

Espécie em anel
Este artigo ou secção contém uma lista de referências no fim do texto, mas as suas fontes não são claras porque não são citadas no corpo do artigo, o que compromete a confiabilidade das informações. (Fevereiro de 2019) |


Em biologia, uma espécie em anel é uma série conectada de populações vizinhas, cada uma das quais cruza com populações relacionadas próximas, mas para as quais existem pelo menos duas populações "finais" na série, que são muito distantemente relacionadas ao cruzamento, embora haja é um fluxo gênico potencial entre cada população "ligada".[1] Essas populações não reprodutoras, embora geneticamente conectadas, podem coexistir na mesma região ( simpatria ) fechando assim um "anel". O termo alemão Rassenkreis , significando um anel de populações, também é usado.
As espécies em anel representam a especiação e foram citadas como evidência da evolução . Eles ilustram o que acontece ao longo do tempo à medida que as populações divergem geneticamente, especificamente porque representam, em populações vivas, o que normalmente acontece ao longo do tempo entre populações de ancestrais falecidos há muito tempo e populações vivas, nas quais os intermediários se extinguiram . O biólogo evolucionista Richard Dawkins observa que as espécies em anel "estão apenas nos mostrando na dimensão espacial algo que sempre deve acontecer na dimensão do tempo".[2]
Formalmente, a questão é que a interfertilidade (capacidade de cruzar) não é uma relação transitiva ; se A cruza com B e B cruza com C, isso não significa que A cruza com C e, portanto, não define uma relação de equivalência . Uma espécie em anel é uma espécie com um contra-exemplo para a transitividade do cruzamento.[3] No entanto, não está claro se algum dos exemplos de espécies em anel citados por cientistas realmente permite o fluxo gênico de ponta a ponta, com muitos sendo debatidos e contestados.
História
[editar | editar código-fonte]-
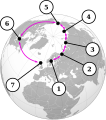 As gaivotas Larus cruzam em um anel ao redor do Ártico. 1: L. fuscus, 2: população siberiana de L. fuscus, 3: EU. heuglini, 4: L. vegae birulai, 5: L. vegae, 6: L. smithsonianus, 7: L. argentatus
As gaivotas Larus cruzam em um anel ao redor do Ártico. 1: L. fuscus, 2: população siberiana de L. fuscus, 3: EU. heuglini, 4: L. vegae birulai, 5: L. vegae, 6: L. smithsonianus, 7: L. argentatus -
 Gaivota-prateada ( Larus argentatus ) (frente) e gaivota-preta ( Larus fuscus ) (atrás) na Noruega: dois fenótipos com diferenças claras
Gaivota-prateada ( Larus argentatus ) (frente) e gaivota-preta ( Larus fuscus ) (atrás) na Noruega: dois fenótipos com diferenças claras


A espécie clássica em anel é a gaivota Larus . Em 1925, Jonathan Dwight descobriu que o gênero formava uma cadeia de variedades ao redor do Círculo Polar Ártico. No entanto, surgiram dúvidas se isso representa uma espécie real em anel.[4] Em 1938, Claud Buchanan Ticehurst argumentou que a toutinegra esverdeada se espalhou do Nepal ao redor do planalto tibetano, enquanto se adaptava a cada novo ambiente, encontrando-se novamente na Sibéria, onde as extremidades não mais se cruzam.[5] Essas e outras descobertas levaram Mayr a formular pela primeira vez uma teoria sobre as espécies em anel em seu estudo de 1942, Systematics and the Origin of Species . Também na década de 1940, Robert C. Stebbins descreveu as salamandras Ensatina ao redor do Vale Central da Califórnia como uma espécie em anel;[6][7] mas, novamente, alguns autores como Jerry Coyne consideram essa classificação incorreta.[8] Finalmente, em 2012, o primeiro exemplo de uma espécie em anel em plantas foi encontrado em um spurge, formando um anel em torno do Mar do Caribe.[9]
Especiação
