JP2011505796A - vaccine - Google Patents
vaccine Download PDFInfo
- Publication number
- JP2011505796A JP2011505796A JP2010536451A JP2010536451A JP2011505796A JP 2011505796 A JP2011505796 A JP 2011505796A JP 2010536451 A JP2010536451 A JP 2010536451A JP 2010536451 A JP2010536451 A JP 2010536451A JP 2011505796 A JP2011505796 A JP 2011505796A
- Authority
- JP
- Japan
- Prior art keywords
- protein
- specific
- ade2
- vaccine
- seq
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 229960005486 vaccine Drugs 0.000 title claims abstract description 37
- 108090000623 proteins and genes Proteins 0.000 claims abstract description 68
- 102000004169 proteins and genes Human genes 0.000 claims abstract description 62
- 239000000203 mixture Substances 0.000 claims abstract description 61
- 239000013598 vector Substances 0.000 claims abstract description 44
- 239000002671 adjuvant Substances 0.000 claims abstract description 38
- 239000000427 antigen Substances 0.000 claims abstract description 36
- 108091007433 antigens Proteins 0.000 claims abstract description 36
- 102000036639 antigens Human genes 0.000 claims abstract description 36
- 201000004792 malaria Diseases 0.000 claims abstract description 33
- 239000013603 viral vector Substances 0.000 claims abstract description 30
- 241000223960 Plasmodium falciparum Species 0.000 claims abstract description 25
- 238000004519 manufacturing process Methods 0.000 claims abstract description 16
- 229930182490 saponin Natural products 0.000 claims abstract description 13
- 150000007949 saponins Chemical class 0.000 claims abstract description 13
- 230000002265 prevention Effects 0.000 claims abstract description 11
- 239000001397 quillaja saponaria molina bark Substances 0.000 claims abstract description 11
- 239000012634 fragment Substances 0.000 claims abstract description 8
- 230000002950 deficient Effects 0.000 claims abstract description 6
- 230000010076 replication Effects 0.000 claims abstract description 6
- 241000990167 unclassified Simian adenoviruses Species 0.000 claims abstract 5
- 238000009472 formulation Methods 0.000 claims description 41
- 230000000638 stimulation Effects 0.000 claims description 17
- 238000000034 method Methods 0.000 claims description 15
- 239000002502 liposome Substances 0.000 claims description 8
- 239000000546 pharmaceutical excipient Substances 0.000 claims description 8
- 239000003814 drug Substances 0.000 claims description 5
- 239000007764 o/w emulsion Substances 0.000 claims description 5
- 238000002360 preparation method Methods 0.000 claims description 4
- 230000037452 priming Effects 0.000 claims 1
- 230000001902 propagating effect Effects 0.000 claims 1
- 208000015181 infectious disease Diseases 0.000 abstract description 7
- 230000005867 T cell response Effects 0.000 description 109
- 238000011748 CB6F1 mouse Methods 0.000 description 67
- 230000004044 response Effects 0.000 description 48
- 235000018102 proteins Nutrition 0.000 description 45
- 108090000695 Cytokines Proteins 0.000 description 43
- 102000004127 Cytokines Human genes 0.000 description 43
- 241000699670 Mus sp. Species 0.000 description 30
- 210000004899 c-terminal region Anatomy 0.000 description 24
- 210000004027 cell Anatomy 0.000 description 23
- 230000005875 antibody response Effects 0.000 description 21
- 210000001744 T-lymphocyte Anatomy 0.000 description 17
- 230000003053 immunization Effects 0.000 description 17
- 238000002649 immunization Methods 0.000 description 17
- 108010002350 Interleukin-2 Proteins 0.000 description 16
- CTMZLDSMFCVUNX-VMIOUTBZSA-N cytidylyl-(3'->5')-guanosine Chemical compound O=C1N=C(N)C=CN1[C@H]1[C@H](O)[C@H](OP(O)(=O)OC[C@@H]2[C@H]([C@@H](O)[C@@H](O2)N2C3=C(C(N=C(N)N3)=O)N=C2)O)[C@@H](CO)O1 CTMZLDSMFCVUNX-VMIOUTBZSA-N 0.000 description 16
- 108090000765 processed proteins & peptides Proteins 0.000 description 16
- 241001135569 Human adenovirus 5 Species 0.000 description 15
- 101150106931 IFNG gene Proteins 0.000 description 15
- 108091034117 Oligonucleotide Proteins 0.000 description 13
- 239000002773 nucleotide Substances 0.000 description 12
- 125000003729 nucleotide group Chemical group 0.000 description 12
- 239000013612 plasmid Substances 0.000 description 12
- 210000003046 sporozoite Anatomy 0.000 description 11
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 10
- 239000000556 agonist Substances 0.000 description 10
- 239000002245 particle Substances 0.000 description 10
- 102000004196 processed proteins & peptides Human genes 0.000 description 10
- 235000017709 saponins Nutrition 0.000 description 10
- 125000003275 alpha amino acid group Chemical group 0.000 description 8
- 235000001014 amino acid Nutrition 0.000 description 8
- 150000001413 amino acids Chemical class 0.000 description 8
- 230000005847 immunogenicity Effects 0.000 description 8
- 230000001225 therapeutic effect Effects 0.000 description 8
- 239000013642 negative control Substances 0.000 description 7
- 241000701161 unidentified adenovirus Species 0.000 description 7
- 102100037850 Interferon gamma Human genes 0.000 description 6
- 108010074328 Interferon-gamma Proteins 0.000 description 6
- 229930182558 Sterol Natural products 0.000 description 6
- 102100039360 Toll-like receptor 4 Human genes 0.000 description 6
- 238000000684 flow cytometry Methods 0.000 description 6
- 239000001963 growth medium Substances 0.000 description 6
- 238000000338 in vitro Methods 0.000 description 6
- 150000002632 lipids Chemical class 0.000 description 6
- 229940035032 monophosphoryl lipid a Drugs 0.000 description 6
- 210000004897 n-terminal region Anatomy 0.000 description 6
- 210000005259 peripheral blood Anatomy 0.000 description 6
- 239000011886 peripheral blood Substances 0.000 description 6
- 150000003432 sterols Chemical class 0.000 description 6
- 235000003702 sterols Nutrition 0.000 description 6
- 241000255925 Diptera Species 0.000 description 5
- 238000002965 ELISA Methods 0.000 description 5
- 101000669447 Homo sapiens Toll-like receptor 4 Proteins 0.000 description 5
- 230000000694 effects Effects 0.000 description 5
- 238000002474 experimental method Methods 0.000 description 5
- 230000003308 immunostimulating effect Effects 0.000 description 5
- 230000001404 mediated effect Effects 0.000 description 5
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 4
- 241000282412 Homo Species 0.000 description 4
- UZQJVUCHXGYFLQ-AYDHOLPZSA-N [(2s,3r,4s,5r,6r)-4-[(2s,3r,4s,5r,6r)-4-[(2r,3r,4s,5r,6r)-4-[(2s,3r,4s,5r,6r)-3,5-dihydroxy-6-(hydroxymethyl)-4-[(2s,3r,4s,5s,6r)-3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxyoxan-2-yl]oxy-3,5-dihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-3,5-dihydroxy-6-(hy Chemical compound O([C@H]1[C@H](O)[C@@H](CO)O[C@H]([C@@H]1O)O[C@H]1[C@H](O)[C@@H](CO)O[C@H]([C@@H]1O)O[C@H]1CC[C@]2(C)[C@H]3CC=C4[C@@]([C@@]3(CC[C@H]2[C@@]1(C=O)C)C)(C)CC(O)[C@]1(CCC(CC14)(C)C)C(=O)O[C@H]1[C@@H]([C@@H](O[C@H]2[C@@H]([C@@H](O[C@H]3[C@@H]([C@@H](O[C@H]4[C@@H]([C@@H](O[C@H]5[C@@H]([C@@H](O)[C@H](O)[C@@H](CO)O5)O)[C@H](O)[C@@H](CO)O4)O)[C@H](O)[C@@H](CO)O3)O)[C@H](O)[C@@H](CO)O2)O)[C@H](O)[C@@H](CO)O1)O)[C@@H]1O[C@H](CO)[C@@H](O)[C@H](O)[C@H]1O UZQJVUCHXGYFLQ-AYDHOLPZSA-N 0.000 description 4
- 230000028993 immune response Effects 0.000 description 4
- 108091033319 polynucleotide Proteins 0.000 description 4
- 102000040430 polynucleotide Human genes 0.000 description 4
- 239000002157 polynucleotide Substances 0.000 description 4
- 229920001184 polypeptide Polymers 0.000 description 4
- 241000282693 Cercopithecidae Species 0.000 description 3
- 241000598171 Human adenovirus sp. Species 0.000 description 3
- 108090001030 Lipoproteins Proteins 0.000 description 3
- 102000004895 Lipoproteins Human genes 0.000 description 3
- 108091028043 Nucleic acid sequence Proteins 0.000 description 3
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 3
- 101710137302 Surface antigen S Proteins 0.000 description 3
- 102000002689 Toll-like receptor Human genes 0.000 description 3
- 108020000411 Toll-like receptor Proteins 0.000 description 3
- 241000235013 Yarrowia Species 0.000 description 3
- 239000000969 carrier Substances 0.000 description 3
- 239000000839 emulsion Substances 0.000 description 3
- 210000003743 erythrocyte Anatomy 0.000 description 3
- 230000036039 immunity Effects 0.000 description 3
- 229960001438 immunostimulant agent Drugs 0.000 description 3
- 239000003022 immunostimulating agent Substances 0.000 description 3
- 238000001727 in vivo Methods 0.000 description 3
- 238000002347 injection Methods 0.000 description 3
- 239000007924 injection Substances 0.000 description 3
- 239000008194 pharmaceutical composition Substances 0.000 description 3
- RYYWUUFWQRZTIU-UHFFFAOYSA-K thiophosphate Chemical compound [O-]P([O-])([O-])=S RYYWUUFWQRZTIU-UHFFFAOYSA-K 0.000 description 3
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 3
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 2
- 108020004705 Codon Proteins 0.000 description 2
- 241000701022 Cytomegalovirus Species 0.000 description 2
- 108020004414 DNA Proteins 0.000 description 2
- 241001092142 Molina Species 0.000 description 2
- 108700026244 Open Reading Frames Proteins 0.000 description 2
- 241000235648 Pichia Species 0.000 description 2
- 241000224016 Plasmodium Species 0.000 description 2
- 206010057190 Respiratory tract infections Diseases 0.000 description 2
- 235000014680 Saccharomyces cerevisiae Nutrition 0.000 description 2
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 2
- 102100024333 Toll-like receptor 2 Human genes 0.000 description 2
- -1 alkyl glucosaminide phosphates Chemical class 0.000 description 2
- 230000015572 biosynthetic process Effects 0.000 description 2
- HVYWMOMLDIMFJA-DPAQBDIFSA-N cholesterol Chemical compound C1C=C2C[C@@H](O)CC[C@]2(C)[C@@H]2[C@@H]1[C@@H]1CC[C@H]([C@H](C)CCCC(C)C)[C@@]1(C)CC2 HVYWMOMLDIMFJA-DPAQBDIFSA-N 0.000 description 2
- 210000001151 cytotoxic T lymphocyte Anatomy 0.000 description 2
- FOTKYAAJKYLFFN-UHFFFAOYSA-N decane-1,10-diol Chemical compound OCCCCCCCCCCO FOTKYAAJKYLFFN-UHFFFAOYSA-N 0.000 description 2
- 201000010099 disease Diseases 0.000 description 2
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 2
- 239000004615 ingredient Substances 0.000 description 2
- 238000011081 inoculation Methods 0.000 description 2
- 239000003446 ligand Substances 0.000 description 2
- GZQKNULLWNGMCW-PWQABINMSA-N lipid A (E. coli) Chemical class O1[C@H](CO)[C@@H](OP(O)(O)=O)[C@H](OC(=O)C[C@@H](CCCCCCCCCCC)OC(=O)CCCCCCCCCCCCC)[C@@H](NC(=O)C[C@@H](CCCCCCCCCCC)OC(=O)CCCCCCCCCCC)[C@@H]1OC[C@@H]1[C@@H](O)[C@H](OC(=O)C[C@H](O)CCCCCCCCCCC)[C@@H](NC(=O)C[C@H](O)CCCCCCCCCCC)[C@@H](OP(O)(O)=O)O1 GZQKNULLWNGMCW-PWQABINMSA-N 0.000 description 2
- 210000004185 liver Anatomy 0.000 description 2
- 229940124735 malaria vaccine Drugs 0.000 description 2
- 210000003936 merozoite Anatomy 0.000 description 2
- 239000000693 micelle Substances 0.000 description 2
- 238000006386 neutralization reaction Methods 0.000 description 2
- 150000007523 nucleic acids Chemical group 0.000 description 2
- WTJKGGKOPKCXLL-RRHRGVEJSA-N phosphatidylcholine Chemical compound CCCCCCCCCCCCCCCC(=O)OC[C@H](COP([O-])(=O)OCC[N+](C)(C)C)OC(=O)CCCCCCCC=CCCCCCCCC WTJKGGKOPKCXLL-RRHRGVEJSA-N 0.000 description 2
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 2
- 108091008146 restriction endonucleases Proteins 0.000 description 2
- 150000003839 salts Chemical class 0.000 description 2
- 239000011780 sodium chloride Substances 0.000 description 2
- 241000894007 species Species 0.000 description 2
- 229940044616 toll-like receptor 7 agonist Drugs 0.000 description 2
- 229940044655 toll-like receptor 9 agonist Drugs 0.000 description 2
- 230000014616 translation Effects 0.000 description 2
- 210000002845 virion Anatomy 0.000 description 2
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 1
- SNKAWJBJQDLSFF-NVKMUCNASA-N 1,2-dioleoyl-sn-glycero-3-phosphocholine Chemical compound CCCCCCCC\C=C/CCCCCCCC(=O)OC[C@H](COP([O-])(=O)OCC[N+](C)(C)C)OC(=O)CCCCCCC\C=C/CCCCCCCC SNKAWJBJQDLSFF-NVKMUCNASA-N 0.000 description 1
- IJFVSSZAOYLHEE-UHFFFAOYSA-N 2,3-di(dodecanoyloxy)propyl 2-(trimethylazaniumyl)ethyl phosphate Chemical compound CCCCCCCCCCCC(=O)OCC(COP([O-])(=O)OCC[N+](C)(C)C)OC(=O)CCCCCCCCCCC IJFVSSZAOYLHEE-UHFFFAOYSA-N 0.000 description 1
- CXURGFRDGROIKG-UHFFFAOYSA-N 3,3-bis(chloromethyl)oxetane Chemical group ClCC1(CCl)COC1 CXURGFRDGROIKG-UHFFFAOYSA-N 0.000 description 1
- RHKWIGHJGOEUSM-UHFFFAOYSA-N 3h-imidazo[4,5-h]quinoline Chemical class C1=CN=C2C(N=CN3)=C3C=CC2=C1 RHKWIGHJGOEUSM-UHFFFAOYSA-N 0.000 description 1
- SLXKOJJOQWFEFD-UHFFFAOYSA-N 6-aminohexanoic acid Chemical compound NCCCCCC(O)=O SLXKOJJOQWFEFD-UHFFFAOYSA-N 0.000 description 1
- 101150116940 AGPS gene Proteins 0.000 description 1
- 208000010370 Adenoviridae Infections Diseases 0.000 description 1
- 206010060931 Adenovirus infection Diseases 0.000 description 1
- 108090000565 Capsid Proteins Proteins 0.000 description 1
- 102100023321 Ceruloplasmin Human genes 0.000 description 1
- 101710117490 Circumsporozoite protein Proteins 0.000 description 1
- 241000759568 Corixa Species 0.000 description 1
- 102100031262 Deleted in malignant brain tumors 1 protein Human genes 0.000 description 1
- 206010012735 Diarrhoea Diseases 0.000 description 1
- 102000002322 Egg Proteins Human genes 0.000 description 1
- 108010000912 Egg Proteins Proteins 0.000 description 1
- 101710145505 Fiber protein Proteins 0.000 description 1
- 101100024440 Globodera rostochiensis MSP-3 gene Proteins 0.000 description 1
