỗi túi cái trong đại bào tử thể tạo một noãn. Toàn bộ thế hệ thể giao tử nằm trọn bên trong sự bảo vệ của cơ thể thể bào tử, trừ hạt phấn được phóng thích ra ngoài để phát tán (những hạt phấn, mang danh là thể giao tử nhưng chỉ gồm ba tế bào, nằm trong thành tế bào của tiểu bào tử.
- Hợp tử đực nguyên phân thành tiểu thể bào tử có khả năng tạo tiểu bào tử nang khi trưởng thành. Tiểu bào tử nang sinh ra tiểu bào tử qua hình thức giảm phân.
Chu kì sống của các nhóm thực vật khác nhau
[sửa | sửa mã nguồn]Sự luân phiên thế hệ diễn ra ở gần như tất cả tảo lục và tảo đỏ đa bào, kể cả tảo nước ngọt như Caldophora hay tảo nước mặn như Ulva. Thường thì các thế hệ đồng hình và sống độc lập. Phức tạp hơn, một số loài tảo đỏ có hệ thống luân phiên ba pha, gồm một pha giao tử thể và hai pha bào tử thể tách biệt. Xem thêm chi tiết ở Tảo đỏ: Sinh sản.
Tất cả thực vật trên cạn đều có sự luân phiên thế hệ dị hình, trong đó thể bào tử và thể giao tử có cấu trúc cơ thể khác nhau rõ rệt.
- Tất cả loài rêu, bao gồm rêu tản, rêu sừng và rêu thật sự đều có thế hệ thể giao tử trội. Xét một loài rêu đồng chu thể giao tử: túi đực và túi cái phát triển trên cây trưởng thành (thể giao tử); khi có nước, tinh trùng có hai đuôi từ túi đực bơi đến túi cái và bắt đầu thụ tinh, tạo ra một thẻ bào tử lưỡng bội. Bào tử này phát triển ngay bên trong túi cái, cơ thể nó chỉ bao gồm một cuống dài có một nang ở trên, sự giảm phân diễn ra trong nang đó tạo thành bào tử đơn bội. Hầu hết các loài rêu phát tán bào tử nhờ gió, nhưng có loài Splachnum sphaericum thì thụ phấn nhờ côn trùng. Xem thêm chi tiết ở Ngành Rêu tản: Vòng đời, Ngành Rêu sừng: Vòng đời, Ngành Rêu thật sự: Vòng đời.
-
 Chu kì sống của rêu tản
Chu kì sống của rêu tản -
 Chu kì sống của rêu thực sự
Chu kì sống của rêu thực sự -
 Chu kì sống của rêu sừng
Chu kì sống của rêu sừng



- Ở dương xỉ và các loài liên quan, bao gồm dương xỉ đuôi ngựa và thạch tùng, thể trội là thể bào tử. Bào tử phát triển trong các ổ bào tử nằm dưới các lá lược và được phát tán bằng gió (một số trường hợp bằng nước). Trong điều kiện thích hợp, bào tử sẽ nảy mầm thành một cơ thể gọi là nguyên tản (thể giao tử). Nguyên tản có hình dạng khác cây dương xỉ (thể bào tử), nên ta nói các loài này là loài dị hình. Nguyên tản không sống lâu, nhưng chúng thực hiện sinh sản hữu tính để tạo ra hợp tử lưỡng bội, hợp tử này phát triển ra khỏi nguyên tản để trở thành thể bào tử. Xem thêm chi tiết ở Ngành Dương xỉ: Vòng đời.
-
 Chu kì sống của dương xỉ
Chu kì sống của dương xỉ -

Nguyên tản (thể giao tử) của loài Dicksonia sp. -

Một thể bào tử lưỡng bội của loài Dicksonia antarctica. -

Mặt dưới của lá lược của loài Dicksonia antarctica, có thể thấy các ổ bào tử - cấu trúc sản xuất bào tử.




- Ở thực vật có hạt, thể trội cũng là thể bào tử; nhưng thể giao tử càng thoái hóa hơn về cả kích thước lẫn cấu tạo, nên có sự khác biệt về hình thái rất lớn. Toàn bộ thế hệ thể giao tử, ngoại trừ hạt phấn, đều được chứa bên trong thể bào tử. Vòng đời của một loài hạt kín biệt chu, cây liễu, đã được phân tích ở phần trên. Vòng đời của các loài hạt trần thì cũng tương tự. Nhưng ở thực vật hạt kín có một hiện tượng đặc biệt gọi là "thụ tinh kép". Hai tinh trùng từ hạt phấn (tiểu giao tử thể), thay vì chỉ một tinh trùng, thâm nhập vào túi cái của đại giao tử thể. Một tinh trùng dung hợp với trứng tạo thành hợp tử, một tinh trùng dung hợp với hai nhân của thể giao tử để tạo thành nội nhũ - một cấu trúc có chứng năng dinh dưỡng cho phôi. Xem thêm chi tiết tại Thụ tinh kép.
