Schizosaccharomyces pombe
| Schizosaccharomyces pombe | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| |||||||||||||||||
| Clasificación científica | |||||||||||||||||
| |||||||||||||||||
| Nome binomial | |||||||||||||||||
| Schizosaccharomyces pombe Lindner (1893) | |||||||||||||||||
Schizosaccharomyces pombe, tamén chamado "lévedo de fisión", é unha especie de lévedo utilizado na fabricación de certas cervexas tradicionais e como organismo modelo en investigacións de bioloxía molecular e bioloxía celular. É un eucariota do reino dos fungos formado por células cilíndricas con forma de bastón (parecen bacilos, pero máis grandes). As células miden xeralmente de 3 a 4 micrómetros de diámetro e de 7 a 14 micrómetros de lonxitude. O seu xenoma, que é de aproximadamente 14,1 millóns de pares de bases, estímase que contén uns 4.970 xenes codificantes de proteínas e polo menos 450 ARNs non codificantes.[1]
Estas células manteñen a súa forma ao creceren exclusivamente polos extremos da célula e dividirse por fisión binaria medial para producir dúas células fillas de igual tamaño, o que o fai un poderoso modelo para estudar o ciclo celular.
O lévedo de fisión foi illado en 1893 por Paul Lindner da cervexa de millo miúdo producida no leste de África. O nome da especie pombe é unha palabra da lingua suahili que significa cervexa. Na década de 1950 empezou a utilizarse como modelo experimental por Urs Leupold para estudos de xenética,[2][3] e por Murdoch Mitchison para estudar o ciclo celular.[4][5] O investigador do lévedo de fisión Paul Nurse conseguiu unir as dúas escolas ata entón independentes dos estudos xenéticos e do ciclo celular. Xunto con Lee Hartwell e Tim Hunt, Nurse gañou o Premio Nobel de Medicina de 2001 polos seus traballos sobre a regulación dio ciclo celular.
A secuencia do xenoma de S. pombe foi publicada en 2002 por un consorcio liderado polo Sanger Institute, converténdose no sexto modelo de organismo eucariótico cuxo xenoma foi secuenciado completamente. Isto fixo del un poderoso organismo modelo, xa que se identificaron nel moitos xenes homólogos de enfermidades humanas. En 2006, foi publicada a localización subcelular de case todas as proteínas de S. pombe utilizando a proteína fluorescente verde como etiqueta molecular.[6]
S. pombe é tamén un organismo importante para estudar as respostas celulares aos danos no ADN e o proceso da replicación do ADN.
Illáronse aproximadamente 160 cepas naturais de S. pombe. Foron recollidas en varias localizacións de diversas partes do mundo como Europa, Norte e Suramérica, e Asia. A maioría destas cepas recolléronse de froitas cultivadas como mazás e uvas, ou de diversas bebidas alcohólicas, como por exemplo a cachaza brasileira. S. pombe tamén está presente no té fermentado kombucha.[7] Porén, non está claro se en ditas bebidas S. pombe é o principal fermentador ou é máis ben un contaminante. A ecoloxía natural dos lévedos Schizosaccharomyces aínda non está ben estudada.
Historia
editarS. pombe descubriuse en 1893 cando un grupo que traballaba no Laboratorio da Asociación de Cervexeiros de Alemaña examinou uns pousos sedimentados en cervexa de millo miúdo importada de África Oriental, que lle daba á cervexa un sabor ácido e desagradable. O termo schizo, significa "división" ou "fisión", e xa fora usado previamente no nome Schizosaccharomycetes. A palabra pombe elixida para nomear a especie significa cervexa en suahili, e débese á procedencia desa cervexa. As cepas estándar de S. pombe foron illadas bastante máis tarde por Urs Leupold en 1946 e 1947 a partir dun cultivo que obtivo da colección de lévedos de Delft, nos Países Baixos. Fora depositada alí por A. Osterwalder co nome S. pombe var. liquefaciens, despois de que a illara en 1924 dun viño francés (moi probablemente picado) na Estación Experimental de Vinicultura e Horticultura de Wädenswil, Suíza. O cultivo utilizado por Urs Leupold contiña (entre outras) células cos tipos de apareamento h90 (cepa 968), h- (cepa 972), e h+ (cepa 975). Despois disto, fixéronse dous grandes esforzos para illar S. pombe de froitas, néctar, ou fermentacións: un feito por Florenzano et al.[8] nos viñedos do oeste de Sicilia, e o outro realizado por Gomes et al. (2002) en catro rexións do sueste do Brasil.[9]
Ecoloxía
editarO lévedo de fisión S. pombe pertence ao filo Ascomycota, que comprende o grupo máis grande e diverso dos fungos. Os ascomicetos de vida libre atópanse comunmente en exsudados de árbores, raíces de plantas e no solo que as rodea, en froitos podres e maduros, e en asociación con insectos vectores que os transportan entre distintos substratos. Moitas destas asociacións son simbióticas ou saprofíticas, aínda que numerosos ascomicetos (e os seus parentes basidiomicetos) son importantes patóxenos que atacan a moitas especies de plantas, incluíndo plantas agrícolas comerciais. Entre os xéneros de lévedos ascomicetos, os lévedos de fisión Schizosaccharomyces son os únicos que depositan a-(1,3)- glucano ou pseudonixerano na parede celular xunto cos mellor coñecidos b-glucanos e presentan unha carencia virtual de quitina. As especies deste xénero tamén difiren na composición de mananos, que presentan azucres terminais de D-galactosa nas cadeas laterais dos seus mananos. S. pombe realiza a fermentación aerobia en presenza de exceso de azucre.[10] S. pombe pode degradar ácido L-málico, un dos ácidos orgánicos dominantes no viño, o que os diferencia doutras cepas de lévedos do xénero Saccharomyces.