- 241000238631 Hexapoda Species 0.000 description 1
- 101000844721 Homo sapiens Deleted in malignant brain tumors 1 protein Proteins 0.000 description 1
- 101001130441 Homo sapiens Ras-related protein Rap-2a Proteins 0.000 description 1
- 101000831567 Homo sapiens Toll-like receptor 2 Proteins 0.000 description 1
- 101000831496 Homo sapiens Toll-like receptor 3 Proteins 0.000 description 1
- 101000669402 Homo sapiens Toll-like receptor 7 Proteins 0.000 description 1
- 101000800483 Homo sapiens Toll-like receptor 8 Proteins 0.000 description 1
- 241000235649 Kluyveromyces Species 0.000 description 1
- 241001138401 Kluyveromyces lactis Species 0.000 description 1
- 241000235058 Komagataella pastoris Species 0.000 description 1
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 1
- 102000018697 Membrane Proteins Human genes 0.000 description 1
- 108010052285 Membrane Proteins Proteins 0.000 description 1
- 108010057081 Merozoite Surface Protein 1 Proteins 0.000 description 1
- 241001465754 Metazoa Species 0.000 description 1
- NTIZESTWPVYFNL-UHFFFAOYSA-N Methyl isobutyl ketone Chemical group CC(C)CC(C)=O NTIZESTWPVYFNL-UHFFFAOYSA-N 0.000 description 1
- MSFSPUZXLOGKHJ-UHFFFAOYSA-N Muraminsaeure Natural products OC(=O)C(C)OC1C(N)C(O)OC(CO)C1O MSFSPUZXLOGKHJ-UHFFFAOYSA-N 0.000 description 1
- 241000699666 Mus <mouse, genus> Species 0.000 description 1
- HCUVEUVIUAJXRB-UHFFFAOYSA-N OC1=C(C=C(CNC(CCCC=2SC=CC=2)=O)C=C1)OC Chemical compound OC1=C(C=C(CNC(CCCC=2SC=CC=2)=O)C=C1)OC HCUVEUVIUAJXRB-UHFFFAOYSA-N 0.000 description 1
- 241000320412 Ogataea angusta Species 0.000 description 1
- 241000283973 Oryctolagus cuniculus Species 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 108010013639 Peptidoglycan Proteins 0.000 description 1
- 240000009188 Phyllostachys vivax Species 0.000 description 1
- 101000983333 Plasmodium falciparum (isolate NF54) 25 kDa ookinete surface antigen Proteins 0.000 description 1
- 241001505483 Plasmodium falciparum 3D7 Species 0.000 description 1
- 241000223980 Plasmodium falciparum NF54 Species 0.000 description 1
- 241001454523 Quillaja saponaria Species 0.000 description 1
- 235000009001 Quillaja saponaria Nutrition 0.000 description 1
- 102100022851 Rab5 GDP/GTP exchange factor Human genes 0.000 description 1
- 102100031420 Ras-related protein Rap-2a Human genes 0.000 description 1
- 101710203837 Replication-associated protein Proteins 0.000 description 1
- 241000235070 Saccharomyces Species 0.000 description 1
- 241000219287 Saponaria Species 0.000 description 1
- 241000235346 Schizosaccharomyces Species 0.000 description 1
- 241000235347 Schizosaccharomyces pombe Species 0.000 description 1
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 1
- 108700005078 Synthetic Genes Proteins 0.000 description 1
- 230000024932 T cell mediated immunity Effects 0.000 description 1
- 210000000447 Th1 cell Anatomy 0.000 description 1
- 108010060804 Toll-Like Receptor 4 Proteins 0.000 description 1
- 108010060888 Toll-like receptor 2 Proteins 0.000 description 1
- 102000008230 Toll-like receptor 3 Human genes 0.000 description 1
- 102100024324 Toll-like receptor 3 Human genes 0.000 description 1
- 108010060885 Toll-like receptor 3 Proteins 0.000 description 1
- 102100039390 Toll-like receptor 7 Human genes 0.000 description 1
- 102100033110 Toll-like receptor 8 Human genes 0.000 description 1
- 229940122089 Toll-like receptor 8 agonist Drugs 0.000 description 1
- 208000035896 Twin-reversed arterial perfusion sequence Diseases 0.000 description 1
- 108020000999 Viral RNA Proteins 0.000 description 1
- 241000700605 Viruses Species 0.000 description 1
- 239000002253 acid Substances 0.000 description 1
- 230000001154 acute effect Effects 0.000 description 1
- 238000007792 addition Methods 0.000 description 1
- 208000011589 adenoviridae infectious disease Diseases 0.000 description 1
- 235000004279 alanine Nutrition 0.000 description 1
- 101150078331 ama-1 gene Proteins 0.000 description 1
- 239000005557 antagonist Substances 0.000 description 1
- 239000003833 bile salt Substances 0.000 description 1
- 229940093761 bile salts Drugs 0.000 description 1
- 230000003115 biocidal effect Effects 0.000 description 1
- 210000004369 blood Anatomy 0.000 description 1
- 239000008280 blood Substances 0.000 description 1
- 125000003178 carboxy group Chemical group [H]OC(*)=O 0.000 description 1
- 235000012000 cholesterol Nutrition 0.000 description 1
- 239000013599 cloning vector Substances 0.000 description 1
- 125000004122 cyclic group Chemical group 0.000 description 1
- OPTASPLRGRRNAP-UHFFFAOYSA-N cytosine Chemical class NC=1C=CNC(=O)N=1 OPTASPLRGRRNAP-UHFFFAOYSA-N 0.000 description 1
- 230000034994 death Effects 0.000 description 1
- 231100000517 death Toxicity 0.000 description 1
- 238000012217 deletion Methods 0.000 description 1
- 230000037430 deletion Effects 0.000 description 1
- 238000011161 development Methods 0.000 description 1
- 230000018109 developmental process Effects 0.000 description 1
- 230000004069 differentiation Effects 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- 239000003085 diluting agent Substances 0.000 description 1
- NAGJZTKCGNOGPW-UHFFFAOYSA-K dioxido-sulfanylidene-sulfido-$l^{5}-phosphane Chemical compound [O-]P([O-])([S-])=S NAGJZTKCGNOGPW-UHFFFAOYSA-K 0.000 description 1
- NAGJZTKCGNOGPW-UHFFFAOYSA-N dithiophosphoric acid Chemical class OP(O)(S)=S NAGJZTKCGNOGPW-UHFFFAOYSA-N 0.000 description 1
- 210000003162 effector t lymphocyte Anatomy 0.000 description 1
- 235000013345 egg yolk Nutrition 0.000 description 1
- 210000002969 egg yolk Anatomy 0.000 description 1
- 238000005538 encapsulation Methods 0.000 description 1
- ZINJLDJMHCUBIP-UHFFFAOYSA-N ethametsulfuron-methyl Chemical compound CCOC1=NC(NC)=NC(NC(=O)NS(=O)(=O)C=2C(=CC=CC=2)C(=O)OC)=N1 ZINJLDJMHCUBIP-UHFFFAOYSA-N 0.000 description 1
- 238000013401 experimental design Methods 0.000 description 1
- 230000002349 favourable effect Effects 0.000 description 1
- 108020001507 fusion proteins Proteins 0.000 description 1
- 102000037865 fusion proteins Human genes 0.000 description 1
- 102000018146 globin Human genes 0.000 description 1
- 108060003196 globin Proteins 0.000 description 1
- 230000013595 glycosylation Effects 0.000 description 1
- 238000006206 glycosylation reaction Methods 0.000 description 1
- UYTPUPDQBNUYGX-UHFFFAOYSA-N guanine Chemical class O=C1NC(N)=NC2=C1N=CN2 UYTPUPDQBNUYGX-UHFFFAOYSA-N 0.000 description 1
- 230000005802 health problem Effects 0.000 description 1
- 208000002672 hepatitis B Diseases 0.000 description 1
- 210000003494 hepatocyte Anatomy 0.000 description 1
- 238000004128 high performance liquid chromatography Methods 0.000 description 1
- 229960002751 imiquimod Drugs 0.000 description 1
- DOUYETYNHWVLEO-UHFFFAOYSA-N imiquimod Chemical compound C1=CC=CC2=C3N(CC(C)C)C=NC3=C(N)N=C21 DOUYETYNHWVLEO-UHFFFAOYSA-N 0.000 description 1
- 230000002163 immunogen Effects 0.000 description 1
- 230000009851 immunogenic response Effects 0.000 description 1
- 230000006698 induction Effects 0.000 description 1
- 230000003834 intracellular effect Effects 0.000 description 1
- 238000002955 isolation Methods 0.000 description 1
- 239000003550 marker Substances 0.000 description 1
- 239000002184 metal Substances 0.000 description 1
- 230000003278 mimic effect Effects 0.000 description 1
- 230000004899 motility Effects 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- 230000003071 parasitic effect Effects 0.000 description 1
- 230000007918 pathogenicity Effects 0.000 description 1
- 235000021317 phosphate Nutrition 0.000 description 1
- 150000004713 phosphodiesters Chemical class 0.000 description 1
- 150000003904 phospholipids Chemical class 0.000 description 1
- 230000001766 physiological effect Effects 0.000 description 1
- 239000013600 plasmid vector Substances 0.000 description 1
- 230000008488 polyadenylation Effects 0.000 description 1
- 239000013641 positive control Substances 0.000 description 1
- 230000000644 propagated effect Effects 0.000 description 1
- 230000000069 prophylactic effect Effects 0.000 description 1
- 230000001105 regulatory effect Effects 0.000 description 1
- 229950010550 resiquimod Drugs 0.000 description 1
- BXNMTOQRYBFHNZ-UHFFFAOYSA-N resiquimod Chemical compound C1=CC=CC2=C(N(C(COCC)=N3)CC(C)(C)O)C3=C(N)N=C21 BXNMTOQRYBFHNZ-UHFFFAOYSA-N 0.000 description 1
- 210000003079 salivary gland Anatomy 0.000 description 1
- 229920006395 saturated elastomer Polymers 0.000 description 1
- 230000001568 sexual effect Effects 0.000 description 1
- 238000010561 standard procedure Methods 0.000 description 1
- 230000000153 supplemental effect Effects 0.000 description 1
- 208000037369 susceptibility to malaria Diseases 0.000 description 1
- 238000003786 synthesis reaction Methods 0.000 description 1
- 239000003970 toll like receptor agonist Substances 0.000 description 1
- 231100000419 toxicity Toxicity 0.000 description 1
- 230000001988 toxicity Effects 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- 230000003612 virological effect Effects 0.000 description 1
Images









































































Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/002—Protozoa antigens
- A61K39/015—Hemosporidia antigens, e.g. Plasmodium antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P33/00—Antiparasitic agents
- A61P33/02—Antiprotozoals, e.g. for leishmaniasis, trichomoniasis, toxoplasmosis
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P33/00—Antiparasitic agents
- A61P33/02—Antiprotozoals, e.g. for leishmaniasis, trichomoniasis, toxoplasmosis
- A61P33/06—Antimalarials
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/86—Viral vectors
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N7/00—Viruses; Bacteriophages; Compositions thereof; Preparation or purification thereof
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/525—Virus
- A61K2039/5256—Virus expressing foreign proteins
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/53—DNA (RNA) vaccination
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55511—Organic adjuvants
- A61K2039/55572—Lipopolysaccharides; Lipid A; Monophosphoryl lipid A
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/555—Medicinal preparations containing antigens or antibodies characterised by a specific combination antigen/adjuvant
- A61K2039/55511—Organic adjuvants
- A61K2039/55577—Saponins; Quil A; QS21; ISCOMS
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2710/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA dsDNA viruses
- C12N2710/00011—Details
- C12N2710/10011—Adenoviridae
- C12N2710/10311—Mastadenovirus, e.g. human or simian adenoviruses
- C12N2710/10341—Use of virus, viral particle or viral elements as a vector
- C12N2710/10343—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- Y—GENERAL TAGGING OF NEW TECHNOLOGICAL DEVELOPMENTS; GENERAL TAGGING OF CROSS-SECTIONAL TECHNOLOGIES SPANNING OVER SEVERAL SECTIONS OF THE IPC; TECHNICAL SUBJECTS COVERED BY FORMER USPC CROSS-REFERENCE ART COLLECTIONS [XRACs] AND DIGESTS
- Y02—TECHNOLOGIES OR APPLICATIONS FOR MITIGATION OR ADAPTATION AGAINST CLIMATE CHANGE
- Y02A—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE
- Y02A50/00—TECHNOLOGIES FOR ADAPTATION TO CLIMATE CHANGE in human health protection, e.g. against extreme weather
- Y02A50/30—Against vector-borne diseases, e.g. mosquito-borne, fly-borne, tick-borne or waterborne diseases whose impact is exacerbated by climate change
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Genetics & Genomics (AREA)
- Organic Chemistry (AREA)
- Tropical Medicine & Parasitology (AREA)
- Veterinary Medicine (AREA)
- Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Medicinal Chemistry (AREA)
- Zoology (AREA)
- Pharmacology & Pharmacy (AREA)
- Animal Behavior & Ethology (AREA)
- Public Health (AREA)
- General Engineering & Computer Science (AREA)
- Microbiology (AREA)
- Biotechnology (AREA)
- Biomedical Technology (AREA)
- Virology (AREA)
- General Chemical & Material Sciences (AREA)
- Biochemistry (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Immunology (AREA)
- Molecular Biology (AREA)
- Plant Pathology (AREA)
- Mycology (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Epidemiology (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Peptides Or Proteins (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicinal Preparation (AREA)
Abstract
本発明は、Plasmodium falciparumからのCSタンパク質またはその断片を含むタンパク質(例えば、配列番号1または配列番号3に示されているもの)をコードする複製欠損型サルアデノウイルスベクターC7に関する。本発明はまた、該ウイルスベクターの製造方法、およびマラリア感染の治療/予防における該ウイルスベクターの使用に関する。該ウイルスベクターを含む組成物、ワクチンおよびキットも記載する。1つの態様においては、本発明は合成C7ウイルスベクターを使用する。本発明のC7ウイルスベクターは、場合によってはアジュバント、例えば3D-MPLおよび/またはサポニン(例えばQS21)を含むアジュバントの存在下、マラリア抗原、例えばRTS,Sと共に共投与または共配合されうる。
【選択図】図16The present invention relates to a replication-deficient simian adenovirus vector C7 that encodes a protein comprising CS protein from Plasmodium falciparum or a fragment thereof (eg, those shown in SEQ ID NO: 1 or SEQ ID NO: 3). The present invention also relates to a method for producing the viral vector and the use of the viral vector in the treatment / prevention of malaria infection. Also described are compositions, vaccines and kits comprising the viral vectors. In one embodiment, the present invention uses a synthetic C7 viral vector. The C7 viral vectors of the invention can be co-administered or co-formulated with a malaria antigen such as RTS, S, optionally in the presence of an adjuvant such as 3D-MPL and / or an saponin (eg QS21).