[editar | editar código-fonte]O biólogo Ernst Mayr defendeu o conceito de espécies em anel, alegando que ele demonstrava inequivocamente o processo de especiação.[10] Uma espécie em anel é um modelo alternativo à especiação alopátrica, "ilustrando como novas espécies podem surgir por meio de 'sobreposição circular', sem interrupção do fluxo gênico através das populações intervenientes ..." [11] No entanto, Jerry Coyne e H. Allen Orr apontam que os anéis espécies modelam mais de perto a especiação parapátrica .[8]
As espécies em anel freqüentemente atraem o interesse de biólogos evolucionistas, sistematistas e pesquisadores da especiação, levando a idéias instigantes e confusão sobre sua definição.[12] Estudiosos contemporâneos reconhecem que os exemplos na natureza se mostraram raros devido a vários fatores, como limitações no delineamento taxonômico [13] ou "zelo taxonômico" [10] - explicado pelo fato de que os taxonomistas classificam os organismos em "espécies", enquanto as espécies em anel frequentemente não pode se enquadrar nesta definição.[12] Outras razões, como interrupção do fluxo gênico por "divergência de vicariato" e populações fragmentadas devido à instabilidade climática, também foram citadas.[10]
As espécies em anel também apresentam um caso interessante do problema das espécies para aqueles que buscam dividir o mundo vivo em espécies distintas. Tudo o que distingue uma espécie em anel de duas espécies separadas é a existência de populações conectadas; se um número suficiente de populações conectadas dentro do anel perecer para cortar a conexão reprodutiva, então as populações distais das espécies em anel serão reconhecidas como duas espécies distintas. O problema é quantificar o anel inteiro como uma única espécie (apesar do fato de que nem todos os indivíduos se cruzam) ou classificar cada população como uma espécie distinta (apesar do fato de cruzar com seus vizinhos próximos). As espécies em anel ilustram que os limites das espécies surgem gradualmente e freqüentemente existem em um continuum.[10]
Exemplos
[editar | editar código-fonte]

Muitos exemplos foram documentados na natureza. Existe debate sobre grande parte da pesquisa, com alguns autores citando evidências inteiramente contra sua existência.[8][14] Os exemplos a seguir fornecem evidências de que - apesar do número limitado de exemplos concretos e idealizados na natureza - continuums de espécies existem e podem ser encontrados em sistemas biológicos.[10] Isso geralmente é caracterizado por classificações em nível de subespécies, como clines, ecótipos, complexos e variedades . Muitos exemplos foram contestados por pesquisadores, e igualmente "muitos dos casos [propostos] receberam muito pouca atenção dos pesquisadores, tornando difícil avaliar se eles exibem as características de espécies anelares ideais." [12]
A lista a seguir dá exemplos de espécies em anel encontradas na natureza. Alguns dos exemplos, como o complexo de gaivota Larus, a toutinegra esverdeada da Ásia e as salamandras Ensatina da América, foram contestados.[14][15][16][17]
- Acanthiza pusilla e A. ewingii [18]
- Acacia karroo [12][19][20]
- Cotovias Alauda ( Alauda arvensis, A. japonica e A. gulgula ) [12]
- Alophoixus [10][21]
- Aulostomus (peixe-trombeta) [22]
- Camarhynchus psittacula e C. pauper [12]
- Complexo de espécies de Chaerephon pumilus [23]
- Salamandras Ensatina [24][25]
- Euphorbia tithymaloides é um grupo dentro da família spurge que se reproduziu e evoluiu em um anel através da América Central e do Caribe, encontrando-se nas Ilhas Virgens, onde parecem ser morfológica e ecologicamente distintos.[9]
- Chapim-real [12] (no entanto, alguns estudos contestam este exemplo [26][27] )
- A toutinegra esverdeada ( Phylloscopus trochiloides ) forma um anel de espécies, em torno do Himalaia .[28][29][30][31] Acredita-se que ele tenha se espalhado do Nepal ao redor do planalto tibetano inóspito, para se reunir na Sibéria, onde o plumbeitarsus e o viridanus pareciam não mais se reproduzir mutuamente.