Comparación co lévedo de xemación Saccharomyces cerevisiae
editarAs especies de lévedos Schizosaccharomyces pombe e Saccharomyces cerevisiae son ambas intensamente estudadas, e son importantes ferramentas na bioloxía molecular e celular. Algunhas características xenómicas e do ciclo celular que as diferencian son:
- S. cerevisiae ten aproximadamente 5.600 marcos de lectura abertos; S. pombe ten uns 4.970.
- Malia o seu número similar de xenes, S. cerevisiae ten só uns 250 intróns, mentres que S. pombe ten case 5.000.
- S. cerevisiae ten 16 cromosomas; S. pombe ten 3.
- S. cerevisiae é xeralmente diploide, mentres que S. pombe adoita a ser haploide.
- S. cerevisiae permanece na fase G1 do ciclo celular durante extensos períodos de tempo (en consecuencia, a transición G1-S está estritamente controlada), mentres que S. pombe permanece na fase G2 do ciclo celular durante un longo período de tempo (en consecuencia, a transición G2-M está baixo estrito control).
- Ambas as especies comparten xenes con eucariotas superiores que non comparten entre si. S. pombe ten xenes para a maquinaria de interferencia de ARN como os dos vertebrados, mentres que estes están ausentes en S. cerevisiae. S. cerevisiae tamén ten unha heterocromatina moi simplificada en comparación con S. pombe.[11] Inversamente, S. cerevisiae ten peroxisomas ben desenvolvidos, pero S. pombe non.
- S. cerevisiae ten centrómeros puntuais pequenos de 125 bp, e orixes de replicación definidas por secuencias de aproximadamente o mesmo tamaño. Ao contrario, S. pombe ten centrómeros grandes, repetitivos de (40–100 kb) máis similares aos de mamíferos, e orixes de replicación dexeneradas de polo menos 1 kb.
Ciclo vital
editar
O lévedo de fisión é un fungo unicelular cun xenoma simple e unha rápida taxa de crecemento. S. pombe ten forma de bastonciños de aproximadamente 3 µm de diámetro, e crece só por prolongación dos seus extremos. Despois da mitose, ten lugar a división pola formación dun septo ou placa celular que divide a célula no seu punto medio.
Os procesos principais da reprodución celular son a duplicación dos cromosomas, que ten lugar na fase S da interfase, seguida da segregación dos cromosomas e a división do núcleo (mitose) e a división do citoplasma (citocinese), que en conxunto forman a fase M (mitótica) do ciclo celular. G1 é o período entre as fases M e S, e G2 é o período entre as fases S e M. S. pombe permanece na fase G2 do ciclo celular durante un extenso período de tempo (a diferenza de Saccharomyces cerevisiae, que permanece moito tempo na fase G1).
A mitose dos lévedos de fisión está controlada por mecanismos similares aos dos organismos multicelulares animais. Normalmente prolifera nun estado haploide. Cando ten escaseza de alimentos, as células de tipos de apareamento opostos (P e M) fusiónanse formando un cigoto diploide, que inmediatamente entra en meiose, xerando catro esporas haploides nun asco. Cando melloran as condicións no seu medio, estas esporas xerminan e producen células haploides proliferantes.[12]
-
 Características xerais do ciclo vital.
Características xerais do ciclo vital. -
 O ciclo celular dun lévedo de fisión.
O ciclo celular dun lévedo de fisión. -
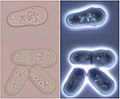 Estadios da división de Schizosaccharomyces vistos con microscopia óptica de campo escuro e brillante.
Estadios da división de Schizosaccharomyces vistos con microscopia óptica de campo escuro e brillante.


Citocinese no lévedo de fisión
editar
As características xerais da citocinese móstranse aquí. O sitio onde se vai producir a división celular determínase antes da anafase. O fuso da anafase (en verde na fuigura) colócase despois de modo que os cromosomas segregados estean en lados opostos do plano de clivaxe predeterminado.
Control do tamaño no lévedo de fisión
editar
No lévedo de fisión, no cal o crecemento da célula controla a progresión da fase G2 á M, unha mutación chamada wee1 causa a entrada en mitose e un tamaño anormalmente pequeno, o que orixina unha fase G2 máis curta. A G1 está prolongada, o que suxire que a progresión desde o comezo do ciclo responde ao crecemento cando se perde o control G2/M. Ademais, as células que están en condicións pobres en nutrientes crecen lentamente e lévalles máis tempo duplicar o seu tamaño e dividirse. Os niveis baixos de nutrientes tamén cambian o limiar de crecemento da célula para que a célula avance polo ciclo celular con tamaños menores. Despois dunha exposición a condicións estresantes (calor a 40 °C ou axentes oxidantes como o peróxido de hidróxeno) as células de S. pombe sofren un envellecemento que se mide polo incremento do tempo de división celular e o aumento da probabilidade de morte celular.[13] Finalmente, as células coa mutación wee1 son menores que as de tipo silvestre, pero lévalles o dobre de tempo completar o ciclo celular. Isto é posible porque as células máis pequenas crecen máis lentamente, é dicir, a súa masa sumada total por unidade de tempo é menor que a das células normais.