[Selection] Figure 16
Description
本発明は、サル由来アデノウイルスベクター、特に、Plasmodium falciparumのサーカムスポロゾイト(circumsporozoite)タンパク質に由来する新規マラリア抗原をコードするサル由来アデノウイルスベクターに関する。本発明は更に、該ウイルスベクターの製造方法、およびマラリア感染の治療/予防におけるその使用に関する。 The present invention relates to monkey-derived adenovirus vectors, and particularly to monkey-derived adenovirus vectors encoding novel malaria antigens derived from the plasma sporozoite protein of Plasmodium falciparum. The invention further relates to a method for producing said viral vector and its use in the treatment / prevention of malaria infection.
マラリアは世界的に重大な健康問題の1つであり、毎年200万人〜400万人以上がこの疾患で死亡している。 Malaria is one of the world's major health problems, with between 2 and 4 million people dying from the disease each year.
この疾患の最も急性な形態の1つは、マラリアに起因する死亡のほとんどの原因となる寄生原虫Plasmodium falciparum(P. falciparum)により引き起こされる。 One of the most acute forms of the disease is caused by the parasitic protozoan Plasmodium falciparum (P. falciparum), which is responsible for most of the deaths caused by malaria.
P. falciparumの生活環は複雑であり、完了のためにはヒトおよび蚊の2つの宿主を要する。ヒトへの感染は、感染蚊が刺すことにより血流中にスポロゾイトが接種されることにより開始される。スポロゾイトは肝臓へ移動し、そこで肝細胞に感染し、ここでスポロゾイトは赤血球外細胞内段階を経てメロゾイト段階へと分化し、これは赤血球(RBC)に感染して無性血液段階における循環的複製を開始する。この循環は、RBC内で多数のメロゾイトが有性段階配偶子細胞へ分化することにより完了し、有性段階配偶子細胞は蚊により摂取され、蚊の体内で該細胞は中腸内での一連の段階を経て発育してスポロゾイトとなり、これは唾液腺に移行する。 The life cycle of P. falciparum is complex and requires two hosts, human and mosquito to complete. Infection to humans is initiated by inoculation of sporozoites into the bloodstream by the bite of infected mosquitoes. Sporozoites migrate to the liver, where they infect hepatocytes, where the sporozoites differentiate into the merozoite stage via the extra-erythrocytic intracellular stage, which infects the red blood cells (RBC) and circulates in the asexual blood stage. To start. This circulation is completed by the differentiation of a large number of merozoites into sexual stage gamete cells in the RBC, which are ingested by mosquitoes, and in the mosquito body, the cells are in a series in the midgut. It develops through this stage and becomes a sporozoite, which moves to the salivary glands.
P. falciparumのスポロゾイト段階はマラリアワクチンの潜在的標的と目されている。不活化(照射)スポロゾイトは実験的ヒトマラリアに対する保護を誘導することが示されている(Am. J, Trop. Med. Hyg 24: 297-402, 1975)。しかし、実際に且つ事業計画的に、照射スポロゾイトを使用するこの方法に基づいて一般集団におけるマラリアに対するワクチンを製造することは不可能である。 The sporozoite stage of P. falciparum is seen as a potential target for malaria vaccines. Inactivated (irradiated) sporozoites have been shown to induce protection against experimental human malaria (Am. J, Trop. Med. Hyg 24: 297-402, 1975). However, in practice and in business planning, it is impossible to produce a vaccine against malaria in the general population based on this method using irradiated sporozoites.
スポロゾイトの主要表面タンパク質はサーカムスポロゾイトタンパク質(CSタンパク質)として公知である。それは蚊による接種の最初の部位から循環内(循環内でそれは肝臓へ移行する)へのスポロゾイトの移行中のスポロゾイトの運動性および侵襲性に関与していると考えられている。 The main surface protein of sporozoites is known as circumsporozoite protein (CS protein). It is thought to be involved in the motility and invasiveness of the sporozoites during the transfer of the sporozoites from the initial site of mosquito inoculation into the circulation (in which it moves to the liver).
Plasmodia種のCSタンパク質は、非反復性アミノ(N末端)およびカルボキシ(C末端)断片に隣接した中央反復ドメイン(反復領域)により特徴づけられる。 Plasmodia species CS proteins are characterized by a central repeat domain (repeat region) adjacent to non-repeating amino (N-terminal) and carboxy (C-terminal) fragments.
現在のところ、臨床において最も進んだマラリアワクチンは、RTS,Sと称されるリポタンパク質粒子(ウイルス様粒子としても公知である)に基づくものである。この粒子は、B型肝炎由来のS抗原のN末端に融合したP. falciparum(株NF54[3D7])のCSタンパク質のアミノ酸207-395に実質的に対応するP. falciparumのCSタンパク質の部分を含有する。該S抗原はプレS2の部分を含みうる。 At present, the most advanced malaria vaccine in the clinic is based on lipoprotein particles called RTS, S (also known as virus-like particles). This particle contains a portion of the CS protein of P. falciparum that substantially corresponds to amino acids 207-395 of the CS protein of P. falciparum (strain NF54 [3D7]) fused to the N-terminus of the S antigen from hepatitis B. contains. The S antigen may comprise a portion of pre-S2.
RTS,S粒子は、通常、強力なアジュバントと共に送達される。 RTS, S particles are usually delivered with a powerful adjuvant.
それにもかかわらず、組換えアデノウイルスベクターを利用するマラリアワクチンが提示されている。例えば、WO 2004/055187は、CSタンパク質をコードする特異的アデノ5(Ad5)およびアデノ35(Ad35)ベクター(共にヒトアデノウイルスに由来する)を含む或るウイルスベクターを記載している。 Nevertheless, malaria vaccines utilizing recombinant adenovirus vectors have been presented. For example, WO 2004/055187 describes certain viral vectors, including specific adeno5 (Ad5) and adeno35 (Ad35) vectors (both derived from human adenoviruses) that encode CS proteins.
病原性が異なる40を超える異なるヒトアデノウイルス血清型が存在する。例えば、Ad5は小児における軽度の呼吸器感染に関連しており、Ad4およびAd7は成人における呼吸器感染に関連していると考えられており、Ad40は乳児における下痢を引き起こすと考えられている。 There are over 40 different human adenovirus serotypes that differ in pathogenicity. For example, Ad5 is associated with mild respiratory infections in children, Ad4 and Ad7 are thought to be associated with respiratory infections in adults, and Ad40 is thought to cause diarrhea in infants.
アデノウイルス感染に対する免疫は感染後生涯にわたると考えられている。特にAd5およびAd35に対する既存免疫は、ヒトアデノウイルスに基づく治療用アデノウイルスベクターの中和をもたらしうると考えられている。これは該ベクターの治療的有効性を軽減しうる。なぜなら、該ベクターが細胞に進入し関連抗原をin vivoで生成するのが妨げられるからである。 Immunity against adenovirus infection is thought to last a lifetime after infection. In particular, it is believed that pre-existing immunity against Ad5 and Ad35 can result in neutralization of therapeutic adenoviral vectors based on human adenovirus. This can reduce the therapeutic effectiveness of the vector. This is because the vector is prevented from entering the cell and generating the relevant antigen in vivo.
本発明は、P. falciparumからのCSタンパク質またはその断片、例えば配列番号1または配列番号3に示すものを含むタンパク質をコードする複製欠損型サル(シミアン)アデノウイルスベクターC7(Pan7またはCV-33とも称される)を含む、マラリアの予防および/または治療のためのワクチンを提供することにより、既存免疫の問題を軽減すると考えられる。 The present invention relates to a replication-deficient monkey (simian) adenoviral vector C7 (also referred to as Pan7 or CV-33) encoding a protein comprising CS protein from P. falciparum or a fragment thereof, such as those shown in SEQ ID NO: 1 or SEQ ID NO: 3. Providing a vaccine for the prevention and / or treatment of malaria, including
配列の簡単な説明
配列番号1:P. falciparumのCSタンパク質に由来するタンパク質/抗原のアミノ酸配列(本明細書においてはAde2と称される)。
配列番号2:配列番号1のタンパク質をコードする核酸配列(本明細書においてはAde2遺伝子と称される)。
配列番号3:P. falciparumのCSタンパク質に由来するタンパク質/抗原の代替的アミノ酸配列(本明細書においてはAde1と称される)。
配列番号4:配列番号3のタンパク質をコードする核酸配列(本明細書においてはAde1遺伝子と称される)。
配列番号5:Chimp Adeno(チンプ・アデノ)7からのカプシドタンパク質配列(WO 03/046124の配列番号17)。
配列番号6:Chimp Adeno(チンプ・アデノ)7からのアミノ酸配列(WO 03/046124の配列番号20)。
配列番号7:P. falciparum CSタンパク質からのアミノ酸配列。
配列番号8:P. falciparum CSタンパク質からのアミノ酸配列。
配列番号9:P. falciparum CSタンパク質からのアミノ酸配列。
配列番号10:P. falciparum CSタンパク質からのアミノ酸配列。
配列番号11:CpG 1826のヌクレオチド配列。
配列番号12:CpG 1758のヌクレオチド配列。
配列番号13:CpGのヌクレオチド配列。
配列番号14:CpG 2006のヌクレオチド配列。
配列番号15:CpG 1668のヌクレオチド配列。
配列番号16:CpG 5456のヌクレオチド配列。
配列番号17:C7アデノウイルスベクター内にクローニングされた、Ade2発現カセットに代わる発現カセットのヌクレオチド配列を示す。
配列番号18:C7アデノウイルスベクター内にクローニングされた、Ade2発現カセットのヌクレオチド配列を示す。
配列番号19:合成組換えベクターC7-Ade2の完全ヌクレオチド配列を示す。
BRIEF DESCRIPTION OF THE SEQUENCES SEQ ID NO: 1: Protein / antigen amino acid sequence derived from the CS protein of P. falciparum (referred to herein as Ade2).
SEQ ID NO: 2: nucleic acid sequence encoding the protein of SEQ ID NO: 1 (referred to herein as the Ade2 gene).
SEQ ID NO: 3: Alternative amino acid sequence of a protein / antigen derived from the CS protein of P. falciparum (referred to herein as Ade1).
SEQ ID NO: 4: nucleic acid sequence encoding the protein of SEQ ID NO: 3 (referred to herein as the Ade1 gene).
SEQ ID NO: 5: Capsid protein sequence from Chimp Adeno 7 (SEQ ID NO: 17 of WO 03/046124).
SEQ ID NO: 6: Amino acid sequence from Chimp Adeno 7 (SEQ ID NO: 20 of WO 03/046124).
SEQ ID NO: 7: amino acid sequence from P. falciparum CS protein.
SEQ ID NO: 8: amino acid sequence from P. falciparum CS protein.
SEQ ID NO: 9: Amino acid sequence from P. falciparum CS protein.
SEQ ID NO: 10: amino acid sequence from P. falciparum CS protein.
SEQ ID NO: 11: nucleotide sequence of CpG 1826.
SEQ ID NO: 12: nucleotide sequence of CpG 1758.
SEQ ID NO: 13: Nucleotide sequence of CpG.
SEQ ID NO: 14: Nucleotide sequence of CpG 2006.
SEQ ID NO: 15: nucleotide sequence of CpG 1668.
SEQ ID NO: 16: nucleotide sequence of CpG 5456.
SEQ ID NO: 17: Shows the nucleotide sequence of an expression cassette that replaces the Ade2 expression cassette, cloned into a C7 adenoviral vector.
SEQ ID NO: 18: Shows the nucleotide sequence of the Ade2 expression cassette cloned into a C7 adenoviral vector.
SEQ ID NO: 19: This shows the complete nucleotide sequence of the synthetic recombinant vector C7-Ade2.
C7の配列および製造はWO 2003/046124に記載されている。WO 2003/046124の配列番号6(ペントン配列)、9(核酸配列)、10および11(ヘキソン配列)、ならびに12(ファイバータンパク質)を参照により本明細書に組み入れる。C7の寄託番号は[ATCC VR-593]である。 The sequence and production of C7 is described in WO 2003/046124. WO 2003/046124, SEQ ID NO: 6 (penton sequence), 9 (nucleic acid sequence), 10 and 11 (hexon sequence), and 12 (fiber protein) are incorporated herein by reference. The deposit number for C7 is [ATCC VR-593].
ベクターは幾つかのファミリーに分類されることが可能であり、所定のファミリー内のアデノウイルスベクターは類似した特徴を有しうるという仮定が存在するものの、任意の所定のアデノウイルスベクターの特徴および特性は特有のものであることが多い。 Vectors can be grouped into several families, and there are assumptions that adenoviral vectors within a given family may have similar characteristics, but the characteristics and characteristics of any given adenoviral vector Is often unique.
C7の使用は特に有利だと考えられる。なぜなら、それは、タンパク質コード化遺伝子が挿入された場合、同様にWO 2003/046124に記載されている例えばC6のような他の公知ベクターより見かけ上安定だからである。つまり、C7は、再構成される傾向がより低いと考えられる。もちろん、医薬品は、それらが販売されうる前に、十分に特徴づけられ安定かつ安全であると示される必要があるため、ワクチン中で使用されるいずれのアデノウイルスベクターも安定であることが非常に重要である。 The use of C7 is considered particularly advantageous. This is because when a protein-encoding gene is inserted, it is apparently more stable than other known vectors such as C6 described in WO 2003/046124. In other words, C7 is considered less prone to reconfiguration. Of course, it is very important that any adenoviral vector used in a vaccine is stable because pharmaceuticals need to be shown to be well-characterized, stable and safe before they can be sold. is important.
C7に対する既存免疫は非常に低いと考えられ、したがって、患者への最初の投与の後の該ウイルスベクターの中和の危険性は低い。 Existing immunity against C7 appears to be very low, so the risk of neutralization of the viral vector after the first administration to the patient is low.
さらに、ヒトへの投与および/またはin vivoでの好ましい免疫応答の生成にC7を特に適したものにしうる、C7の1以上の他の特性が存在すると考えられている。 Furthermore, it is believed that there are one or more other properties of C7 that can make it particularly suitable for administration to humans and / or generating a favorable immune response in vivo.
1つの態様においては、本発明は、ヒトへの投与に関する規制機関の承認を得るのに特に好適でありうる合成C7ウイルスベクターを使用する。 In one embodiment, the present invention uses a synthetic C7 viral vector that may be particularly suitable for obtaining regulatory approval for human administration.
1つの態様においては、CSタンパク質からのマラリア抗原成分の最後の12〜14アミノ酸が除去されている。 In one embodiment, the last 12-14 amino acids of the malaria antigen component from the CS protein have been removed.