- Hoplitis producta [12]
- Rato doméstico [12]
- Junonia coenia e J. genoveva / J. evarete [12]
- Lalage leucopygialis, L. nigra e L. sueurii [12]
- As gaivotas larus formam um "anel" circumpolar ao redor do Pólo Norte . A gaivota arenosa europeia ( L. argentatus argenteus ), que vive principalmente na Grã-Bretanha e na Irlanda, pode hibridizar com a gaivota arenque americana ( L. smithsonianus ), (que vive na América do Norte ), que também pode hibridar com o Vega ou Siberian A gaivota arenosa ( L. vegae ), cuja subespécie ocidental, a gaivota de Birula ( L. vegae birulai ), pode hibridizar com a gaivota de Heuglin ( L. heuglini ), que por sua vez pode hibridizar com a gaivota-preta siberiana ( L. fuscus ). Todos os quatro vivem no norte da Sibéria . O último é o representante oriental das gaivotas-de-costas-negras menores no noroeste da Europa, incluindo a Grã-Bretanha. As gaivotas-de-dorso-preto menores e as gaivotas-arenque são suficientemente diferentes que normalmente não hibridizam; assim, o grupo de gaivotas forma um continuum, exceto onde as duas linhagens se encontram na Europa. No entanto, um estudo genético de 2004 intitulado "O complexo de gaivota de arenque não é uma espécie em anel" mostrou que este exemplo é muito mais complicado do que o apresentado aqui (Liebers et al., 2004):[32] este exemplo fala apenas do complexo de espécies desde a gaivota arenosa clássica até a gaivota-de-costas-negras. Existem vários outros exemplos taxonomicamente obscuros que pertencem ao mesmo complexo de espécies, como gaivota de patas amarelas ( L. michahellis ), gaivota glaucosa ( L. hyperboreus ) e gaivota do Cáspio ( L. cachinnans ).
- Pelophylax nigromaculatus e P. porosus / P. porosus brevipodus [12] (os nomes e classificação dessas espécies mudaram desde a publicação, sugerindo uma espécie em anel)
- Pernis ptilorhynchus e P. celebensis [12]
- Perognathus amplus e P. longimembris [12]
- Peromyscus maniculatus [33]
- Phellinus [34]
- Complexo de Platycercus elegans ( rosela carmesim ) [35][36]
- Drosophila paulistorum [37]
- Phylloscopus collybita e P. sindianus [12]
- Phylloscopus ( toutinegras do salgueiro ) [38][39]
- Powelliphanta [40]
- Rhymogona silvatica e R. cervina [12] (os nomes e classificação dessas espécies mudaram desde a publicação, sugerindo uma espécie em anel)
- Melospiza melodia, um pardal cantor, forma um anel ao redor da Sierra Nevada da Califórnia [41] com as subespécies heermanni e fallax encontrando-se nas proximidades da passagem de San Gorgonio .
- Todiramphus chloris e T. cinnamominus [12]
Ver também
[editar | editar código-fonte]- ↑ Irwin, Darren E.; Irwin, J. H.; Price, T. D. (2001). «Ring species as bridges between microevolution and speciation» (PDF). Genetica. 112–113: 223–43. PMID 11838767. doi:10.1023/A:1013319217703
- ↑ Dawkins, Richard (2004). The Ancestor's Tale. Houghton Mifflin. [S.l.: s.n.] ISBN 0-618-00583-8
- ↑ Brown, Rob. «'Same Species' vs. 'Interfertile: concise wording can avoid confusion when discussing evolution»
- ↑ Liebers, D.; De Knijff, P.; Helbig, A. J. (2004). «The herring gull complex is not a ring species». Proceedings of the Royal Society B: Biological Sciences. 271: 893–901. PMC 1691675
 . PMID 15255043. doi:10.1098/rspb.2004.2679
. PMID 15255043. doi:10.1098/rspb.2004.2679
- ↑ Irwin, Darren. «The greenish warbler ring species»
- ↑ «A closer look at a classic ring species: The work of Tom Devitt». Understanding Evolution
- ↑ This species ring forms the subject of "The Salamander's tale" in Richard Dawkins' The Ancestor's Tale, 2004.