Crese que un gradiente espacial coordina o tamaño celular e a entrada na mitose.[14][15][16] A proteína quinase Pom1 está localizada no córtex celular, e ten a súa maior concentración nos extremos da célula. Os reguladores do ciclo celular Cdr2, Cdr1 e Wee1 están presentes en nodos corticais da parte media da célula. Nas células pequenas, o gradiente Pom1 chega á maioría dos nodos corticais. Pom1 inhibe Cdr2, impedindo que Cdr2 e Cdr1 inhiban Wee1, e permitindo que Wee1 fosforile Cdk1, o que inactiva a actividade de quinase dependente de ciclina (CDK) e impide a entrada na mitose. En células longas, o gradiente de Pom1 non chega aos nodos corticais, e, por tanto, Cdr2 e Cdr1 permanecen activas nos nodos. Cdr2 e Cdr1 inhiben Wee1, impedindo a fosforilación de Cdk1, o que leva á activación de CDK e a entrada na mitose. (Hai ademais varios outros reguladores da actividade de CDK.)
Cambios de tipo de apareamento no lévedo de fisión
editarOs lévedos de fisión cambian os tipos de apareamento por un evento de recombinación acoplado á replicación, que ten lugar durante a fase S do ciclo celular. Os lévedos de fisión usan asimetría intrínseca no proceso de replicación do ADN para cambiar o tipo de apareamento; foi o primeiro sistema no que se viu que se requiría unha dirección da replicación para o cambio de tipo celular. Os estudos do sistema de cambio do tipo de apareamento levaron ao descubrimento e caracterización do sitio RTS1 de terminación da replicación específico de sitio, e a un sitio MPS1 de pausa de replicación específico de sitio MPS1, e tamén a un novo tipo de impronta cromosómica, que marca unha das cromátides irmás no locus de tipo de apareamento mat1. Ademais, os traballos sobre a rexión doante silenciada levaron a grandes avances na comprensión da formación e mantemento da heterocromatina.[17]
Respostas aos danos no ADN
editarS. pombe é un microorganismo sexual facultativo que pode aparearse cando os nutrientes son limitados.[18] A exposición de S. pombe ao peróxido de hidróxeno, que é un axente que causa estrés oxidativo, orixina danos oxidativos no ADN, o que induce fortemente o apareamento entre os lévedos de fisión e a formación de esporas meióticas.[19] Este descubrimento suxire que a meiose, e especialmente a recombinación meiótica, poden ser unha adaptación para reparar o ADN danado.[20][21] Un descubrimento que apoia esta idea é que as lesións que afectan a unha soa base do ADN do tipo dU:dG en S. pombe estimulan a recombinación meiótica.[22] Esta recombinación require a actuación da uracilo-ADN glicosilase, un encima que quita o uracilo do ADN e inicia a reparación de excisión de base. Baseándose neste descubrimento, propúxose que a reparación de excisión de base dunha base uracilo, ou dun sitio abásico, ou dunha rotura ou amosega dunha soa febra do ADN é suficiente para iniciar a recombinación en S. pombe.[22] Outros experimentos feitos con S. pombe indicaron que o procesamento defectuoso dos intermediarios da replicación do ADN, é dicir, os fragmentos de Okazaki, causan danos no ADN, como roturas ou amosegas dunha soa febra ou ocos (gaps), e que isto estimula a recombinación meiótica.[23]
O lévedo de fisión como sistema modelo
editarO lévedo de fisión converteuse nun notable sistema modelo para estudar procesos básicos da célula que poden utilizarse para comprender o funcionamento de organismos máis complexos como os mamíferos e especialmente o home.[24][25] Este eucariota unicelular non é patóxeno e é facilmente cultivable e manipulable no laboratorio.[26] O lévedo de fisión é un dos eucariotas secuenciados que contén un menor número de xenes no seu xenoma, e ten só tres cromosomas.[27] Moitos deses xenes que son responsables do control da división celular e da organización da célula no lévedo de fisión, teñen xenes homólogos no xenoma humano.[26][28][29] A regulación do ciclo celular e da división son cruciais para o crecemento e desenvolvemento das células. Os xenes conservados do lévedo de fisión foron moi estudados e isto influíu en moitos desenvolvementos biomédicos recentes.[30][31] O lévedo de fisión é tamén un sistema modelo práctico para observar a división celular, que se pode seguir facilmente polo microscopio.[26] Ademais, ten un tempo de xeración extremadamente curto de só 2 a 4 horas, o que facilita a súa observación e cultivo no laboratorio.[28] A simplicidade da estrutura xenómica do lévedo de fisión pero con semellanzas cos xenomas de mamíferos, a facilidade para manipulalo, e a capacidade de usalo para análises de fármacos son as principais razóns polas que se usa tanto en biomedicina, bioloxía celular e en análises xenéticas.[28][32][33][34]
Xenoma de S. pombe
editarS. pombe utilízase para o estudo da división e crecemento celular debido ás rexións xenómicas conservadas que tamén se encontran en humanos, como son: proteínas da heterocromatina, orixes de replicación grandes, centrómeros grandes, puntos de control celular conservados, función dos telómeros, splicing de xenes, e outros procesos celulares.[27][35][36] O xenoma de S. pombe foi completamente secuenciado en 2002, e foi o sexto xenoma eucariótico secuenciado como parte do Proxecto Xenoma. Descubríronse uns 4.979 xenes nos seus tres cromosomas, que contiñan uns 14 Mb de ADN, con ocos nas rexións centroméricas (40kb) e teloméricas (260kb).[27] Despois da secuenciación inicial do seu xenoma, secuenciáronse outras rexións previamente non secuenciadas. A análise estrutural e funcional de todas estas rexións xénicas pode atoparse en bases de datos como PomBase.[37]