1つの態様においては、該アデノウイルスベクターによりコードされるマラリア抗原は、潜在的グリコシル化部位を除去するよう改変されている。例えば、配列番号1の約379位に示すとおり、アミノ酸アラニンがセリンの代わりに使用されうる。 In one embodiment, the malaria antigen encoded by the adenoviral vector has been modified to remove a potential glycosylation site. For example, as shown at about position 379 of SEQ ID NO: 1, the amino acid alanine can be used in place of serine.
本発明の1つの態様においては、使用されるタンパク質/抗原は以下のアミノ酸を含む:
![]()
(これは、場合によっては、アミノ酸約81-99に位置する)。
In one embodiment of the invention, the protein / antigen used comprises the following amino acids:
![]()
(This is optionally located at about amino acids 81-99).
1つの態様においては、コードされるタンパク質/抗原はアミノ酸:
AIGL(配列番号8)
を例えばC末端に含む。
In one embodiment, the encoded protein / antigen is an amino acid:
AIGL (SEQ ID NO: 8)
For example at the C-terminus.
1つの態様においては、本発明は、以下のアミノ酸を含むタンパク質を使用する:
1つの態様においては、本発明は、以下のアミノ酸を含むタンパク質を使用する:
もう1つの態様においては、使用されるタンパク質/抗原は配列番号7および/または配列番号8および/または配列番号9の配列を含む。 In another embodiment, the protein / antigen used comprises the sequence of SEQ ID NO: 7 and / or SEQ ID NO: 8 and / or SEQ ID NO: 9.
もう1つの態様においては、使用されるタンパク質/抗原は配列番号7および/または配列番号8および/または配列番号10の配列を含む。 In another embodiment, the protein / antigen used comprises the sequence of SEQ ID NO: 7 and / or SEQ ID NO: 8 and / or SEQ ID NO: 10.
1つの態様においては、コードされるタンパク質/抗原は配列番号1または3である。 In one embodiment, the encoded protein / antigen is SEQ ID NO: 1 or 3.
配列番号1に示されるタンパク質配列は新規であり、本発明の1つの態様を構成する。 The protein sequence shown in SEQ ID NO: 1 is novel and constitutes one embodiment of the present invention.
配列番号1のタンパク質配列をコードするポリヌクレオチド、特に、配列番号2のポリヌクレオチド配列も、本発明の1つの態様を構成する。このポリヌクレオチド配列(配列番号2)はヒトにおける発現に関して既にコドン最適化されている。 The polynucleotide encoding the protein sequence of SEQ ID NO: 1, in particular the polynucleotide sequence of SEQ ID NO: 2 also constitutes one embodiment of the present invention. This polynucleotide sequence (SEQ ID NO: 2) has already been codon optimized for expression in humans.
場合によっては、配列番号1のタンパク質をコードするポリヌクレオチド配列はコドン最適化されうる。 In some cases, the polynucleotide sequence encoding the protein of SEQ ID NO: 1 may be codon optimized.
本発明はまた、配列番号1の新規ハイブリッド融合タンパク質の製造において使用される又は本発明のウイルスベクターの製造において使用されるベクター/プラスミド/宿主にも及ぶ。 The invention also extends to a vector / plasmid / host used in the production of a novel hybrid fusion protein of SEQ ID NO: 1 or used in the production of a viral vector of the invention.
タンパク質の製造および単離が必要な場合には、該タンパク質をコードする配列を合成のための適当な宿主内に挿入するために、適当なプラスミドを使用することができる。適当なプラスミドの一例は、適当な発現カセットを運搬するための2ミクロンに基づくベクターであるpRIT15546である。該プラスミドは、一般に、選択を補助するための組込まれたマーカー、例えば、抗生物質耐性またはLEU2もしくはHIS栄養要求性をコードする遺伝子を含有する。 If protein production and isolation is required, a suitable plasmid can be used to insert the sequence encoding the protein into a suitable host for synthesis. An example of a suitable plasmid is pRIT15546, a 2 micron based vector for carrying a suitable expression cassette. The plasmid generally contains an integrated marker to aid selection, eg, a gene encoding antibiotic resistance or LEU2 or HIS auxotrophy.
宿主細胞は原核生物または真核生物でありうるが、好ましくは、酵母、例えばSaccharomyces(例えば、Saccharomyces cerevisiae、例えば、名称RIT DC5 cir(o)、寄託者Smith Kline-RITであるATCCデータベース(受託番号20820)におけるDC5)および非Saccharomyces酵母である。これらには、Schizosaccharomyces(例えば、Schizosaccharomyces pombe)、Kluyveromyces(例えば、Kluyveromyces lactis)、Pichia(例えば、Pichia pastoris)、Hansenula(例えば、Hansenula polymorpha)、Yarrowia(例えば、Yarrowia lipolytica)およびSchwanniomyces(例えば、Schwanniomyces occidentalis)が含まれる。 The host cell can be prokaryotic or eukaryotic, but is preferably a yeast, such as Saccharomyces (eg, Saccharomyces cerevisiae, eg, name RIT DC5 cir (o), ATCC database with depositor Smith Kline-RIT (accession number) 20820) DC5) and non-Saccharomyces yeasts. These include Schizosaccharomyces (e.g. Schizosaccharomyces pombe), Kluyveromyces (e.g. Kluyveromyces lactis), Pichia (e.g. Pichia pastoris), Hansenula (e.g. Hansenula polymorpha), Yarrowia (e.g. Yarrowia lipolyticaces (e.g. Yarrowia lipolyticaces) ) Is included.
1つの態様においては、本発明は、マラリアの治療または予防のための、本発明のベクターまたは配列番号1のタンパク質の使用を提供する。 In one embodiment, the present invention provides the use of the vector of the present invention or the protein of SEQ ID NO: 1 for the treatment or prevention of malaria.
1つの態様においては、本発明は、本発明のウイルスベクターと、賦形剤、例えば注射に適した等張性担体とを含む医薬製剤を提供する。適当な賦形剤は、後記において、より詳しく説明されている。 In one embodiment, the present invention provides a pharmaceutical formulation comprising a viral vector of the present invention and an excipient, eg, an isotonic carrier suitable for injection. Suitable excipients are described in more detail below.
1つの実施形態においては、製剤は、
・本発明のアデノウイルスベクター、
・マラリア抗原、例えばリポタンパク質粒子、特にRTS,S、および
・場合によっては、アジュバント、例えば、サポニンおよび/または3D-MPLを含むアジュバント
を含む。
In one embodiment, the formulation is
The adenoviral vector of the present invention,
A malaria antigen, such as lipoprotein particles, in particular RTS, S, and optionally an adjuvant, such as an adjuvant comprising saponin and / or 3D-MPL.
ベクターが配列番号1の配列をコードする場合、該ベクターは、RTS,Sとして公知のタンパク質を使用する治療計画における使用に特に適している。これは、アデノウイルスベクターによりコードされるタンパク質が、可能な限り厳密に、RTS,Sにおける「RT」成分に対応しているからである。RTS,Sを使用する計画における該ベクターの使用はRTS,Sの効力を効率的に増強しうると考えられる。 If the vector encodes the sequence of SEQ ID NO: 1, the vector is particularly suitable for use in therapeutic regimes using a protein known as RTS, S. This is because the protein encoded by the adenoviral vector corresponds as closely as possible to the “RT” component in RTS, S. It is believed that the use of the vector in a plan to use RTS, S can effectively enhance the efficacy of RTS, S.
本明細書に記載されているウイルスベクターは、マラリアワクチンの成分としての使用に適している。本発明のウイルスベクターは、感染に対する適度な保護をもたらすために他の抗原を含む他の成分と組合せて使用される必要がありうる。それでも、本発明のベクターは、少なくとも、ワクチンまたは治療計画の成分としての使用に適している。 The viral vectors described herein are suitable for use as a component of a malaria vaccine. The viral vectors of the present invention may need to be used in combination with other components, including other antigens, to provide reasonable protection against infection. Nevertheless, the vectors of the present invention are at least suitable for use as a component of a vaccine or therapeutic regime.
RTS,S
RTS,Sは、(例えば、P. falciparum NF54/3D7株から)WO 93/10152に記載されているとおりに製造されうる。RTS発現カセットおよび推定翻訳産物のヌクレオチド配列はWO 93/10152の図9(そこにおいては、RTS*と称されている)に記載されている。
RTS, S
RTS, S can be produced as described in WO 93/10152 (eg from P. falciparum NF54 / 3D7 strain). The nucleotide sequence of the RTS expression cassette and putative translation product is described in FIG. 9 of WO 93/10152 (in which it is referred to as RTS * ).
本明細書の文脈においては、賦形剤は、それ自体は治療効果を有さない、医薬製剤中の成分を意味する。希釈剤または担体は賦形剤の定義に含まれる。適当な担体には、PBS、塩類液(食塩水)などが含まれる。アジュバントも賦形剤の定義に含まれる。なぜなら、アジュバントはin vivoで生理的効果を示しうるが、この効果は全体的なものであり、治療成分の非存在下では特異的治療効果ではないからである。 In the context of the present specification, an excipient means an ingredient in a pharmaceutical formulation that itself has no therapeutic effect. Diluents or carriers are included in the definition of excipients. Suitable carriers include PBS, saline (saline) and the like. Adjuvants are also included in the definition of excipients. This is because adjuvants can show physiological effects in vivo, but this effect is global and not a specific therapeutic effect in the absence of therapeutic ingredients.
アジュバント
個々のアジュバントは、金属塩、水中油型エマルション、Toll様受容体アゴニスト(特にToll様受容体2アゴニスト、Toll様受容体3アゴニスト、Toll様受容体4アゴニスト、Toll様受容体7アゴニスト、Toll様受容体8アゴニストおよびToll様受容体9アゴニスト)、サポニンまたはそれらの組合せの群から選ばれるアジュバントである。
Adjuvants Individual adjuvants include metal salts, oil-in-water emulsions, Toll-like receptor agonists (especially Toll-
1つの実施形態においては、該アジュバントはToll様受容体(TLR)4リガンド、例えばアゴニスト、例えばリピドA誘導体、特にモノホスホリルリピドA、より詳しくは3-デアシル化モノホスホリルリピドA(3D-MPL)である。 In one embodiment, the adjuvant is a Toll-like receptor (TLR) 4 ligand, such as an agonist, such as a lipid A derivative, in particular monophosphoryl lipid A, more particularly 3-deacylated monophosphoryl lipid A (3D-MPL). It is.
3-デアシル化モノホスホリルリピドAは米国特許第4,912,094号および英国特許出願第2,220,211号(Ribi)から公知であり、Ribi Immunochem, Montana, USAから入手可能である。 3-Deacylated monophosphoryl lipid A is known from US Pat. No. 4,912,094 and UK Patent Application No. 2,220,211 (Ribi) and is available from Ribi Immunochem, Montana, USA.
3D-MPLはCorixa社により商標MPL(登録商標)で販売されており、主として、IFN-g(Th1)表現型を伴うCD4+ T細胞の応答を促進する。それは、GB 2 220 211 Aに開示されている方法に従い製造されうる。化学的には、それは、3、4、5または6本のアシル化鎖を有する3-デアシル化モノホスホリルリピドAの混合物である。一般に、本発明の組成物においては、小粒子3D-MPLが使用される。小粒子3-MPLは、それが0.22μmフィルターで滅菌濾過されうるような粒径を有する。そのような調製物はWO 94/21292に記載されている。リピドAの合成誘導体は公知であり、以下のものを含む(それらに限定されるものではない)TLR4アゴニストであると考えられている。
3D-MPL is sold by Corixa under the trademark MPL® and primarily promotes the response of CD4 + T cells with an IFN-g (Th1) phenotype. It can be manufactured according to the method disclosed in
OM174 (2-デオキシ-6-O-[2-デオキシ-2-[(R)-3-ドデカノイルオキシテトラ-デカノイルアミノ]-4-o-ホスホノ-β-D-グルコピラノシル]-2-[(R)-3-ヒドロキシテトラデカノイルアミノ]-α-D-グルコピラノシルジヒドロゲンホスファート)(WO 95/14026)、
OM 294 DP (3S,9R)-3-[(R)-ドデカノイルオキシテトラデカノイルアミノ]-4-オキソ-5-アザ-9(R)-[(R)-3-ヒドロキシテトラデカノイルアミノ]デカン-1,10-ジオール,1,10-ビス(ジヒドロゲノホスファート)(WO99/64301およびWO 00/0462)、
OM 197 MP-Ac DP (3S-,9R)-3-[(R)-ドデカノイルオキシテトラデカノイルアミノ]-4-オキソ-5-アザ-9-[(R)-3-ヒドロキシテトラデカノイルアミノ]デカン-1,10-ジオール,1-ジヒドロゲノホスファート 10-(6-アミノヘキサノアート)(WO 01/46127)。
OM174 (2-deoxy-6-O- [2-deoxy-2-[(R) -3-dodecanoyloxytetra-decanoylamino] -4-o-phosphono-β-D-glucopyranosyl] -2- [ (R) -3-hydroxytetradecanoylamino] -α-D-glucopyranosyl dihydrogen phosphate) (WO 95/14026),
OM 294 DP (3S, 9R) -3-[(R) -Dodecanoyloxytetradecanoylamino] -4-oxo-5-aza-9 (R)-[(R) -3-hydroxytetradecanoylamino Decane-1,10-diol, 1,10-bis (dihydrogenophosphate) (WO99 / 64301 and WO 00/0462),
OM 197 MP-Ac DP (3S-, 9R) -3-[(R) -Dodecanoyloxytetradecanoylamino] -4-oxo-5-aza-9-[(R) -3-hydroxytetradecanoyl Amino] decane-1,10-diol, 1-dihydrogenophosphate 10- (6-aminohexanoate) (WO 01/46127).
典型的には、3D-MPLが使用される場合、抗原および3D-MPLは水中油型エマルションまたは多重水中油型エマルションとして送達される。3D-MPLの取り込みは、それがエフェクターT細胞応答の刺激物質であることから、有利である。 Typically, when 3D-MPL is used, the antigen and 3D-MPL are delivered as an oil-in-water emulsion or multiple oil-in-water emulsions. The uptake of 3D-MPL is advantageous because it is a stimulator of effector T cell responses.
使用されうる他のTLR4リガンドとしては、アルキルグルコサミニドホスファート(AGP)、例えば、WO 9850399もしくはUS 6303347(AGPの製造方法も開示されている)に開示されているもの、またはUS 6764840に開示されているAGPの製薬上許容される塩が挙げられる。いくつかのAGPはTLR4アゴニストであり、いくつかはTLR4アンタゴニストである。いずれもアジュバントとして有用であると考えられる。 Other TLR4 ligands that can be used include alkyl glucosaminide phosphates (AGP), such as those disclosed in WO 9850399 or US 6303347 (a method for producing AGP is also disclosed), or disclosed in US 6764840 And pharmaceutically acceptable salts of AGP. Some AGPs are TLR4 agonists and some are TLR4 antagonists. Both are considered useful as adjuvants.