- ↑ a b c Coyne, Jerry A.; Orr, H. Allen (2004). Speciation. Sinauer Associates. [S.l.: s.n.] pp. 102–105. ISBN 0-87893-091-4
- ↑ a b Ivalú Cacho, N.; Baum, David A. (2012). «The Caribbean slipper spurge Euphorbia tithymaloides: the first example of a ring species in plants». Proc. R. Soc. B. 279: 3377–3383. PMC 3396892. PMID 22696529. doi:10.1098/rspb.2012.0498
- ↑ a b c d e f Pereira, Ricardo J.; Wake, David B. (2015). «Ring species as demonstrations of the continuum of species formation». Molecular Ecology. 24: 5312–5314. PMID 26509692. doi:10.1111/mec.13412
- ↑ Helbig, A. J. (2005). «A ring of species». Heredity. 95: 113–114. PMID 15999143. doi:10.1038/sj.hdy.6800679
- ↑ a b c d e f g h i j k l m n o p q Irwin, Darren E.; Irwin, Jessica H.; Price, Trevor D. (2001). «Ring species as bridges between microevolution and speciation». Genetica. 112–113: 223–243. PMID 11838767. doi:10.1023/A:1013319217703
- ↑ Irwin, Darren E. (2012). «A novel approach for finding ring species: look for barriers rather than rings». BMC Biology. 10. 21 páginas. PMC 3299606. PMID 22410355. doi:10.1186/1741-7007-10-21
- ↑ a b Coyne, Jerry (16 julho de 2014). «There are no ring species». Why Evolution is True
- ↑ Alcaide, M.; Scordato, E. S. C.; et al. (2014). «Genomic divergence in a ring species complex». Nature. 511: 83–85. Bibcode:2014Natur.511...83A. PMID 24870239. doi:10.1038/nature13285
|hdl-access=requer|hdl=(ajuda) - ↑ Liebers, Dorit; Knijff, Peter de; Helbig, Andreas J. (2004). «The herring gull complex is not a ring species». Proc Biol Sci. 271: 893–901. PMC 1691675. PMID 15255043. doi:10.1098/rspb.2004.2679
- ↑ Highton, R. (1998). «Is Ensatina eschscholtzii a ring species?». Herpetologica. 54: 254–278. JSTOR 3893431
- ↑ Simpson, K.; Day, N.; Trusler, P. (1999), Birds of Australia 6 ed. , Princeton University Press
- ↑ Brain, P. (1989), «Genetic races in a ring species, Acacia-Karroo», South African Journal of Science, 85 (3): 181–185
- ↑ Ward, David (2011), «Population differentiation in a purported ring species, Acacia karroo (Mimosoideae)», Biological Journal of the Linnean Society, 104 (4): 748–755, doi:10.1111/j.1095-8312.2011.01757.x
- ↑ Jérôme Fuchs; et al. (2015), «The complex phylogeography of the Indo-Malayan Alophoixus bulbuls with the description of a putative new ring species complex», Molecular Ecology, 24 (21): 5460–5474, PMID 26224534, doi:10.1111/mec.13337
- ↑ Bowen, B. W.; et al. (2001), «Phytogeography of the Trumpetfishes (Aulostomus): Ring Species Complex on a Global Scale», Evolution, 55 (5): 1029–1039, PMID 11430639, doi:10.1111/j.0014-3820.2001.tb00619.x
- ↑ Naidoo, Theshnie; et al. (2016), «Partial support for the classical ring species hypothesis in the Chaerephon pumilus species complex (Chiroptera: Molossidae) from southeastern Africa and western Indian Ocean islands», Mammalia, 80 (6), doi:10.1515/mammalia-2015-0062
- ↑ Moritz, Craig; et al. (1992), «Evolutionary Relationships Within the Ensatina Eschscholtzii Complex Confirm the Ring Species Interpretation», Systematic Biology, 41 (3): 273–291, doi:10.1093/sysbio/41.3.273
- ↑ Moritz, Craig; Schneider, C.J. (1992). «Evolutionary relationships within the Ensatina eschscholtzii complex confirm the ring species interpretation». Systematic Biology. 41: 273–291. JSTOR 2992567. doi:10.2307/2992567