O lévedo de fisión non ten tantos xenes duplicados coma o lévedo de xemación, xa que estes supoñen só o 5%, o que fai deste organismo un gran modelo xenómico para investigar. O 40% dos xenes contiñan intróns, o que é un número elevado e dá a oportunidade de incrementar o número de tipos de proteínas producidos por medio de empalme alternativo.[27] Secuenciouse tamén o 81% dos seus tres centrómeros. As lonxitudes dos tres centrómeros son 34, 65, e 110 kb. Isto é de 300 a 100 veces meirandes que os do lévedo de xemación. Tamén se observou un extraordinariamente alto nivel de conservación (97%) nunha rexión de 1.780 bp nas rexións DGS do centrómero. Este alongamento dos centrómeros e as súas secuencias conservadas fano moi axeitado para o estudo da división celular, polo seu parecido coa humana.[27][38][39]
Durante o proxecto xenómico de S. pombe atopáronse 50 xenes que codifican proteínas ou mutacións que están ligadas con doenzas humanas, e case a metade (23) están relacionados co cancro.[27] Por tanto, estase usando para estudar xenes humanos, vías metabólicas de doenzas, a división celular e sistemas de comprobacion ou puntos de control (checkpoints) do ADN.[39][40][41][42]
Análise do ciclo celular
editarA replicación do ADN neste lévedo está sendo crecementemente estudada. Unha maior comprensión da replicación do ADN, expresión xénica e mecanismos conservados neste lévedo pode proporcionar información sobre eses procesos en mamíferos e no home.[36][43][44][45] Outros procesos, como o crecemento celular e o envellecemento son tamén estudados para comprender a partir deles outros sistemas máis complexos.[29][46][47][48]
A citocinese é un dos compoñentes da división celular que tamén se estuda no lévedo de fisión. Nel obsérvanse compoñentes ben conservados da citocinese que nos permiten estudar mutacións puntuais noutros escenarios xenómicos.[41][49][50][50][51] A citocinese é unha etapa crucial para a célula.[52] A formación do anel contráctil durante a citocinese está sendo moi estudada en S. pombe, xa que está moi conservado con respecto aos humanos.[41] As mutacións na citocinese poden orixinar un mal funcionamento da célula ou mesmo a morte celular e o desenvolvemento de células cancerosas.[41] Este é un proceso complexo na división das células humanas, pero é máis doado de estudar en S. pombe.
Unha das precaucións que toma a célula para asegurarse de que está tendo lugar unha división celular correcta é a existencia dunha serie de puntos de control ou checkpoints,[53][54] os cales aseguran que os mutantes serán eliminados.[55] Isto faise xeralmente por sinais que estimulan a ubiquitinación das dianas e atrasan a citocinese.[27] Sen estes puntos de control mitóticos, créanse e replícanse os mutantes, o que dá lugar a morte celular ou xeración de tumores. Paul Nurse, Leland Hartwell, e Tim Hunt recibiron o Premio Nobel de Fisioloxía e Medicina de 2001 polos seus descubrimentos de puntos de control claves conservados esenciais para que as células se dividan correctamente. Isto está ligado co cancro e células enfermas e foi moi importante en biomedicina.[56]
Tamén se estudou a dinámica de orgánulos e as respostas e posibles correlacións entre as células do lévedo e as de mamíferos.[57][58] As enfermidades mitocondriais e varios sistemas de orgánulos como o aparato de Golgi e o retículo endoplasmático poden comprenderse mellor estudando neste organismo modelo a dinámica dos cromosomas, os niveis de expresión de proteínas e a súa regulación.[42][44][59][60][60][61][62]
Ferramenta biomédica
editarMalia todo, hai tamén limitacións á hora de usar o lévedo de fisión como sistema modelo, debido á súa resistencia a múltiples fármacos. A resposta de resistencia a múltiples fármacos débese entre outras razóns á sobreexpresión de bombas de efluxo de drogas.[30] Paul Nurse e colegas crearon recentemente cepas de S. pombe sensibles a inhibidores químicos e a sondas de uso común para ver se é posible utilizar o lévedo de fisión como sistema modelo na investigación de fármacos.[30]
Por exemplo a doxorrubicina, un antibiótico quimioterapéutico moi común, ten moitos efectos adversos. Estanse buscando maneiras de comprender mellor como actúa a doxorrubicina observando os xenes ligados á resistencia usando o lévedo de fisión como sistema modelo. Atopátonse ligazóns entre os efectos adversos da doxorrubicina e o metabolismo de cromosomas e transporte de membrana. Os modelos metabólicos para a dirixir os fármacos ao punto desexado (drug targeting) están utilizándose agora en biotecnoloxía, e agárdanse máis avances no futuro usando como sistema modelo S. pombe.[31]
Enfoques experimentais
editarO lévedo de fisión é facilmente accesible, fácil de cultivar e manipular para crear mutantes, e poden manterse nun estado haploide ou diploide. Normalmente é unha célula haploide, pero posto en condicións de estrés, xeralmente deficiencia de nitróxeno, dúas células conxúganse para formar unha diploide que posteriormente forma catro esporas nun asco de tétrade.[63] Este proceso é doadamente visible e observable con microscopio e permite estudar o funcionamento da meiose.