本発明において使用されるもう1つの免疫刺激物質はQuil(クイル)Aおよびその誘導体である。Quil Aは、南米樹木Quilaja Saponaria Molinaから単離されたサポニン調製物であり、1974年にDalsgaardら(“Saponin adjuvants”, Archiv. fur die gesamte Virusforschung, Vol. 44, Springer Verlag, Berlin, p243-254)により、アジュバント活性を有すると最初に記載された。Quil Aに伴う毒性を有することなくアジュバント活性を保有する、Quil Aの精製断片、例えばQS7およびQS21(QA7およびQA21としても公知である)が、HPLCにより単離されている。QS21は、CD8+細胞傷害性T細胞(CTL)、Th1細胞および優勢なIgG2a抗体応答を誘導する、Quillaja saponaria Molinaの樹皮に由来する天然サポニンである。 Another immunostimulant used in the present invention is Quil A and its derivatives. Quil A is a saponin preparation isolated from the South American tree Quilaja Saponaria Molina. In 1974, Dalsgaard et al. (“Saponin adjuvants”, Archiv. Fur die gesamte Virusforschung, Vol. 44, Springer Verlag, Berlin, p243-254 ) Was first described as having adjuvant activity. Purified fragments of Quil A, such as QS7 and QS21 (also known as QA7 and QA21), which retain adjuvant activity without the toxicity associated with Quil A, have been isolated by HPLC. QS21 is a natural saponin derived from the bark of Quillaja saponaria Molina that induces CD8 + cytotoxic T cells (CTL), Th1 cells and a prevalent IgG2a antibody response.
ステロールを更に含むQS21の特定の製剤が記載されている(WO 96/33739)。QS21:ステロールの比は、典型的には、1:100〜1:1(重量対重量)のオーダーである。一般に、過剰なステロールが存在し、QS21:ステロールの比は少なくとも1:2 w/wである。典型的には、ヒトへの投与では、QS21およびステロールは、用量当たり約1μg〜約100μg、例えば約10μg〜約50μgの範囲で、ワクチン中に存在する。 Specific formulations of QS21 that further contain sterols have been described (WO 96/33739). The ratio of QS21: sterol is typically on the order of 1: 100 to 1: 1 (weight to weight). In general, there is an excess of sterol and the ratio of QS21: sterol is at least 1: 2 w / w. Typically, for human administration, QS21 and sterol are present in the vaccine in the range of about 1 μg to about 100 μg, such as about 10 μg to about 50 μg per dose.
リポソーム製剤は、一般に、中性脂質、例えばホスファチジルコリン(これは通常、室温で非結晶性である)、例えば卵黄ホスファチジルコリン、ジオレオイルホスファチジルコリンまたはジラウリルホスファチジルコリンを含有する。該リポソームは、飽和脂質から構成されるリポソームの場合、リポソーム-QS21構造の安定性を増強する荷電脂質をも含有しうる。これらの場合、荷電脂質の量は、しばしば、1〜20% w/w、例えば5〜10%である。リン脂質に対するステロールの比率は1〜50%(mol/mol)、例えば20〜25%である。 Liposomal formulations generally contain a neutral lipid, such as phosphatidylcholine (which is usually amorphous at room temperature), such as egg yolk phosphatidylcholine, dioleoylphosphatidylcholine or dilaurylphosphatidylcholine. The liposome may also contain a charged lipid that enhances the stability of the liposome-QS21 structure in the case of liposomes composed of saturated lipids. In these cases, the amount of charged lipid is often 1-20% w / w, for example 5-10%. The ratio of sterol to phospholipid is 1-50% (mol / mol), for example 20-25%.
これらの組成物は、MPL(3D-MPLとしても公知である3-デアシル化モノ-ホスホリルリピドA)を含有しうる。3D-MPLは、4、5または6本のアシル化鎖を有する3種類のデ-O-アシル化モノホスホルリピドAの混合物としてGB 2 220 211(Ribi)にて公知であり、Ribi Immunochem, Montanaにより製造されている。
These compositions may contain MPL (3-deacylated mono-phosphoryl lipid A, also known as 3D-MPL). 3D-MPL is known in
サポニンはミセル、混合ミセル(限定的なものではないが一般には胆汁酸塩を伴う)の形態であることが可能であり、あるいはISCOMマトリックス(EP 0 109 942)、リポソームまたは関連コロイド構造、例えば虫状(らせん状)または環状多量体複合体または脂質/層化構造、およびラメラ(コレステロールおよび脂質と共に製剤化された場合)の形態、あるいは水中油型エマルション(例えば、WO 95/17210におけるもの)の形態であることが可能である。
Saponins can be in the form of micelles, mixed micelles (generally but not exclusively with bile salts), or ISCOM matrices (
通常、該サポニンは、リポソーム製剤、ISCOMまたは水中油型エマルションの形態で提供される。 Usually, the saponin is provided in the form of a liposomal formulation, ISCOM or an oil-in-water emulsion.
免疫刺激性オリゴヌクレオチドも使用されうる。本発明のアジュバントまたはワクチンにおいて使用されるオリゴヌクレオチドの具体例には、CpG含有オリゴヌクレオチド、一般には、少なくとも3つ、より好ましくは少なくとも6つ以上のヌクレオチドにより隔てられた2以上のジヌクレオチドCpGモチーフを含有するオリゴヌクレオチドが含まれる。CpGモチーフはシトシンヌクレオチドおよびそれに続くグアニンヌクレオチドである。CpGオリゴヌクレオチドは典型的にはデオキシヌクレオチドである。1つの実施形態においては、該オリゴヌクレオチドにおけるヌクレオチド間には、ホスホロジチオアート、またはより好ましくはホスホロチオアート結合が存在するが、ホスホジエステルおよび他のヌクレオチド間結合も本発明の範囲内である。混合ヌクレオチド間連結を有するオリゴヌクレオチドも本発明の範囲内に含まれる。ホスホロチオアートオリゴヌクレオチドまたはホスホロジチオアートの製造方法はUS 5,666,153、US 5,278,302およびWO 95/26204に記載されている。 Immunostimulatory oligonucleotides can also be used. Specific examples of oligonucleotides used in the adjuvants or vaccines of the present invention include CpG-containing oligonucleotides, generally two or more dinucleotide CpG motifs separated by at least 3, more preferably at least 6 or more nucleotides. Are included. A CpG motif is a cytosine nucleotide followed by a guanine nucleotide. CpG oligonucleotides are typically deoxynucleotides. In one embodiment, there is a phosphorodithioate, or more preferably a phosphorothioate linkage, between nucleotides in the oligonucleotide, although phosphodiester and other internucleotide linkages are also within the scope of the present invention. is there. Oligonucleotides having mixed internucleotide linkages are also included within the scope of the present invention. Methods for the production of phosphorothioate oligonucleotides or phosphorodithioates are described in US 5,666,153, US 5,278,302 and WO 95/26204.
オリゴヌクレオチドの具体例としては以下のものが挙げられる:
他のCpGオリゴヌクレオチドは、重要でない欠失または付加を伴う前記の1以上の配列を含みうる。 Other CpG oligonucleotides may contain one or more of the above sequences with minor deletions or additions.
該CpGオリゴヌクレオチドは、当技術分野で公知のいずれかの方法(例えば、EP 468520を参照されたい)により合成されうる。簡便には、そのようなオリゴヌクレオチドは、自動合成装置を使用して合成されうる。 The CpG oligonucleotide can be synthesized by any method known in the art (see, eg, EP 468520). Conveniently, such oligonucleotides can be synthesized using an automated synthesizer.
TLR2アゴニストの具体例には、ペプチドグリカンまたはリポタンパク質が含まれる。 Specific examples of TLR2 agonists include peptidoglycan or lipoprotein.
ImiquimodおよびResiquimodのようなイミダゾキノリンは公知TLR7アゴニストである。一本鎖RNAも公知TLRアゴニスト(ヒトにおけるTLR8およびマウスにおけるTLR7)であり、二本鎖RNAおよびポリIC(ウイルスRNAの市販合成擬似体であるポリイノシン-ポリシチジル酸)はTLR3アゴニストの典型例である。3D-MPLはTLR4アゴニストの一例であり、CpGはTLR9アゴニストの一例である。 Imidazoquinolines such as Imiquimod and Resiquimod are known TLR7 agonists. Single-stranded RNA is also a known TLR agonist (TLR8 in humans and TLR7 in mice), and double-stranded RNA and poly IC (polyinosin-polycytidylic acid, a commercially available synthetic mimic of viral RNA) are typical examples of TLR3 agonists . 3D-MPL is an example of a TLR4 agonist, and CpG is an example of a TLR9 agonist.
免疫刺激物質が代替的または追加的に含まれうる。1つの実施形態においては、この免疫刺激物質は3-デアシル化モノホスホリルリピドA(3D-MPL)である。 Immunostimulants can be included alternatively or additionally. In one embodiment, the immunostimulator is 3-deacylated monophosphoryl lipid A (3D-MPL).
1つの態様においては、該アジュバントは3D-MPLを含む。 In one embodiment, the adjuvant comprises 3D-MPL.
1つの態様においては、該アジュバントはQS21を含む。 In one embodiment, the adjuvant comprises QS21.
1つの態様においては、該アジュバントはCpGを含む。 In one embodiment, the adjuvant comprises CpG.
1つの態様においては、該アジュバントは水中油型エマルションとして配合される。 In one embodiment, the adjuvant is formulated as an oil-in-water emulsion.
1つの態様においては、該アジュバントはリポソームとして配合される。 In one embodiment, the adjuvant is formulated as a liposome.
アジュバントの組合せには、3D-MPLおよびQS21(EP 0 671 948 B1)水中油型エマルション、または他の担体と共に配合される3D-MPLおよびQS21もしくは3D-MPLを含むリポソーム製剤(EP 0 689 454 B1)が含まれる。他の好ましいアジュバント系は、US 6558670およびUS 6544518に記載されている3D-MPL、QS21およびCpGオリゴヌクレオチドの組合せを含む。
Adjuvant combinations include 3D-MPL and QS21 (
製剤
ワクチンの製造は、全般的には、New Trends and Developments in Vaccines, Vollerら編, University Park Press, Baltimore, Maryland, U.S.A., 1978に記載されている。リポソーム内への封入は、例えばFullerton, 米国特許第4,235,877号に記載されている。
The production of pharmaceutical vaccines is generally described in New Trends and Developments in Vaccines, Voller et al., University Park Press, Baltimore, Maryland, USA, 1978. Encapsulation within liposomes is described, for example, in Fullerton, US Pat. No. 4,235,877.
本発明の製剤は予防目的および治療目的の両方に使用されうる。したがって、本発明は、医薬における使用のための、例えば、マラリアの治療および/または予防のための、本明細書に記載のワクチン組成物を提供する。 The formulations of the present invention can be used for both prophylactic and therapeutic purposes. Accordingly, the present invention provides a vaccine composition as described herein for use in medicine, for example for the treatment and / or prevention of malaria.
1つの態様においては、本発明は、本発明のC7アデノウイルスベクター、およびマラリア抗原、例えばRTS,Sまたは配列番号1の新規抗原またはそのウイルス様粒子、および賦形剤を、場合によってはアジュバントの存在下で含む組成物を提供する。 In one embodiment, the present invention provides a C7 adenoviral vector of the present invention and a malaria antigen, such as a novel antigen of RTS, S or SEQ ID NO: 1 or a virus-like particle thereof, and an excipient, optionally an adjuvant. Compositions comprising in the presence are provided.
本明細書の文脈における免疫原性は、免疫応答を惹起する能力を意味すると意図され、ここで、該応答は関連製剤中のマラリア成分に特異的である。この応答は適当なアジュバントおよび/または追加抗原刺激の存在を要しうる。例えば、適当な免疫原性応答を得るためには、元の用量と同様の又はそれ未満の用量を含む追加抗原刺激(ブースター)が要求されうる。 Immunogenicity in the context of the present specification is intended to mean the ability to elicit an immune response, where the response is specific for the malaria component in the relevant formulation. This response may require the presence of appropriate adjuvants and / or booster stimuli. For example, in order to obtain an appropriate immunogenic response, a booster that includes a dose similar to or less than the original dose may be required.
本発明の組成物/医薬製剤は、1以上の他の抗原、例えばP. falcipariumおよび/またはP. vivaxに由来する抗原を、混合されて含むことが可能であり、ここで、該抗原は、DBP、PvTRAP、PvMSP2、PvMSP4、PvMSP5、PvMSP6、PvMSP7、PvMSP8、PvMSP9、PvAMA1およびRBPまたはその断片から選ばれる。 The composition / pharmaceutical formulation of the invention can comprise a mixture of one or more other antigens, such as antigens derived from P. falciparium and / or P. vivax, wherein the antigen comprises It is selected from DBP, PvTRAP, PvMSP2, PvMSP4, PvMSP5, PvMSP6, PvMSP7, PvMSP8, PvMSP9, PvAMA1 and RBP or fragments thereof.
P falciparumに由来する抗原の他の具体例には、PfEMP-1、Pfs 16抗原、MSP-1、MSP-3、LSA-1、LSA-3、AMA-1およびTRAPが含まれる。他のPlasmodium抗原には、P. falciparum EBA、GLURP、RAP1、RAP2、Sequestrin(セクエストリン)、Pf332、STARP、SALSA、PfEXP1、Pfs25、Pfs28、PFS27/25、Pfs48/45、Pfs230および他のPlasmodium属種におけるそれらの類似体(アナログ)が含まれる。
Other specific examples of antigens derived from P falciparum include PfEMP-1,
本発明はまた、マラリア抗原をコードするための、例えば、特に、本明細書に記載されているとおり、マラリアの治療および/または予防のための、あるいはそれに対する医薬の製造のための、C7の使用に関する。 The present invention also relates to the use of C7 for encoding malaria antigens, for example, for the treatment and / or prevention of malaria, or for the manufacture of medicaments thereto, particularly as described herein. Regarding use.
本発明はまた、本発明の1以上の態様の治療的有効量を投与することを含む治療方法を含む。場合によっては、本発明のC7ウイルスベクターは、場合によっては例えば3D-MPLおよび/またはQS21のようなサポニンを含むアジュバントの存在下、マラリア抗原、例えばRTS,Sまたは配列番号1の抗原と共に共投与または共配合されうる。 The present invention also includes a method of treatment comprising administering a therapeutically effective amount of one or more embodiments of the present invention. In some cases, a C7 viral vector of the invention is co-administered with a malaria antigen, such as RTS, S or an antigen of SEQ ID NO: 1, optionally in the presence of an adjuvant comprising saponins such as 3D-MPL and / or QS21. Or it can be co-blended.
C7ベクターはまた、同じ又は異なる抗原をコードする異なる血清型および/または起源の別のアデノウイルスベクターと共に共投与または共配合されうる。 A C7 vector can also be co-administered or co-formulated with another adenoviral vector of different serotype and / or origin encoding the same or different antigens.