Sự tiến hoá của pha lưỡng bội trội
[sửa | sửa mã nguồn]Người ta cho rằng nền tảng của pha lưỡng bội trội là ở thể lưỡng bội (2n), mỗi gen đều có hai bản sao (alen), nên nếu một trong hai alen là alen xấu, đột biến hay gây hại cho cây, chúng sẽ có thể bị lấn át bởi alen còn lại.[28][29] Nói cách khác, nếu hệ gen của một trong hai cơ thể cha mẹ có alen xấu, chúng có thể được bù đắp bởi hệ gen của cơ thể cha mẹ còn lại (sự bổ trợ gen).
Khi thể lưỡng bội trở thành thể trội, sự bổ trợ gen giúp hệ gen và dung lượng thông tin trong hệ gen tăng kích thước mà không cần cải thiện độ chính xác của sự nhân đôi DNA. Đây là một lợi thế lớn vì nó cho phép nhiều cơ chế thích nghi mới xuất hiện. Tuy nhiên, thuyết này đang bị thách thức khi có nhiều bằng chứng cho thấy chọn lọc tự nhiên có hiệu quả cao hơn ở pha đơn bội, ở cả rêu và thực vật hạt kín.[30]
Người ta cho rằng nền tảng của việc thể lưỡng bội trở thành thể trội là vì sự lưỡng bội cho phép alen xấu bị lấn át và không thể hiện ra (sự bổ trợ gen).[28][29] Vì thế, nếu hệ gen của một trong hai cá thể cha mẹ có alen xấu, những thiếu sót này có thể được bù đắp bởi hệ gen của cơ thể cha mẹ còn lại (cơ thể này cũng có những alen xấu ở những gen khác).
-
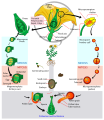 Chu kì sống của thực vật hạt kín
Chu kì sống của thực vật hạt kín -
 Hạt phấn (thể giao tử) dính trên đầu nhị của hoa Tulip
Hạt phấn (thể giao tử) dính trên đầu nhị của hoa Tulip -
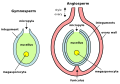 Noãn (đại giao tử thế) của thực vật hạt kín (bên trái) và thực vật hạt trần (bên phải)
Noãn (đại giao tử thế) của thực vật hạt kín (bên trái) và thực vật hạt trần (bên phải) -
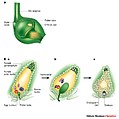 Thụ tinh kép ở thực vật
Thụ tinh kép ở thực vật




Chu trình tương tự ở các nhóm sinh vật khác
[sửa | sửa mã nguồn]Rhizaria
[sửa | sửa mã nguồn]
Một số sinh vật thuộc nhóm Rhizaria (không phải thực vật) cũng thực hiện sự luân phiên thế hệ. Hầu hết các loài trùng lỗ trải qua quá trình luân phiên thế hệ dị hình giữa thể hữu tính (gamont đơn bội) và thể vô tính (agamont lưỡng bội). Thể hữu tính đơn bội đơn bào thường có kích thước lớn hơn nhiều thể lưỡng bội.
Nấm
[sửa | sửa mã nguồn]Các hệ sợi nấm thường là đơn bội. Khi các hệ sợi nấm thuộc các loại giao cấu khác nhau tiếp hợp, chúng tạo ra hai tế bào đa nhân dạng cầu, nối nhau qua một "cầu giao cấu". Nhân tế bào di chuyển từ sợi nấm này sang sợi nấm kia, tạo thành một tế bào dị hạch (có nhân khác nhau). Quá trình này được gọi là tiếp hợp tế bào chất (hoặc tiếp hợp nguyên sinh chất, plasmogamy).
Tiếp đó, chúng sẽ thực hiện tiếp hợp nhân - quá trình thực sự tạo ra tế bào lưỡng bội. Quá trình này có thể bị hoãn lại cho đến khi túi bào tử được phát triển hoàn toàn. Tiếp hợp nhân tạo ra một hợp tử lưỡng bội, cũng là một thể bào tử sắp đi vào giảm phân để hình thành bào tử đơn bội. Khi bào tử này nảy mầm, chúng phát triển thành một hệ sợi nấm.