Virtualmente pode aplicarse calquera experimento xenético ou técnica pode a este sistema modelo, como as seguintes: disección de tétrades, análise de mutáxenos, transformacións, e técnicas de microscopia como FRAT e FREP. Estánse usando tamén novos modelos como Tug-Of-War (gTOW) para analizar a robustez do lévedo e observar a súa expresión xenética. É doado preparar nel experimentos de knock-in e knockout de xenes, e como o xenoma do lévedo foi secuenciado esta tarefa é moi accesible e ben coñecida.[64][65]
Notas
editar- ↑ Wilhelm; et al. (2008). "Dynamic repertoire of a eukaryotic transcriptome surveyed at single-nucleotide resolution". Nature 453 (7199): 1239–1243. doi:10.1038/nature07002.
- ↑ Leupold, U (1950). "Die Vererbung von Homothallie und Heterothallie bei Schizosaccharomyces pombe". CR Trav Lab Carlsberg Ser Physiol 24: 381–480.
- ↑ Leupold U. (1993) The origins of Schizosaccharomyces pombe genetics. In: Hall MN, Linder P. eds. The Early Days of Yeast Genetics . New York. Cold Spring Harbor Laboratory Press. p 125–128.
- ↑ Mitchison, JM (1957). "The growth of single cells. I. Schizosaccharomyces pombe". Exp Cell Res 13: 244–262. doi:10.1016/0014-4827(57)90005-8.
- ↑ Mitchison, JM (1990). "My favourite cell: The fission yeast, Schizosaccharomyces pombe". BioEssays 4: 189–191. doi:10.1002/bies.950120409.
- ↑ Matsuyama; et al. (2006). "ORFeome cloning and global analysis of protein localization in the fission yeast Schizosaccharomyces pombe". Nature Biotechnology 24 (7): 841–847. doi:10.1038/nbt1222.
- ↑ Teoh, Ai Leng (setembro de 2004). "Yeast ecology of Kombucha fermentation". International Journal of Food Microbiology 95 (2): 119–26. PMID 15282124. doi:10.1016/j.ijfoodmicro.2003.12.020.
- ↑ Florenzano; et al. (1977). "Contributo alla ecologia dei lieviti Schizosaccharomyces sulle uve". Vitis 16: 38–44.
- ↑ Gómez; et al. (2002). "Fission yeast enters a joyful new era". Genome Biol 3 (6): 4017. doi:10.1186/gb-2002-3-6-reports4017.
- ↑ Lin; et al. (2011). "The evolution of aerobic fermentation in Schizosaccharomyces pombe was associated with regulatory reprogramming but not nucleosome reorganization". Mol Biol Evol 28 (4): 1407–13. doi:10.1093/molbev/msq324.
- ↑ Grunstein, Michael, and Susan Gasser. "Epigenetics in Saccharomyces cerevisiae." Epigenetics. 1. Cold Spring Harbor Press, 2007.
- ↑ Cell Cycle. Principles of Control" by David O Morgan, Primers in Biology
- ↑ Coelho M, Dereli A, Haese A; et al. (outubro de 2013). "Fission Yeast Does Not Age under Favorable Conditions, but Does So after Stress". Curr. Biol. 23 (19): 1844–52. PMID 24035542. doi:10.1016/j.cub.2013.07.084.
- ↑ A spatial gradient coordinates cell size and mitotic entry in fission yeast by James B. Moseley, Adeline Mayeux, Anne Paoletti & Paul Nurse, Nature, 11 de xuño de 2009
- ↑ Polar gradients of the DYRK-family kinase Pom1 couple cell length with the cell cycle. Sophie G Martin and Martine Berthelot-Grosjean. Nature. 2009
- ↑ Sawin, Kenneth E. (2009). "Cell cycle: Cell division brought down to size". Nature 459 (7248): 782–783. doi:10.1038/459782a.
- ↑ Lessons learned from studies of fission yeast mating-type switching and silencing by Amar J.S. Klar, Annual Review of Genetics, 5 de xullo de 2007
- ↑ Davey J (decembro de 1998). "Fusion of a fission yeast". Yeast 14 (16): 1529–66. PMID 9885154. doi:10.1002/(SICI)1097-0061(199812)14:16<1529::AID-YEA357>3.0.CO;2-0.
- ↑ Bernstein C, Johns V (abril de 1989). "Sexual reproduction as a response to H2O2 damage in Schizosaccharomyces pombe". J. Bacteriol. 171 (4): 1893–7. PMC 209837. PMID 2703462.