本発明は、初回-追加抗原刺激法における、本明細書に記載のいずれかの態様の使用にも及び、例えば、この場合、時点ゼロにおいて初回抗原刺激量(そして例えば3ヶ月以内に後続の初回抗原刺激)が投与され、最終初回抗原投与の約4、5、6、7、8、9、10、11または12週後に追加抗原が投与され、場合によっては、最初の追加抗原投与の1年後までに、更なる追加抗原が投与される。
The invention extends to the use of any of the embodiments described herein in a prime-boost method, eg, in this case the prime dose at time zero (and the subsequent prime within
有利には、前記の組合せワクチンを含む本発明の1以上の態様は、特異的体液性(すなわち、抗体応答)および/または細胞性免疫応答(例えば、CD8+および/またはCD4+)、例えば抗体応答ならびにCD8+および/またはCD4+応答、特にCD8+および抗体応答、すなわち、(適宜)CSタンパク質および/またはS抗原に特異的な応答を刺激する。 Advantageously, one or more embodiments of the present invention comprising the combination vaccines described above are specific humoral (ie antibody responses) and / or cellular immune responses (eg CD8 + and / or CD4 +), eg antibody responses and Stimulate a CD8 + and / or CD4 + response, in particular a CD8 + and antibody response, i.e. a response specific to the CS protein and / or S antigen, as appropriate.
このタイプのバランス化された免疫応答は、マラリア感染に対するいわゆる無菌(sterile)保護を得るために要求されうる。 This type of balanced immune response may be required to obtain so-called sterile protection against malaria infection.
さらに、組合せの抗体応答は、アジュバント化タンパク質のみの方法に対する抗体応答と比較して増強されうる。 Furthermore, the combined antibody response can be enhanced compared to the antibody response to the adjuvanted protein-only method.
1つの実施形態においては、本発明は、初回-追加抗原刺激法の補足的成分として、
・同じ又は異なるマラリア抗原をコードするC7アデノウイルスベクター、
・マラリア抗原、例えばP. falciparum由来のCSタンパク質をコードする別のウイルスベクター、例えばヒトアデノウイルスベクター、例えばAd5もしくはAd35、または異なる血清型(すなわち、C7以外)のサルアデノウイルスベクター、ならびに/または
・マラリア抗原、例えばRTS,S、ならびにアジュバント、例えばサポニンおよび/または3D-MPLを含むアジュバント
を使用する該初回-追加抗原刺激法における初回抗原または追加抗原としてのC7の使用を提供する。
In one embodiment, the present invention provides a supplemental component of the first-boost stimulation method,
A C7 adenoviral vector encoding the same or different malaria antigen,
Another viral vector encoding a malaria antigen, for example a CS protein from P. falciparum, for example a human adenoviral vector, for example Ad5 or Ad35, or a simian adenoviral vector of a different serotype (ie other than C7), and / or Provide the use of C7 as primary or booster in the first-boost stimulation method using malaria antigens such as RTS, S and adjuvants such as adjuvants including saponin and / or 3D-MPL.
本発明はまた、マラリア感染の治療および/または予防のための医薬の製造のための、本明細書に記載されている態様のいずれかを提供する。 The present invention also provides any of the embodiments described herein for the manufacture of a medicament for the treatment and / or prevention of malaria infection.
量
使用する3D-MPLの量は一般に少量であるが、ワクチン製剤に応じて、1〜1000μg/用量、例えば1〜500μg/用量の範囲、例えば1〜100μg/用量、例えば50または25μg/用量の範囲でありうる。
The amount of 3D-MPL used is generally small, but depending on the vaccine formulation, it is 1-1000 μg / dose, for example in the range of 1-500 μg / dose, for example 1-100 μg / dose, for example 50 or 25 μg / dose. Can be a range.
本発明のアジュバントまたはワクチンにおけるCpGまたは免疫刺激性オリゴヌクレオチドの量は一般に少量であるが、ワクチン製剤に応じて、1〜1000μg/用量、例えば1〜500μg/用量の範囲、例えば1〜100μg/用量の範囲でありうる。 The amount of CpG or immunostimulatory oligonucleotide in the adjuvants or vaccines of the invention is generally small, but depending on the vaccine formulation, 1-1000 μg / dose, such as in the range of 1-500 μg / dose, such as 1-100 μg / dose. Range.
本発明のアジュバントにおいて使用するサポニンの量は1〜1000μg/用量、例えば1〜500μg/用量、例えば1〜250μg/用量の範囲、特に1〜100μg/用量、例えば50または25μg/用量の範囲でありうる。 The amount of saponin used in the adjuvant of the present invention is 1-1000 μg / dose, for example 1-500 μg / dose, for example in the range 1-250 μg / dose, in particular 1-100 μg / dose, for example in the range 50 or 25 μg / dose. sell.
タンパク質を投与する場合、その用量は、例えば1〜500μg、例えば10〜100μg、特に20、25、30、35、40、45、50、55、60、65、70、75または80μg/用量でありうる。 When administering the protein, the dose is for example 1 to 500 μg, for example 10 to 100 μg, in particular 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75 or 80 μg / dose sell.
アデノウイルスベクターを投与する場合、その用量は、例えば103〜1016 vpu、例えば106〜1010 vpuでありうる。 When administering an adenoviral vector, the dose can be, for example, 10 3 to 10 16 vpu, such as 10 6 to 10 10 vpu.
組合せを用いる場合、該組合せの各成分に用いられる量は、その成分が単独で投与される量に対応しうる。 When a combination is used, the amount used for each component of the combination may correspond to the amount that the component is administered alone.
本発明は、本発明の組合せにおいて使用される要素を含むキットにも及ぶ。 The invention also extends to kits containing the elements used in the combination of the invention.
本発明は更に、本発明のアデノウイルスベクターの製造方法およびそれを含む製剤に関する。 The present invention further relates to a method for producing the adenoviral vector of the present invention and a preparation containing the same.
本発明はまた、配列番号1のタンパク質の製造方法に関する。 The present invention also relates to a method for producing the protein of SEQ ID NO: 1.
本明細書の文脈においては、含んでなる、とは、含む、と解釈されるべきである。 In the context of this specification, comprising shall be construed as comprising.
本発明は、或る要素を含むと本明細書に記載されている実施形態に対応する、関連要素からなる又は関連要素から実質的になる実施形態に及ぶ。 The invention extends to embodiments consisting of or consisting essentially of related elements, including certain elements, corresponding to the embodiments described herein.
本明細書の背景の節における考察は、本発明をその技術的背景内に置くという目的で記載されている。それは、当技術分野において何が公知であるのかについての自認であると解釈されるべきではなく、特に、何が技術常識に相当するのかの自認ではない。 The discussion in the background section of this specification is described for the purpose of placing the invention within its technical background. It should not be construed as an admission of what is known in the art, and in particular, no admission of what is equivalent to common general knowledge.
(実施例)
以下の実施例を、本発明に用いることのできる方法論を例示するために示す。
(Example)
The following examples are presented to illustrate the methodologies that can be used in the present invention.
Medigenomix社により合成遺伝子が製造された。該遺伝子をpCR2.1-TOPO-TAクローニングベクター(Invitrogen;図1を参照されたい)内にクローニングした。このベクターをNotIおよびBamHIで消化し、組換えシャトルプラスミドベクター(-Ade2)を作製した。該シャトルプラスミドの地図を図2に示す。 A synthetic gene was produced by Medigenomix. The gene was cloned into the pCR2.1-TOPO-TA cloning vector (Invitrogen; see Figure 1). This vector was digested with NotI and BamHI to produce a recombinant shuttle plasmid vector (-Ade2). A map of the shuttle plasmid is shown in FIG.
ウイルスレスキューのための方法論および発現カセットの説明
発現カセットは、サイトメガロウイルス(CMV)初期プロモーターおよび第1エキソン、プラスミドpCI(Promegaから購入)由来のイントロン、Ade2をコードするDNA、およびウサギグロビンポリアデニル化シグナルを含有する。該完全カセットは、それぞれ制限酵素I-CeuIおよびPI-SceIに対する認識部位に隣接している。該発現カセットを、I-CeuIおよびPI-SceIを使用して該シャトルプラスミドから切り出し、SAdV-24のE1欠失ゲノム(すなわちC7)-pC7000pkGFP(Royら, Hum Gene Ther. (2004) 5:519-530に記載されている)のプラスミド分子クローン内に導入して、図3に示すプラスミドを得た。該プラスミド分子クローンDNAを制限酵素PacIでの消化により線状化し、HEK293細胞内にトランスフェクトして組換えアデノウイルスをレスキューした。標準的な技術を用いて、該アデノウイルスを増殖させ、増幅し、精製した。
Methodology and Expression Cassette Description for Viral Rescue Expression cassettes are cytomegalovirus (CMV) early promoter and first exon, intron from plasmid pCI (purchased from Promega), DNA encoding Ade2, and rabbit globin polyadenylation Contains signal. The complete cassette is flanked by recognition sites for restriction enzymes I-CeuI and PI-SceI, respectively. The expression cassette was excised from the shuttle plasmid using I-CeuI and PI-SceI and the SA1V-24 E1-deleted genome (ie C7) -pC7000pkGFP (Roy et al., Hum Gene Ther. (2004) 5: 519 Was introduced into the plasmid molecular clone described in FIG. The plasmid molecular clone DNA was linearized by digestion with the restriction enzyme PacI and transfected into HEK293 cells to rescue the recombinant adenovirus. The adenovirus was propagated, amplified and purified using standard techniques.
I-CeuIからPI-Sce認識部位までの(Ade2)の発現カセットの配列を配列番号18に示す。 The sequence of the expression cassette (Ade2) from I-CeuI to the PI-Sce recognition site is shown in SEQ ID NO: 18.
C57Bl/6マウスにおけるC7-Ade1およびC7-Ade2の免疫原性
構築物Ade1またはAde2のいずれかを発現するC7チンプアデノウイルス(chimpadenovirus)の、10e10、10e9、10e8ウイルス粒子の用量範囲で、C57Bl/6マウスを筋肉内に1回、免疫化した。陽性対照として、構築物Ade1またはAde2のいずれかを発現するヒトアデノウイルス5(10e9および10e8の用量)で数匹のマウスを免疫化した。陰性対照として、空C7および空Ad5ウイルスベクターで数匹のマウスを免疫化した。
C57Bl / 6 in C57Bl / 6 mice at a dose range of 10e10, 10e9, 10e8 virions of C7 chimpadenovirus expressing either Ade1 or Ade2 immunogenic constructs of C7-Ade1 and C7-Ade2 in C57Bl / 6 mice Mice were immunized once intramuscularly. As a positive control, several mice were immunized with human adenovirus 5 (10e9 and 10e8 doses) expressing either construct Ade1 or Ade2. As negative controls, several mice were immunized with empty C7 and empty Ad5 viral vectors.
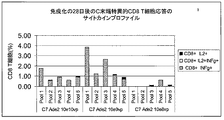
免疫化後第14日、28日、34日および49日に末梢血を集め、プールし、対象の配列、すなわち、CSタンパク質のN末端領域(N末端)またはC末端領域(C末端)を包含する15マー(15量体)ペプチドのプールでの一晩のin vitro再刺激の後、IL-2および/またはIFN-ガンマを産生するAg特異的CD4およびCD8 T細胞応答をフローサイトメトリーにより測定した。また、陰性対照として、幾つかの細胞を培地内でin vitroで一晩培養した(未刺激)。該ペプチド刺激細胞によりもたらされた平均サイトカイン応答から、未刺激細胞によりもたらされた平均サイトカイン応答を差し引くことにより、該Ag特異的応答を計算した。
Peripheral blood is collected and pooled on
結果は、前記の実験条件下、両方の構築物がCS特異的CD4およびCD8 T細胞応答を誘導したことを示している(図4〜15)。注目すべきことに、C末端特異的CD8 T細胞応答は、Ade2インサートを含有するアデノウイルスで免疫化されたマウスにおいてのみ検出された(図4および8)。特に、これらのC末端特異的CD8 T細胞応答は、10e10vp のC7 Ade2または10e9vp のAd5 Ade2で免疫化されたマウスにおいて類似していた(図8)。 The results show that both constructs induced CS-specific CD4 and CD8 T cell responses under the experimental conditions described above (FIGS. 4-15). Of note, C-terminal specific CD8 T cell responses were detected only in mice immunized with adenovirus containing the Ade2 insert (FIGS. 4 and 8). In particular, these C-terminal specific CD8 T cell responses were similar in mice immunized with 10e10vp C7 Ade2 or 10e9vp Ad5 Ade2 (FIG. 8).
また、免疫化の48日後に集めた血清に関してELISAにより抗CS抗体応答を測定した。特に、測定したのは、R32LRポリペプチド(すなわち、P. falciparum CSPの中央部分を包含するもの)に対する全Ig応答であった(Mettensら, Vaccine 2008)。結果は、C7 Ade1またはC7 Ade2での1回の免疫化が低レベルのR32LR特異的抗体応答を誘導することを示している。この応答の強度は、免疫化に使用されるウイルス粒子の数と相関する(用量範囲効果)。 Anti-CS antibody responses were also measured by ELISA on sera collected 48 days after immunization. In particular, the total Ig response to the R32LR polypeptide (ie, including the central portion of P. falciparum CSP) was measured (Mettens et al., Vaccine 2008). The results show that a single immunization with C7 Ade1 or C7 Ade2 induces a low level of R32LR specific antibody response. The intensity of this response correlates with the number of virus particles used for immunization (dose range effect).
CB6F1マウスにおけるC7-Ade2の免疫原性
Ade2構築物を発現するC7チンプアデノウイルス(chimpadenovirus)の、10e10、10e9、10e8ウイルス粒子の用量範囲で、CB6F1マウスを筋肉内に1回、免疫化した(5プールのマウス/群)。免疫化後第21日、28日および35日に末梢血を集め、プールし、対象の配列、すなわち、CSタンパク質のN末端領域(N末端)またはC末端領域(C末端)を包含する15マーペプチドのプールでの一晩のin vitro再刺激の後、IL-2および/またはIFN-ガンマを産生するCS C末端およびCS N末端特異的CD4およびCD8 T細胞応答をフローサイトメトリーにより測定した。また、陰性対照として、幾つかの細胞を培地内でin vitroで一晩培養した(未刺激)。該ペプチド刺激細胞によりもたらされた平均サイトカイン応答から、未刺激細胞によりもたらされた平均サイトカイン応答を差し引くことにより、該Ag特異的応答を計算した。
Immunogenicity of C7-Ade2 in CB6F1 mice
CB6F1 mice were immunized once intramuscularly (5 pools of mice / group) at a dose range of 10e10, 10e9, 10e8 virions of C7 chimpadenovirus expressing Ade2 construct. Peripheral blood was collected and pooled on
結果は、このマウス系統においては、10e9vpまたは10e10vpのC7 Ade2での1回の免疫化がC末端およびN末端特異的CD4およびCD8 T細胞応答を誘導することを示している(図17〜20)。特に、平均観察応答の強度は、10e9vp用量を使用した場合には、より高くなかったとしても、少なくとも同等である。CS特異的CD8 T細胞応答は主としてN末端特異的であるが、CS特異的CD4 T細胞応答はCSタンパク質のN末端およびC末端領域を同等に標的化している。 The results show that in this mouse strain, a single immunization with 10e9vp or 10e10vp C7 Ade2 induces C-terminal and N-terminal specific CD4 and CD8 T cell responses (FIGS. 17-20). . In particular, the intensity of the average observed response is at least equivalent if not higher when the 10e9vp dose is used. While the CS-specific CD8 T cell response is primarily N-terminal specific, the CS-specific CD4 T cell response equally targets the N-terminal and C-terminal regions of the CS protein.