- ↑ Päckert, Martin; et al. (2005), «The great tit (Parus major) – a misclassified ring species», Biological Journal of the Linnean Society, 86 (2): 153–174, doi:10.1111/j.1095-8312.2005.00529.x
- ↑ Kvist, Laura; et al. (2003), «Evolution and genetic structure of the great tit (Parus major) complex», Proc. R. Soc. Lond. B, 270 (1523): 1447–1454, PMC 1691391, PMID 12965008, doi:10.1098/rspb.2002.2321
- ↑ Alström, Per (2006). «Species concepts and their application: insights from the genera Seicercus and Phylloscopus». Acta Zoologica Sinica. 52: 429–434. Cópia arquivada em 2 de março de 2014
- ↑ Irwin, Darren E.; et al. (2005), «Speciation by Distance in a Ring Species», Science, 307 (5708): 414–416, Bibcode:2005Sci...307..414I, PMID 15662011, doi:10.1126/science.1105201
- ↑ Irwin, Darren E. (2000). «Song variation in an avian ring species». Evolution. 54: 998–1010. PMID 10937272. doi:10.1111/j.0014-3820.2000.tb00099.x
- ↑ Alcaide, Miguel; et al. (2014). «Genomic divergence in a ring species complex». Nature. 511: 83–85. Bibcode:2014Natur.511...83A. PMID 24870239. doi:10.1038/nature13285
|hdl-access=requer|hdl=(ajuda) - ↑ Liebers, Dorit; de Knijff, Peter; Helbig, Andreas J. (2004). «The herring gull complex is not a ring species». Proceedings of the Royal Society B. 271: 893–901. PMC 1691675. PMID 15255043. doi:10.1098/rspb.2004.2679 Electronic Appendix[ligação inativa]
- ↑ Caire, William; Zimmerman, Earl G. (1975). «Chromosomal and Morphological Variation and Circular Overlap in the Deer Mouse, Peromyscus Maniculatus, in Texas and Oklahoma». Systematic Zoology. 24: 89–95. doi:10.1093/sysbio/24.1.89
- ↑ Parmasto, Erast (2007). «Phellinus laevigatus s. l. (Hymenochaetales): a ring species». Folia Cryptogamica Estonica. 43: 39–49
- ↑ Joseph, Leo; et al. (2008). «Where and when does a ring start and end? Testing the ring-species hypothesis in a species complex of Australian parrots». Proc. R. Soc. B. 275: 2431–2440. PMC 2603204. PMID 18664434. doi:10.1098/rspb.2008.0765
- ↑ Eastwood, Justin R.; et al. (2014). «Phylogenetic analysis of beak and feather disease virus across a host ring-species complex». PNAS. 111: 14153–14158. Bibcode:2014PNAS..11114153E. PMC 4191811. PMID 25225394. doi:10.1073/pnas.1403255111
- ↑ Dobzhansky, T.; Spassky, B. (1959). «Drosophila paulistorum, a cluster of species in statu nascendi». Proc. Natl. Acad. Sci. 45: 419–428. Bibcode:1959PNAS...45..419D. PMC 222578. PMID 16590403. doi:10.1073/pnas.45.3.419
- ↑ Bensch, Staffan; et al. (2009), «Genetic, morphological, and feather isotope variation of migratory willow warblers show gradual divergence in a ring», Molecular Ecology, 18 (14): 3087–3096, PMID 19457197, doi:10.1111/j.1365-294X.2009.04210.x
- ↑ Irwin, Darren E. Irwin (2009), «Incipient ring speciation revealed by a migratory divide», Molecular Ecology, 18 (14): 2923–2925, PMID 19457189, doi:10.1111/j.1365-294X.2009.04211.x
- ↑ Climo, F. M. (1978), «The Powelliphanta gilliesi - traversi - hochstetteri - rossiana - lignaria - superba ring species (Mollusca: Pulmonata)», New Zealand Journal of Zoology, 5 (2): 289–294, doi:10.1080/03014223.1978.10428318
- ↑ Patten, Michael A.; Pruett, Christin L. (2009). «The Song Sparrow,Melospiza melodia,as a ring species: Patterns of geographic variation, a revision of subspecies, and implications for speciation». Systematics and Biodiversity. 7: 33–62. doi:10.1017/S1477200008002867