- ↑ Bernstein H and Bernstein C (2013). Evolutionary Origin and Adaptive Function of Meiosis. “Meiosis,” Dr. Carol Bernstein (Ed.), ISBN 978-953-511-197-9, InTech, http://www.intechopen.com/books/meiosis/evolutionary-origin-and-adaptive-function-of-meiosis
- ↑ Elvira Hörandl (2013). Meiosis and the Paradox of Sex in Nature, Meiosis, Dr. Carol Bernstein (Ed.), ISBN 978-953-511-197-9, InTech, DOI: 10.5772/56542. Available from: http://www.intechopen.com/books/meiosis/meiosis-and-the-paradox-of-sex-in-nature
- ↑ 22,0 22,1 Pauklin S, Burkert JS, Martin J; et al. (maio de 2009). "Alternative induction of meiotic recombination from single-base lesions of DNA deaminases". Genetics 182 (1): 41–54. PMC 2674839. PMID 19237686. doi:10.1534/genetics.109.101683.
- ↑ Farah JA, Cromie G, Davis L, Steiner WW, Smith GR (decembro de 2005). "Activation of an alternative, rec12 (spo11)-independent pathway of fission yeast meiotic recombination in the absence of a DNA flap endonuclease". Genetics 171 (4): 1499–511. PMC 1456079. PMID 16118186. doi:10.1534/genetics.105.046821.
- ↑ Forsburg, S. L. (Jun 2005). "The yeasts Saccharomyces cerevisiae and Schizosaccharomyces pombe: models for cell biology research". Gravit Space Biol Bull 18 (2): 3–9. PMID 16038088.
- ↑ Forsburg, Susan L; Rhind, Nicholas (febreiro de 2006). "Basic methods for fission yeast". Yeast 23 (3): 173–183. doi:10.1002/yea.1347.
- ↑ 26,0 26,1 26,2 Wixon, Jo (2002). "Featured Organism: Schizosaccharomyces pombe, The Fission Yeast". Comparative and Functional Genomics 3 (2): 194–204. doi:10.1002/cfg.92.
- ↑ 27,0 27,1 27,2 27,3 27,4 27,5 27,6 Wood, V.; Gwilliam, R.; Rajandream, M.-A.; Lyne, M.; Lyne, R.; Stewart, A.; Sgouros, J.; Peat, N.; Hayles, J.; Baker, S.; Basham, D.; Bowman, S.; Brooks, K.; Brown, D.; Brown, S.; Chillingworth, T.; Churcher, C.; Collins, M.; Connor, R.; Cronin, A.; Davis, P.; Feltwell, T.; Fraser, A.; Gentles, S.; Goble, A.; Hamlin, N.; Harris, D.; Hidalgo, J.; Hodgson, G.; Holroyd, S.; Hornsby, T.; Howarth, S.; Huckle, E. J.; Hunt, S.; Jagels, K.; James, K.; Jones, L.; Jones, M.; Leather, S.; McDonald, S.; McLean, J.; Mooney, P.; Moule, S.; Mungall, K.; Murphy, L.; Niblett, D.; Odell, C.; Oliver, K.; O'Neil, S.; Pearson, D.; Quail, M. A.; Rabbinowitsch, E.; Rutherford, K.; Rutter, S.; Saunders, D.; Seeger, K.; Sharp, S.; Skelton, J.; Simmonds, M.; Squares, R.; Squares, S.; Stevens, K.; Taylor, K.; Taylor, R. G.; Tivey, A.; Walsh, S.; Warren, T.; Whitehead, S.; Woodward, J.; Volckaert, G.; Aert, R.; Robben, J.; Grymonprez, B.; Weltjens, I.; Vanstreels, E.; Rieger, M.; Schäfer, M.; Müller-Auer, S.; Gabel, C.; Fuchs, M.; Fritzc, C.; Holzer, E.; Moestl, D.; Hilbert, H.; Borzym, K.; Langer, I.; Beck, A.; Lehrach, H.; Reinhardt, R.; Pohl, T. M.; Eger, P.; Zimmermann, W.; Wedler, H.; Wambutt, R.; Purnelle, B.; Goffeau, A.; Cadieu, E.; Dréano, S.; Gloux, S.; Lelaure, V.; Mottier, S.; Galibert, F.; Aves, S. J.; Xiang, Z.; Hunt, C.; Moore, K.; Hurst, S. M.; Lucas, M.; Rochet, M.; Gaillardin, C.; Tallada, V. A.; Garzon, A.; Thode, G.; Daga, R. R.; Cruzado, L.; Jimenez, J.; Sánchez, M.; del Rey, F.; Benito, J.; Domínguez, A.; Revuelta, J. L.; Moreno, S.; Armstrong, J.; Forsburg, S. L.; Cerrutti, L.; Lowe, T.; McCombie, W. R.; Paulsen, I.; Potashkin, J.; Shpakovski, G. V.; Ussery, D.; Barrell, B. G.; Nurse, P. (21 de febreiro de 2002). "The genome sequence of Schizosaccharomyces pombe". Nature 415 (6874): 871–880. PMID 11859360. doi:10.1038/nature724.
- ↑ 28,0 28,1 28,2 Forsburg, Susan L. "PombeNet". Arquivado dende o orixinal o 06 de xuño de 2015. Consultado o 05 de xuño de 2015.
- ↑ 29,0 29,1 Das, M. (2007). "Regulation of Cell Diameter, For3p Localization, and Cell Symmetry by Fission Yeast Rho-GAP Rga4p". Molecular Biology of the Cell 18 (6): 2090–2101. doi:10.1091/mbc.E06-09-0883.