CS特異的CD4およびCD8 T細胞応答のサイトカインプロファイルも決定された。それは、試験された全ての時点にわたって類似していた。免疫化の28日後に示されたプロファイルが図21〜24に示されており、試験されたその他の時点を代表するものである。簡潔に説明すると、該CS特異的CD8 T細胞応答は、大部分は、IFNgのみを産生するCD8 T細胞によりもたらされる(図21および22)。また、該CS特異的CD4 T細胞応答はIFNg産生CD4 T細胞、そしてそれより低い度合ではあるが、IL-2のみ又はIL-2およびIFNgの両方を産生するCD4 T細胞によりもたらされる(図23および24)。 Cytokine profiles of CS-specific CD4 and CD8 T cell responses were also determined. It was similar across all time points tested. The profiles shown 28 days after immunization are shown in FIGS. 21-24 and are representative of other time points tested. Briefly, the CS-specific CD8 T cell response is largely mediated by CD8 T cells that produce only IFNg (FIGS. 21 and 22). The CS-specific CD4 T cell response is also caused by IFNg-producing CD4 T cells and, to a lesser extent, CD2 T cells that produce only IL-2 or both IL-2 and IFNg (FIG. 23). And 24).
CB6F1マウスにおけるRTS,S/AS01Bを使用する初回/追加抗原刺激または共配合におけるC7-Ade2の免疫原性
本発明者らは、CB6F1マウスにおけるRTS,S/AS01Bを使用する初回-追加抗原刺激または共配合(コンボ)においてAde2構築物を発現するC7チンプアデノウイルス(chimpadenovirus)の免疫原性を試験した(4プールのマウス/群)。AS01Bは、リポソームで配合された3D-MPLおよびQS21を含有するアジュバント系である。以下のとおりに、第0日、14日および28日にマウスを筋肉内に免疫化した。
ここで、
A = 10e9 vpのC7 Ade2、
P = 5μg RTS,S/50μl AS01B、
C = 10e10 vpのC7 Ade2+ 5μg RTS,S/50μl AS01B。
here,
A = 10e9 vp C7 Ade2,
P = 5μg RTS, S / 50μl AS01B,
C = 10e10 vp C7 Ade2 + 5 μg RTS, S / 50 μl AS01B.
免疫化後第21日(7d pII)、35日(7d pIII)、49日(21d pIII)、63日(35d pIII)、77日(49d pIII)に末梢血を集め、プールし、対象の配列(CS N末端、CS C末端またはHBs)を包含する15マーペプチドのプールでの一晩のin vitro再刺激の後、IL-2および/またはIFN-ガンマを産生するCS C末端、CS N末端およびHBs特異的CD4およびCD8 T細胞応答をフローサイトメトリーにより測定した。また、陰性対照として、幾つかの細胞を培地内でin vitroで一晩培養した(未刺激)。該ペプチド刺激細胞によりもたらされた平均サイトカイン応答から、未刺激細胞によりもたらされた平均サイトカイン応答を差し引くことにより、該Ag特異的応答を計算した。 Peripheral blood was collected, pooled and sequenced on day 21 (7d pII), 35 (7d pIII), 49 (21d pIII), 63 (35d pIII), 77 (49d pIII) after immunization CS C-terminus, CS N-terminus producing IL-2 and / or IFN-gamma after overnight in vitro restimulation with a pool of 15-mer peptides including (CS N-terminus, CS C-terminus or HBs) And HBs-specific CD4 and CD8 T cell responses were measured by flow cytometry. As a negative control, some cells were cultured overnight in culture medium (unstimulated). The Ag-specific response was calculated by subtracting the average cytokine response produced by unstimulated cells from the average cytokine response produced by the peptide stimulated cells.
結果は以下のことを示している。すなわち、
・APP、PPAおよびCCCは全て、N末端特異的CD8 T細胞応答を誘導する。7d pIIの時点で、N末端特異的CD8 T細胞応答がCCC群において観察され、ついでAPP群において観察される。試験したその後の時点(21d pIII、35d pIIIおよび49d pIII)では、これらの応答はAPP、PPAおよびCCC群において類似した強度であり、そのように維持されている(図25)。
・APP、PPAおよびCCCは、3回目の免疫化の49日後に尚も持続するC末端特異的CD8 T細胞応答を誘導する(図26)。
・N末端特異的CD4 T細胞応答は、主として、APPまたはCCCで免疫化されたマウスにおいて検出されており、APP群において、より高い強度のそのような応答が検出されている(図27)。
・全ての群がC末端特異的CD4 T細胞応答を示している。しかし、APPおよびCCCは、より高いレベルの類似したC末端特異的CD4 T細胞応答を誘導し、これらは、試験した全ての時点においてPPAおよびPPPにより誘導されたものより約2〜3倍高い(図28)。
・HBs特異的CD4およびCD8 T細胞応答は、PPPで免疫化された動物においては、APP、PPAまたはCCCで免疫化された場合より高い(図29および30)。
・CCC治療計画は、CSおよびHBs特異的CD4およびCD8 T細胞応答の同時誘導を伴う唯一ものである。
The results show that: That is,
APP, PPA and CCC all induce an N-terminal specific CD8 T cell response. At 7d pII, an N-terminal specific CD8 T cell response is observed in the CCC group and then in the APP group. At subsequent time points tested (21d pIII, 35d pIII and 49d pIII), these responses are of similar intensity in the APP, PPA and CCC groups and are maintained as such (FIG. 25).
APP, PPA and CCC induce a C-terminal specific CD8 T cell response that still persists 49 days after the third immunization (FIG. 26).
• N-terminal specific CD4 T cell responses have been detected primarily in mice immunized with APP or CCC, with higher intensity such responses detected in the APP group (FIG. 27).
• All groups show C-terminal specific CD4 T cell responses. However, APP and CCC induce higher levels of similar C-terminal specific CD4 T cell responses, which are about 2-3 times higher than those induced by PPA and PPP at all time points tested ( Figure 28).
HBs-specific CD4 and CD8 T cell responses are higher in animals immunized with PPP than when immunized with APP, PPA or CCC (FIGS. 29 and 30).
• The CCC treatment plan is the only one with simultaneous induction of CS and HBs-specific CD4 and CD8 T cell responses.
CSおよびHBs特異的CD4およびCD8 T細胞応答のサイトカインプロファイルも測定され、試験された全ての時点にわたって類似していた。試験された全ての時点の代表例として、21d pIII時点からのものを以下に示す(図31〜36)。簡潔に説明すると、Ag特異的CD8 T細胞応答は、大部分は、IFNgを産生するCD8 T細胞によりもたらされる(図31〜33)。これとは対照的に、Ag特異的CD4 T細胞応答は、IFNg、IFNgおよびIL-2を産生するCD4 T細胞と、それより低い度合ではあるが、IL-2を産生するCD4 T細胞との混合物によりもたらされる(図34〜36)。 Cytokine profiles of CS and HBs-specific CD4 and CD8 T cell responses were also measured and were similar across all time points tested. Representative examples of all time points tested are shown below from the 21d pIII time points (FIGS. 31-36). Briefly, the Ag-specific CD8 T cell response is largely mediated by CD8 T cells that produce IFNg (FIGS. 31-33). In contrast, the Ag-specific CD4 T cell response is between CD4 T cells producing IFNg, IFNg and IL-2, and to a lesser extent, CD4 T cells producing IL-2. Provided by the mixture (Figures 34-36).
また、3回目の免疫化の14日後および42日後に集めた血清に関してELISAによりAg特異的抗体応答を測定した。特に、R32LRポリペプチド(すなわち、P. falciparum CSPの中央部分を包含するもの)に対する及びHBsに対する全Ig応答を測定した(Mettensら, Vaccine 2008)。全ての免疫化法が、試験した最終時点(すなわち、3回目の免疫化の42日後)まで持続するR32LRおよびHBs特異的抗体応答を惹起した(図37および38)。 In addition, Ag-specific antibody responses were measured by ELISA on sera collected 14 and 42 days after the third immunization. In particular, total Ig responses to R32LR polypeptides (ie, including the central portion of P. falciparum CSP) and to HBs were measured (Mettens et al., Vaccine 2008). All immunization methods elicited R32LR and HBs specific antibody responses that persisted until the last time point tested (ie, 42 days after the third immunization) (FIGS. 37 and 38).
CB6F1マウスにおけるC7-Ade2+RTS,S/AS01Bの共配合の免疫原性
この実験においては、RTS,S/AS01Bと共配合(Combo)されたチンプアデノウイルス(chimpadenovirus)C7 Ade2の免疫原性を比較した。特に、CB6F1マウスにおけるコンボの1回、2回または3回の注射により惹起される免疫応答を比較した(4プールのマウス/群)。また、該コンボの2回の注射の間の異なる間隔(すなわち、14日および21日)を評価した。最後に、A-P-Pで免疫化されたマウスの群を該実験における対照として用いた。実験計画を以下にまとめる。
ここで、
A =10e9 vpのC7 Ade2、
P = 5μg RTS,S/50μl AS01B、
C = 10e10 vpのC7 Ade2+ 5μg RTS,S/50μl AS01B。
here,
A = C7 Ade2 with 10e9 vp,
P = 5μg RTS, S / 50μl AS01B,
C = 10e10 vp C7 Ade2 + 5 μg RTS, S / 50 μl AS01B.
第35日、42日、49日、63日および98日に末梢血を集め、プールし、対象の配列(すなわち、CSタンパク質のN末端領域(N末端)、C末端領域(C末端)またはHBs)を包含する15マーペプチドのプールでの一晩のin vitro再刺激の後、IL-2および/またはIFN-ガンマを産生するCS C末端、CS N末端およびHBs特異的CD4およびCD8 T細胞応答をフローサイトメトリーにより測定した。また、陰性対照として、幾つかの細胞を培地内でin vitroで一晩培養した(未刺激)。該ペプチド刺激細胞によりもたらされた平均サイトカイン応答から、未刺激細胞によりもたらされた平均サイトカイン応答を差し引くことにより、該Ag特異的応答を計算した。
Peripheral blood is collected and pooled on
結果は、これらの実験条件下、CS C末端、CS N末端およびHBs特異的CD4およびCD8 T細胞応答を同時に誘導するためにはコンボの2回の免疫化が必要であったことを示している。コンボの2回の免疫化の間の間隔は、検出されるAg特異的T細胞応答のレベルに有意な影響を及ぼさないようであった。コンボで3回免疫化されたマウスにおいては、より高いHBs特異的CD4およびCD8 T細胞応答の傾向が認められた。Ag特異的T細胞応答の速度論を図39〜44に示す。 The results show that under these experimental conditions, two immunizations of the combo were required to simultaneously induce CS C-terminal, CS N-terminal and HBs-specific CD4 and CD8 T cell responses . The interval between the two immunizations of the combo did not appear to have a significant effect on the level of Ag-specific T cell response detected. In mice immunized 3 times with the combo, there was a trend towards higher HBs-specific CD4 and CD8 T cell responses. The kinetics of Ag-specific T cell responses are shown in Figures 39-44.
CSおよびHBs特異的CD4およびCD8 T細胞応答のサイトカインプロファイルも測定し、試験された全ての時点にわたって類似していた。試験された全ての時点の代表例として、該研究の第42日からのものを以下に示す(図42〜47)。簡潔に説明すると、Ag特異的CD8 T細胞応答は、大部分は、IFNgを産生するCD8 T細胞によりもたらされる(図45〜47)。これとは対照的に、Ag特異的CD4 T細胞応答は、IFNg、IFNgおよびIL-2を産生するCD4 T細胞と、それより低い度合ではあるが、IL-2を産生するCD4 T細胞との混合物によりもたらされる(図48〜50)。 Cytokine profiles of CS and HBs-specific CD4 and CD8 T cell responses were also measured and were similar across all time points tested. Representative examples from all time points tested are shown below from day 42 of the study (FIGS. 42-47). Briefly, the Ag-specific CD8 T cell response is largely mediated by CD8 T cells that produce IFNg (FIGS. 45-47). In contrast, the Ag-specific CD4 T cell response is between CD4 T cells producing IFNg, IFNg and IL-2, and to a lesser extent, CD4 T cells producing IL-2. Provided by the mixture (Figures 48-50).
また、該研究の第56日および第99日に集めた血清に関してELISAによりAg特異的抗体応答を測定した。特に、R32LRポリペプチド(すなわち、P. falciparum CSPの中央部分を包含するもの)に対する及びHBsに対する全Ig応答を測定した(Mettensら, Vaccine 2008)。全ての免疫化法が、試験した最終時点(すなわち、3回目の免疫化の42日後)まで持続するR32LRおよびHBs特異的抗体応答を惹起した。各群内で、これらの応答は、試験した両方の時点において同様の強度であった。また、コンボで免疫化した群を比較したところ、3用量のコンボで免疫化された群において、より高い応答の傾向が認められた(図51および52)。
Ag-specific antibody responses were also measured by ELISA on sera collected on
CB6F1マウスにおいてCSおよびHBs T細胞およびAb応答を同時に惹起するための、コンボの各成分(すなわち、C7-Ade2、RTS,SおよびAS01B)の必要性の評価
この実験においては、CSおよびHBs特異的CD4およびCD8 T細胞応答を同時に惹起するための、コンボの各成分(すなわち、C7-Ade2、RTS,SおよびAS01B)の必要性を評価した。この実験においては、以下に示すとおり、コンボまたはその成分でCB6F1マウス(6プールのマウス/群)を筋肉内に2回(第0日および第14日)免疫化した。
ここで、
A =10e9 vpのC7 Ade2、
P = 5μg RTS,S/50μl AS01B、
C =10e9 vpのC7 Ade2 + 5μg RTS,S/50μl AS01B、
C7空 =10e9 vpのC7空ベクター(インサート無し)、
RTS,S = 5μgのRTS,S、
AS01B = GSK所有権所有アジュバント系1B、
バッファー= AS01Bバッファー(免疫刺激物質無し)。
here,
A = C7 Ade2 with 10e9 vp,
P = 5μg RTS, S / 50μl AS01B,
C = 10e9 vp C7 Ade2 + 5μg RTS, S / 50μl AS01B,
C7 empty = 10e9 vp C7 empty vector (no insert),
RTS, S = 5 μg RTS, S,
AS01B = GSK proprietary adjuvant system 1B,
Buffer = AS01B buffer (no immunostimulant).
第14日、28日、70日、91日および112日に末梢血を集め、プールし、対象の配列(すなわち、CSタンパク質のN末端領域(N末端)、C末端領域(C末端)またはHBs)を包含する15マーペプチドのプールでの一晩のin vitro再刺激の後、IL-2および/またはIFN-ガンマを産生するCS C末端、CS N末端およびHBs特異的CD4およびCD8 T細胞応答をフローサイトメトリーにより測定した。また、陰性対照として、幾つかの細胞を培地内でin vitroで一晩培養した(未刺激)。該ペプチド刺激細胞によりもたらされた平均サイトカイン応答から、未刺激細胞によりもたらされた平均サイトカイン応答を差し引くことにより、該Ag特異的応答を計算した。
Peripheral blood is collected and pooled on
また、該研究の第42日および第84日に集めた血清に関してELISAによりAg特異的抗体応答を測定した。特に、R32LRポリペプチド(すなわち、P. falciparum CSPの中央部分を包含するもの)に対する及びHBsに対する全Ig応答を測定した(Mettensら, Vaccine 2008)。 Ag-specific antibody responses were also measured by ELISA on sera collected on days 42 and 84 of the study. In particular, total Ig responses to R32LR polypeptides (ie, including the central portion of P. falciparum CSP) and to HBs were measured (Mettens et al., Vaccine 2008).