Nấm nhầy
[sửa | sửa mã nguồn]Vòng đời của nấm nhầy cũng không khác nấm nhiều. Bào tử đơn bội nảy mầm tạo thành các bào tử động hoặc các amip nhầy. Chúng thực hiện tiếp hợp nguyên sinh chất, rồi tiếp hợp nhân để tảo thành hợp tử lưỡng bội. Hợp tử này phát triển thành một khối hợp bào, khối hợp bào trưởng thành tạo ra một hay nhiều thể quá (tùy theo loài) chứa các bào tử đơn bội.
Động vật
[sửa | sửa mã nguồn]Sự luôn phiên giữa một thế hệ đơn bội và một thệ hệ lưỡng bội chưa bao giờ xuất hiện ở động vật.[31] Một số động vật có sự luân phiên giữa các pha sinh sản vô tính và hữu tính (dị giao), nhưng cả hai pha đều là lưỡng bội. Sự luân phiên này đôi lúc được gọi là "luân phiên thế hệ",[32] nhưng thực chất hai thứ này khác biệt. Ở một số động vật khác, bao gồm ong, kiến và các động vật khác thuộc bộ Cánh màng, con đực là đơn bội và con cái là lưỡng bội, nhưng đây không phải là sự luân phiên thế hệ vì chúng diễn ra cũng lúc ở mọi thế hệ, chứ không phải ở các thế hệ tách biệt.
Xem thêm
[sửa | sửa mã nguồn]Ghi chú và chú thích
[sửa | sửa mã nguồn]- ^ “alternation of generations | Definition & Examples”. Encyclopedia Britannica (bằng tiếng Anh). Truy cập ngày 25 tháng 2 năm 2021.
- ^ Thomas, R.J.; Stanton, D.S.; Longendorfer, D.H. & Farr, M.E. (1978), “Physiological evaluation of the nutritional autonomy of a hornwort sporophyte”, Botanical Gazette, 139 (3): 306–311, doi:10.1086/337006, S2CID 84413961
- ^ Glime, J.M. (2007), Bryophyte Ecology: Vol. 1 Physiological Ecology (PDF), Michigan Technological University and the International Association of Bryologists, truy cập ngày 4 tháng 3 năm 2013
- ^ Taylor, Kerp & Hass 2005
- ^ "“Plant Science 4 U”. Truy cập ngày 5 tháng 7 năm 2016.
- ^ Kerp, H.; Trewin, N.H. & Hass, H. (2003), “New gametophytes from the Lower Devonian Rhynie Chert”, Transactions of the Royal Society of Edinburgh: Earth Sciences, 94 (4): 411–428, doi:10.1017/S026359330000078X, S2CID 128629425
- ^ a b Bateman & Dimichele 1994, tr. 403
- ^ a b Stewart & Rothwell 1993
- ^ a b c d Haig, David (2008), “Homologous versus antithetic alternation of generations and the origin of sporophytes” (PDF), The Botanical Review, 74 (3): 395–418, doi:10.1007/s12229-008-9012-x, S2CID 207403936, truy cập ngày 17 tháng 8 năm 2014
- ^ Svedelius, Nils (1927), “Alternation of Generations in Relation to Reduction Division”, Botanical Gazette, 83 (4): 362–384, doi:10.1086/333745, JSTOR 2470766, S2CID 84406292
- ^ Hofmeister, W. (1851), Vergleichende Untersuchungen der Keimung, Entfaltung und Fruchtbildildiung höherer Kryptogamen (Moose, Farne, Equisetaceen, Rhizocarpeen und Lycopodiaceen) und der Samenbildung der Coniferen (bằng tiếng Đức), Leipzig: F. Hofmeister, truy cập ngày 17 tháng 8 năm 2014. Translated as Currey, Frederick (1862), On the germination, development, and fructification of the higher Cryptogamia, and on the fructification of the Coniferæ, London: Robert Hardwicke, truy cập ngày 17 tháng 8 năm 2014
- ^ Feldmann, J. & Feldmann, G. (1942), “Recherches sur les Bonnemaisoniacées et leur alternance de generations” (PDF), Ann. Sci. Natl. Bot., Series 11 (bằng tiếng Pháp), 3: 75–175, Bản gốc (PDF) lưu trữ ngày 19 tháng 8 năm 2014, truy cập ngày 7 tháng 10 năm 2013, p. 157
- ^ a b Feldmann, J. (1972), “Les problèmes actuels de l'alternance de génerations chez les Algues”, Bulletin de la Société Botanique de France (bằng tiếng Pháp), 119: 7–38, doi:10.1080/00378941.1972.10839073
- ^ Schopfer, P.; Mohr, H. (1995). “Physiology of Development”. Plant physiology. Berlin: Springer. tr. 288–291. ISBN 978-3-540-58016-4.
- ^ Unless otherwise indicated, the material in the whole of this section is based on Foster & Gifford 1974, Sporne 1974a and Sporne 1974b.