- ↑ 30,0 30,1 30,2 Kawashima, Shigehiro A.; Takemoto, Ai; Nurse, Paul; Kapoor, Tarun M. (xullo de 2012). "Analyzing Fission Yeast Multidrug Resistance Mechanisms to Develop a Genetically Tractable Model System for Chemical Biology". Chemistry & Biology 19 (7): 893–901. doi:10.1016/j.chembiol.2012.06.008.
- ↑ 31,0 31,1 Tay, Zoey; Eng, Ru Jun; Sajiki, Kenichi; Lim, Kim Kiat; Tang, Ming Yi; Yanagida, Mitsuhiro; Chen, Ee Sin; Mata, Juan (24 de xaneiro de 2013). "Cellular Robustness Conferred by Genetic Crosstalk Underlies Resistance against Chemotherapeutic Drug Doxorubicin in Fission Yeast". PLoS ONE 8 (1): e55041. doi:10.1371/journal.pone.0055041.
- ↑ Davey, John (decembro de 1998). "Fusion of a fission yeast". Yeast 14 (16): 1529–1566. PMID 9885154. doi:10.1002/(SICI)1097-0061(199812)14:16<1529::AID-YEA357>3.0.CO;2-0.
- ↑ Forsburg, Susan L (2006). "Basic methods for fission yeast". Yeast 23 (3): 173–183. doi:10.1002/yea.1347.
- ↑ Forsburg, S. L. (Sep 1999). "The best yeast?". Trends Genet 15 (9): 340–4. PMID 10461200. doi:10.1016/s0168-9525(99)01798-9.
- ↑ Sabatinos, S. A.; Mastro, T. L.; Green, M. D.; Forsburg, S. L. (12 de novembro de 2012). "A Mammalian-Like DNA Damage Response of Fission Yeast to Nucleoside Analogs". Genetics 193 (1): 143–157. doi:10.1534/genetics.112.145730.
- ↑ 36,0 36,1 Hayano, M.; Kanoh, Y.; Matsumoto, S.; Renard-Guillet, C.; Shirahige, K.; Masai, H. (25 de xaneiro de 2012). "Rif1 is a global regulator of timing of replication origin firing in fission yeast". Genes & Development 26 (2): 137–150. doi:10.1101/gad.178491.111.
- ↑ "PomBase Series Edition". The EMBL-European Bioinformatics Institute. Wellcome Trust Genome Campus Place Published: Hinxton, Cambridge.
- ↑ Burrack, Laura S.; Berman, Judith (22 de xuño de 2012). "Neocentromeres and epigenetically inherited features of centromeres". Chromosome Research 20 (5): 607–619. doi:10.1007/s10577-012-9296-x.
- ↑ 39,0 39,1 Stimpson, Kaitlin M.; Matheny, Justyne E.; Sullivan, Beth A. (17 de xullo de 2012). "Dicentric chromosomes: unique models to study centromere function and inactivation". Chromosome Research 20 (5): 595–605. doi:10.1007/s10577-012-9302-3.
- ↑ Kadura, Sheila; Sazer, Shelley (xullo de 2005). "SAC-ing mitotic errors: How the spindle assembly checkpoint (SAC) plays defense against chromosome mis-segregation". Cell Motility and the Cytoskeleton 61 (3): 145–160. doi:10.1002/cm.20072.
- ↑ 41,0 41,1 41,2 41,3 Lee, I-Ju; Coffman, Valerie C.; Wu, Jian-Qiu (outubro de 2012). "Contractile-ring assembly in fission yeast cytokinesis: Recent advances and new perspectives". Cytoskeleton 69 (10): 751–763. doi:10.1002/cm.21052.
- ↑ 42,0 42,1 Teresa, Rinaldi, (2010). "Mitochondrial diseases and the role of the yeast models". FEMS Mini Review.
- ↑ Mojardín, Laura; Vázquez, Enrique; Antequera, Francisco (novembro de 2013). "Specification of DNA Replication Origins and Genomic Base Composition in Fission Yeasts". Journal of Molecular Biology 425 (23): 4706–4713. doi:10.1016/j.jmb.2013.09.023.
- ↑ 44,0 44,1 Forsburg, S. L. (Apr 2002). "Only connect: linking meiotic DNA replication to chromosome dynamics". Mol Cell 9 (4): 703–11.
- ↑ Moriya, Hisao; Chino, Ayako; Kapuy, Orsolya; Csikász-Nagy, Attila; Novák, Béla (6 de decembro de 2011). "Overexpression limits of fission yeast cell-cycle regulators in vivo and in silico". Molecular Systems Biology 7 (1): 556. doi:10.1038/msb.2011.91.
- ↑ Das, Maitreyi; Wiley, David J.; Chen, Xi; Shah, Kavita; Verde, Fulvia (agosto de 2009). "The Conserved NDR Kinase Orb6 Controls Polarized Cell Growth by Spatial Regulation of the Small GTPase Cdc42". Current Biology 19 (15): 1314–1319. doi:10.1016/j.cub.2009.06.057.
- ↑ Moseley, James B. (outubro de 2013). "Cellular Aging: Symmetry Evades Senescence". Current Biology 23 (19): R871–R873. PMID 24112980. doi:10.1016/j.cub.2013.08.013.
- ↑ Cooper, S. (2013). "Schizosaccharomyces pombe grows exponentially during the division cycle with no rate change points.". FEMS Yeast Res.