結果は、CS(N末端およびC末端)CD4およびCD8 T細胞応答(図53〜58)ならびにR32LRおよびHBs抗体応答(図65および66)を同時に惹起するためにはコンボの各成分が必要であることを示している。 The results show that each component of the combo is required to elicit CS (N-terminal and C-terminal) CD4 and CD8 T cell responses (Figures 53-58) and R32LR and HBs antibody responses (Figures 65 and 66) simultaneously It is shown that.
CSおよびHBs特異的CD4およびCD8 T細胞応答の平均サイトカインプロファイルをも該研究の各時点で測定した。これらを図59〜64に示す。簡潔に説明すると、Ag特異的CD8 T細胞応答は、大部分は、IFNgを産生するCD8 T細胞によりもたらされる(図59〜61)。これとは対照的に、Ag特異的CD4 T細胞応答は、IFNg、IFNgおよびIL-2を産生するCD4 T細胞と、それより低い度合ではあるが、IL-2を産生するCD4 T細胞との混合物によりもたらされる(図62〜64)。 Mean cytokine profiles of CS and HBs specific CD4 and CD8 T cell responses were also measured at each time point in the study. These are shown in FIGS. Briefly, Ag-specific CD8 T cell responses are largely mediated by CD8 T cells that produce IFNg (FIGS. 59-61). In contrast, the Ag-specific CD4 T cell response is between CD4 T cells producing IFNg, IFNg and IL-2, and to a lesser extent, CD4 T cells producing IL-2. Provided by the mixture (Figures 62-64).
CB6F1マウスにおける合成C7 Ade2の免疫原性
Ade2構築物を発現する合成C7チンプアデノウイルス(chimpadenovirus)を利用可能にし、マウスにおけるその免疫原性を元のC7 Ade2の場合と比較した。この実験においては、CB6F1マウス(6プールのマウス/群)を10e9 vpの元のC7 Ade2またはその合成対応体で免疫化した。免疫化の21日後、28日後および35日後に末梢血を集め、プールし、対象の配列(すなわち、CSタンパク質のN末端領域(N末端)、C末端領域(C末端))を包含する15マーペプチドのプールでの一晩のin vitro再刺激の後、IL-2および/またはIFN-ガンマを産生するCS C末端およびCS N末端CD4およびCD8 T細胞応答をフローサイトメトリーにより測定した。また、陰性対照として、幾つかの細胞を培地内でin vitroで一晩培養した(未刺激)。該ペプチド刺激細胞によりもたらされた平均サイトカイン応答から、未刺激細胞によりもたらされた平均サイトカイン応答を差し引くことにより、該Ag特異的応答を計算した。
Immunogenicity of synthetic C7 Ade2 in CB6F1 mice
A synthetic C7 chimpadenovirus expressing the Ade2 construct was made available and its immunogenicity in mice compared to that of the original C7 Ade2. In this experiment, CB6F1 mice (6 pool mice / group) were immunized with 10e9 vp original C7 Ade2 or its synthetic counterpart. Peripheral blood collected 21 days, 28 days and 35 days after immunization, pooled, and 15-mer containing the sequence of interest (ie, N-terminal region (N-terminal), C-terminal region (C-terminal) of CS protein) After overnight in vitro restimulation with a pool of peptides, CSC-terminal and CSN-terminal CD4 and CD8 T cell responses producing IL-2 and / or IFN-gamma were measured by flow cytometry. As a negative control, some cells were cultured overnight in culture medium (unstimulated). The Ag-specific response was calculated by subtracting the average cytokine response produced by unstimulated cells from the average cytokine response produced by the peptide stimulated cells.
結果は、両方のC7アデノウイルスが同レベルのN末端およびC末端特異的CD4およびCD8 T細胞応答を惹起したことを示している。特に、使用したウイルスベクター(元のまたは合成の)に無関係に、Ag特異的CD8 T細胞応答は主としてN末端特異的であった(図67および68)。N末端およびC末端特異的CD4 T細胞応答は、使用したウイルスベクターに無関係に、同様の強度であった(図69および70)。CS特異的CD4およびCD8 T細胞応答の平均サイトカインプロファイルをも測定し、これらを図71〜74に示す。簡潔に説明すると、Ag特異的CD8 T細胞応答は、大部分は、IFNgを産生するCD8 T細胞によりもたらされる(図71〜72)。これとは対照的に、Ag特異的CD4 T細胞応答は、IFNg、IFNgおよびIL-2を産生するCD4 T細胞と、それより低い度合ではあるが、IL-2を産生するCD4 T細胞との混合物によりもたらされる(図73〜74)。 The results show that both C7 adenoviruses elicited similar levels of N-terminal and C-terminal specific CD4 and CD8 T cell responses. In particular, regardless of the viral vector used (original or synthetic), the Ag-specific CD8 T cell response was primarily N-terminal specific (FIGS. 67 and 68). N-terminal and C-terminal specific CD4 T cell responses were of similar intensity regardless of the viral vector used (FIGS. 69 and 70). Average cytokine profiles of CS-specific CD4 and CD8 T cell responses were also measured and are shown in FIGS. Briefly, the Ag-specific CD8 T cell response is largely mediated by CD8 T cells that produce IFNg (FIGS. 71-72). In contrast, the Ag-specific CD4 T cell response is between CD4 T cells producing IFNg, IFNg and IL-2, and to a lesser extent, CD4 T cells producing IL-2. Provided by the mixture (Figures 73-74).
配列番号1
アミノ酸配列
ヌクレオチド配列
チンプアデノ 7(WO 03/046124の配列番号17)
チンプアデノ 7(WO 03/046124の配列番号20)
配列番号3のタンパク質の発現カセットの配列。タンパク質コード領域は下線で示されている。
Ade2発現カセットの配列を以下に示す。タンパク質コード領域は下線で示されている。
Amino acid sequence
Nucleotide sequence
Thimpadeno 7 (SEQ ID NO: 17 of WO 03/046124)
Thimpadeno 7 (SEQ ID NO: 20 of WO 03/046124)
The sequence of the expression cassette of the protein of SEQ ID NO: 3. The protein coding region is underlined.
The sequence of the Ade2 expression cassette is shown below. The protein coding region is underlined.
Claims (26)
a.適当な細胞系上で該ベクターを増殖させ、
b.該ベクターを回収する工程を含んでなる、製造方法。 A method for producing a viral vector according to any one of claims 1 to 5,
a. Propagating the vector on a suitable cell line,
b. A production method comprising a step of recovering the vector.
b.請求項6記載の組成物、または
c.請求項7〜10もしくは12〜20のいずれか1項記載のワクチン
の治療的有効量を投与することを含んでなる治療方法。 a. The viral vector according to any one of claims 1 to 5,
b. The composition of claim 6, or
c. 21. A method of treatment comprising administering a therapeutically effective amount of the vaccine of any one of claims 7-10 or 12-20.
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US99280207P | 2007-12-06 | 2007-12-06 | |
| PCT/EP2008/066762 WO2009071613A2 (en) | 2007-12-06 | 2008-12-04 | Vaccine |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2011505796A true JP2011505796A (en) | 2011-03-03 |
Family
ID=40591828
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2010536451A Pending JP2011505796A (en) | 2007-12-06 | 2008-12-04 | vaccine |
Country Status (20)
| Country | Link |
|---|---|
| EP (1) | EP2227550A2 (en) |
| JP (1) | JP2011505796A (en) |
| KR (1) | KR20100108544A (en) |
| CN (1) | CN101939438A (en) |
| AR (1) | AR069568A1 (en) |
| AU (1) | AU2008333208A1 (en) |
| BR (1) | BRPI0819889A2 (en) |
| CA (1) | CA2707245A1 (en) |
| CL (1) | CL2008003614A1 (en) |
| CO (1) | CO6300795A2 (en) |
| CR (1) | CR11537A (en) |
| DO (1) | DOP2010000164A (en) |
| IL (1) | IL205953A0 (en) |
| MA (1) | MA32003B1 (en) |
| MX (1) | MX2010006207A (en) |
| PE (1) | PE20091106A1 (en) |
| TW (1) | TW200938633A (en) |
| UY (1) | UY31510A1 (en) |
| WO (1) | WO2009071613A2 (en) |
| ZA (1) | ZA201003851B (en) |
Families Citing this family (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| GB0815872D0 (en) * | 2008-09-01 | 2008-10-08 | Pasteur Institut | Novel method and compositions |
| GB201016471D0 (en) * | 2010-09-30 | 2010-11-17 | Isis Innovation | Viral vector immunogenic compositions |
| MX350795B (en) * | 2011-04-08 | 2017-09-19 | Inmune Design Corp | Immunogenic compositions and methods of using the compositions for inducing humoral and cellular immune responses. |
| CN110687289B (en) * | 2019-10-17 | 2023-04-18 | 中国人民解放军陆军军医大学 | Application of FGL2 protein as malaria infection marker |
Citations (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2004055187A1 (en) * | 2002-12-17 | 2004-07-01 | Crucell Holland B.V. | Recombinant viral-based malaria vaccines |
| JP2006512405A (en) * | 2002-10-23 | 2006-04-13 | グラクソスミスクライン・バイオロジカルス・ソシエテ・アノニム | How to vaccinate against malaria |
| WO2006040334A1 (en) * | 2004-10-14 | 2006-04-20 | Crucell Holland B.V. | Malaria prime/boost vaccines |
| WO2007003384A1 (en) * | 2005-06-30 | 2007-01-11 | Glaxosmithkline Biologicals Sa | Anti-malaria vaccine |
| WO2007110409A1 (en) * | 2006-03-27 | 2007-10-04 | Crucell Holland B.V. | Compositions comprising a recombinant adenovirus and an adjuvant |
| WO2008107370A1 (en) * | 2007-03-02 | 2008-09-12 | Glaxosmithkline Biologicals S.A. | Novel method and compositions |
Family Cites Families (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US20040136963A1 (en) * | 2001-06-22 | 2004-07-15 | The Trustees Of The University Of Pennsylvania | Simian adenovirus vectors and methods of use |
| WO2005063805A1 (en) * | 2003-12-23 | 2005-07-14 | The Government Of The United States Of America, As Represented By The Secretary, Department Of Health And Human Services | Antibodies against the amino terminus region of circumsporozoite protein prevent the onset of malaria infection |
| CA2880060C (en) * | 2004-01-23 | 2018-03-13 | Agostino Cirillo | Chimpanzee adenovirus vaccine carriers |
| WO2008009652A2 (en) * | 2006-07-18 | 2008-01-24 | Glaxosmithkline Biologicals S.A. | Vaccines for malaria |
| GB0706914D0 (en) * | 2007-04-10 | 2007-05-16 | Isis Innovation | Novel adenovirus vectors |
-
2008
- 2008-12-04 KR KR1020107014859A patent/KR20100108544A/en not_active Withdrawn
- 2008-12-04 WO PCT/EP2008/066762 patent/WO2009071613A2/en not_active Ceased
- 2008-12-04 CA CA2707245A patent/CA2707245A1/en not_active Abandoned
- 2008-12-04 TW TW097147198A patent/TW200938633A/en unknown
- 2008-12-04 CN CN2008801267101A patent/CN101939438A/en active Pending
- 2008-12-04 PE PE2008002018A patent/PE20091106A1/en not_active Application Discontinuation
- 2008-12-04 MX MX2010006207A patent/MX2010006207A/en not_active Application Discontinuation
- 2008-12-04 AR ARP080105281A patent/AR069568A1/en not_active Application Discontinuation
- 2008-12-04 JP JP2010536451A patent/JP2011505796A/en active Pending
- 2008-12-04 CL CL2008003614A patent/CL2008003614A1/en unknown
- 2008-12-04 AU AU2008333208A patent/AU2008333208A1/en not_active Abandoned
- 2008-12-04 BR BRPI0819889-6A patent/BRPI0819889A2/en not_active IP Right Cessation
- 2008-12-04 EP EP08857977A patent/EP2227550A2/en not_active Withdrawn
- 2008-12-04 UY UY31510A patent/UY31510A1/en unknown
-
2010
- 2010-05-25 IL IL205953A patent/IL205953A0/en unknown
- 2010-05-28 ZA ZA2010/03851A patent/ZA201003851B/en unknown
- 2010-06-02 DO DO2010000164A patent/DOP2010000164A/en unknown
- 2010-06-10 CO CO10070274A patent/CO6300795A2/en not_active Application Discontinuation
- 2010-06-29 CR CR11537A patent/CR11537A/en not_active Application Discontinuation
- 2010-07-05 MA MA32991A patent/MA32003B1/en unknown
Patent Citations (6)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2006512405A (en) * | 2002-10-23 | 2006-04-13 | グラクソスミスクライン・バイオロジカルス・ソシエテ・アノニム | How to vaccinate against malaria |
| WO2004055187A1 (en) * | 2002-12-17 | 2004-07-01 | Crucell Holland B.V. | Recombinant viral-based malaria vaccines |
| WO2006040334A1 (en) * | 2004-10-14 | 2006-04-20 | Crucell Holland B.V. | Malaria prime/boost vaccines |
| WO2007003384A1 (en) * | 2005-06-30 | 2007-01-11 | Glaxosmithkline Biologicals Sa | Anti-malaria vaccine |
| WO2007110409A1 (en) * | 2006-03-27 | 2007-10-04 | Crucell Holland B.V. | Compositions comprising a recombinant adenovirus and an adjuvant |
| WO2008107370A1 (en) * | 2007-03-02 | 2008-09-12 | Glaxosmithkline Biologicals S.A. | Novel method and compositions |
Non-Patent Citations (1)
| Title |
|---|
| JPN6013046280; Science Vol.225, 1984, p.593-599 * |
Also Published As
| Publication number | Publication date |
|---|---|
| BRPI0819889A2 (en) | 2015-06-16 |
| PE20091106A1 (en) | 2009-08-24 |
| TW200938633A (en) | 2009-09-16 |
| MX2010006207A (en) | 2010-10-04 |
| IL205953A0 (en) | 2010-11-30 |
| CA2707245A1 (en) | 2009-06-11 |
| WO2009071613A3 (en) | 2009-08-13 |
| KR20100108544A (en) | 2010-10-07 |
| CR11537A (en) | 2010-08-18 |
| MA32003B1 (en) | 2011-01-03 |
| EP2227550A2 (en) | 2010-09-15 |
| AR069568A1 (en) | 2010-02-03 |
| CN101939438A (en) | 2011-01-05 |
| AU2008333208A1 (en) | 2009-06-11 |
| WO2009071613A2 (en) | 2009-06-11 |
| UY31510A1 (en) | 2009-08-03 |
| CL2008003614A1 (en) | 2010-01-15 |
| DOP2010000164A (en) | 2010-07-31 |
| CO6300795A2 (en) | 2011-07-21 |
| ZA201003851B (en) | 2012-11-28 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20100272786A1 (en) | Vaccine | |
| JP5592110B2 (en) | Malaria vaccine | |
| US20100272745A1 (en) | Vaccines for malaria | |
| JP2011505796A (en) | vaccine | |
| US8999347B2 (en) | Vaccines for malaria | |
| CN101553249A (en) | Vaccines for malaria |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20111202 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20130917 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20131210 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20131217 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20140603 |