- ^ Guiry & Guiry 2008
- ^ Bateman & Dimichele 1994, tr. 347
- ^ a b Shyam 1980
- ^ Watson 1981, tr. 2
- ^ Kirby 2001
- ^ Watson 1981, tr. 33
- ^ Bell & Hemsley 2000, tr. 104
- ^ Watson 1981, tr. 425–6
- ^ Watson 1981, tr. 287–8
- ^ Sporne 1974a, tr. 17–21.
- ^ a b Bateman & Dimichele 1994, tr. 350–1
- ^ Chisholm, Hugh biên tập (1911). . Encyclopædia Britannica. 28 (ấn bản thứ 11). Cambridge University Press. tr. 688–689.
- ^ a b Bernstein, H.; Byers, G.S. & Michod, R.E. (1981), “Evolution of sexual reproduction: Importance of DNA repair, complementation, and variation”, The American Naturalist, 117 (4): 537–549, doi:10.1086/283734, S2CID 84568130
- ^ a b Michod, R.E. & Gayley, T.W. (1992), “Masking of mutations and the evolution of sex”, The American Naturalist, 139 (4): 706–734, doi:10.1086/285354, S2CID 85407883
- ^ Szövényi, Péter; Ricca, Mariana; Hock, Zsófia; Shaw, Jonathan A.; Shimizu, Kentaro K. & Wagner, Andreas (2013), “Selection is no more efficient in haploid than in diploid life stages of an angiosperm and a moss”, Molecular Biology and Evolution, 30 (8): 1929–39, doi:10.1093/molbev/mst095, PMID 23686659
- ^ Barnes và đồng nghiệp 2001, tr. 321
- ^ Scott 1996, tr. 35
Tham khảo
[sửa | sửa mã nguồn]- Barnes, R.S.K.; Calow, P.; Olive, P.J.W.; Golding, D.W. & Spicer, J.I. (2001), The Invertebrates: a synthesis, Oxford; Malden, MA: Blackwell, ISBN 978-0-632-04761-1
- Bateman, R.M. & Dimichele, W.A. (1994), “Heterospory – the most iterative key innovation in the evolutionary history of the plant kingdom” (PDF), Biological Reviews of the Cambridge Philosophical Society, 69 (3): 345–417, doi:10.1111/j.1469-185x.1994.tb01276.x, S2CID 29709953, Bản gốc (PDF) lưu trữ ngày 15 tháng 4 năm 2012, truy cập ngày 30 tháng 12 năm 2010
- Bell, P.R. & Hemsley, A.R. (2000), Green Plants: their Origin and Diversity (ấn bản thứ 2), Cambridge, etc.: Cambridge University Press, ISBN 978-0-521-64109-8
- Foster, A.S. & Gifford, E.M. (1974), Comparative Morphology of Vascular Plants (ấn bản thứ 2), San Francisco: W.H. Freeman, ISBN 978-0-7167-0712-7
- Guiry, M.D.; Guiry, G.M. (2008), “Cladophora”, AlgaeBase, World-wide electronic publication, National University of Ireland, Galway, truy cập ngày 21 tháng 7 năm 2011
- Kirby, A. (2001), Ulva, the sea lettuce, Monterey Bay Aquarium Research Institute, Bản gốc lưu trữ ngày 16 tháng 5 năm 2011, truy cập ngày 1 tháng 1 năm 2011
- Scott, Thomas (1996), Concise Encyclopedia Biology, Berlin: Walter de Gruyter, ISBN 978-3-11-010661-9
- Shyam, R. (1980), “On the life-cycle, cytology and taxonomy of Cladophora callicoma from India”, American Journal of Botany, 67 (5): 619–24, doi:10.2307/2442655, JSTOR 2442655
- Sporne, K.R. (1974a), The Morphology of Angiosperms, London: Hutchinson, ISBN 978-0-09-120611-6
- Sporne, K.R. (1974b), The Morphology of Gymnosperms (ấn bản thứ 2), London: Hutchinson, ISBN 978-0-09-077152-3
- Stewart, W.N. & Rothwell, G.W. (1993), Paleobotany and the Evolution of Plants (ấn bản thứ 2), Cambridge, UK: Cambridge University Press, ISBN 978-0-521-38294-6
- Watson, E.V. (1981), British Mosses and Liverworts (ấn bản thứ 3), Cambridge, UK: Cambridge University Press, ISBN 978-0-521-28536-0
- Taylor, T.N.; Kerp, H. & Hass, H. (2005), “Life history biology of early land plants: Deciphering the gametophyte phase”, Proceedings of the National Academy of Sciences of the United States of America, 102 (16): 5892–5897, doi:10.1073/pnas.0501985102, PMC 556298, PMID 15809414