- ↑ Cadou, Angela (2013). "The Kin1 kinase and the calcineurin phosphatase cooperate to link actin ring assembly and septum synthesis in fission yeast". Biology of the Cell 105 (3): 129–148. doi:10.1111/boc.201200042.
- ↑ 50,0 50,1 Cadou, Angela; Couturier, Anne; Le Goff, Cathy; Xie, Linfeng; Paulson, James R.; Le Goff, Xavier (marzo de 2013). "The Kin1 kinase and the calcineurin phosphatase cooperate to link actin ring assembly and septum synthesis in fission yeast". Biology of the Cell 105 (3): 129–148. doi:10.1111/boc.201200042.
- ↑ Balazs, Anita; Batta, Gyula; Miklos, Ida; Acs-Szabo, Lajos; Vazquez de Aldana, Carlos R.; Sipiczki, Matthias (marzo de 2012). "Conserved regulators of the cell separation process in Schizosaccharomyces". Fungal Genetics and Biology 49 (3): 235–249. doi:10.1016/j.fgb.2012.01.003.
- ↑ Rincon, Sergio A.; Paoletti, Anne (outubro de 2012). "Mid1/anillin and the spatial regulation of cytokinesis in fission yeast". Cytoskeleton 69 (10): 764–777. doi:10.1002/cm.21056.
- ↑ Das, M (2010). "Microtubule-dependent spatial organization of mitochondria in fission yeast Journal". Methods Cell BIo 97: 203–21. PMID 20719273. doi:10.1016/S0091-679X(10)97012-X.
- ↑ Fraser, Hunter B (2013). "Cell-cycle regulated transcription associates with DNA replication timing in yeast and human". Genome Biology 14 (10): R111. doi:10.1186/gb-2013-14-10-r111.
- ↑ Li. C. Green, M. D. Forsburg, S. L. (2013). "Mutations disrupting histone methylation have different effects on replication timing in S. pombe centromere". PLoS ONE 8 (4): e61464. PMID 23658693. doi:10.1371/journal.pone.0061464.
- ↑ "The Official Site of the Nobel Prize".
- ↑ Zhao J, Lendahl U, Nistér M. (2012). "Regulation of mitochondrial dynamics: convergences and divergences between yeast and vertebrates.". J Biomed Biotechnol 70 (6): 951–76. doi:10.1007/s00018-012-1066-6.
- ↑ Abelovska, Lenka (2011). "Mitochondria as protean organelles: membrane processes that influence mitochondrial shape in yeast". General physiology and biophysics 30 (05): 13–24. doi:10.4149/gpb_2011_SI1_13.
- ↑ Chino, Ayako; Makanae, Koji; Moriya, Hisao; Bähler, Jürg (3 de setembro de 2013). "Relationships between Cell Cycle Regulator Gene Copy Numbers and Protein Expression Levels in Schizosaccharomyces pombe". PLoS ONE 8 (9): e73319. doi:10.1371/journal.pone.0073319.
- ↑ 60,0 60,1 Raychaudhuri, Sumana; Young, Barry P; Espenshade, Peter J; Loewen, Christopher JR (agosto de 2012). "Regulation of lipid metabolism: a tale of two yeasts". Current Opinion in Cell Biology 24 (4): 502–508. doi:10.1016/j.ceb.2012.05.006.
- ↑ Babu, Mohan; Vlasblom, James; Pu, Shuye; Guo, Xinghua; Graham, Chris; Bean, Björn D. M.; Burston, Helen E.; Vizeacoumar, Franco J.; Snider, Jamie; Phanse, Sadhna; Fong, Vincent; Tam, Yuen Yi C.; Davey, Michael; Hnatshak, Olha; Bajaj, Navgeet; Chandran, Shamanta; Punna, Thanuja; Christopolous, Constantine; Wong, Victoria; Yu, Analyn; Zhong, Gouqing; Li, Joyce; Stagljar, Igor; Conibear, Elizabeth; Wodak, Shoshana J.; Emili, Andrew; Greenblatt, Jack F. (2 de setembro de 2012). "Interaction landscape of membrane-protein complexes in Saccharomyces cerevisiae". Nature 489 (7417): 585–589. doi:10.1038/nature11354.
- ↑ Suda, Yasuyuki (2011). "The Yeast Golgi Apparatus". Traffic 13 (4): 505–510. PMID 22132734. doi:10.1111/j.1600-0854.2011.01316.x.
- ↑ Forsburg, Susan L. (2013). "PombeNet". Arquivado dende o orixinal o 06 de xuño de 2015. Consultado o Sep 2013.
- ↑ "Trans-NIH.pombe Initiative". 2002.
- ↑ Green, M. D. Sabatinos, S. A. Forsburg, S. L. (2009). "Microscopy techniques to examine DNA replication in fission yeast Journal". Methods Mol Biol 521: 463–82. PMID 19563123. doi:10.1007/978-1-60327-815-7_26.
Véxase tamén
editarOutros artigos
editarLigazóns externas
editar- Víde o
- Pombe Gene Database Arquivado 21 de agosto de 2006 en Wayback Machine.
- Xenoma de S. pombe no Sanger Centre
- European Bioinformatics Institute
- Schizosaccharomyces pombe e MicrobeWiki Arquivado 21 de novembro de 2011 en Wayback Machine.