

Trilobita
| Trilobites | ||
|---|---|---|
| Rango temporal: Cámbrico-Pérmico superior | ||
 Fósil de Eldredgeops rana, un trilobites del Devónico | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Arthropoda | |
| Subfilo: | Trilobitomorpha † | |
| Clase: | Trilobita † | |
| Órdenes | ||
Los trilobites[1] (Trilobita, del latín, "tres lóbulos") son una clase de artrópodos extintos, dentro del subfilo Trilobitomorpha. Son los fósiles más característicos del Paleozoico (antigua «era Primaria»). Se han descrito cerca de 22 000 especies.[2]
Aparecieron en el período Cámbrico (al inicio del Paleozoico, hace unos 540 millones de años), cuando los organismos ediacáricos ya habían desaparecido, y empezaron a diversificarse ya en el Cámbrico inferior. Tras la extinción masiva de finales del Cámbrico solo sobrevivieron las formas que habitaban ambientes pelágicos, de aguas profundas. Durante el Ordovícico alcanzaron su máxima diversidad y ocuparon casi todos los nichos ecológicos marinos. A partir del Silúrico presentaron pocos cambios, hasta que durante la extinción masiva del Devónico sufrieron una importante reducción con la extinción de todos los órdenes excepto Proetida. Durante el Carbonífero los representantes del grupo son escasos y restringidos a ambientes de arrecife. Los últimos trilobites, ya solo habitantes de aguas someras, desaparecieron durante la extinción masiva del Pérmico-Triásico, hace unos 250 millones de años, junto con los euriptéridos o «escorpiones marinos», blastoideos, algunas especies de peces, esponjas, equinodermos, etcétera. Por tanto, su presencia en la Tierra se prolongó durante todo el Paleozoico, casi 300 millones de años. Los restos fósiles de trilobites son tan abundantes y han sido tan profundamente estudiados, que probablemente sean el grupo de animales extintos más conocido.
Inicialmente se consideraron antepasados de los crustáceos (en especial la cochinilla de humedad terrestre, que comparte ciertas características en común) o incluso de todos los artrópodos (ya que fueron los primeros en aparecer en el registro fósil). Hoy se les considera como un grupo independiente, separados de los mandibulados y quelicerados.
Morfología
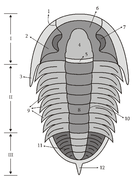
[editar]Los trilobites tienen el cuerpo aplanado y liso, más o menos ovalado y dividido en tres tagmas, céfalon (cabeza), tórax y pigidio; el tórax y el pigidio forman el tronco. Presentan dos surcos longitudinales que dividen el cuerpo en tres lóbulos claramente delimitados (de donde deriva su nombre): uno central (llamado glabela en el céfalon y raquis en el tronco) y dos laterales (denominados genas o mejillas en el céfalon y pleuras en el tronco). El tegumento dorsal era una gruesa cutícula impregnada de carbonato cálcico que ha facilitado su fosilización. Su tamaño varía desde unos pocos milímetros a más de 60 cm en algunas especies gigantes.[3]
-
 Tagmas
Tagmas -
 Morfología
Morfología -
 Céfalon
Céfalon



Céfalon
[editar]
El céfalon ("cabeza") es el resultado de la fusión total de diversos segmentos, y no muestra restos externos de metamerización. Las genas se prolongan por los lados y por detrás por dos puntas genales; las genas están divididas en dos partes por una sutura: una gena fija interna (fixigena) y una gena móvil externa (librigena). La estructura formada por la glabela y las fixigenas se denomina cranidio ("cranidium"). En la cara ventral se localiza el hipostoma, una placa suspendida bajo la glabela que protegía la boca.
Sobre las genas hay un par de grandes ojos compuestos sorprendentemente evolucionados (en algunas especies situados sobre pedúnculos), análogos a los de parientes actuales como los crustáceos e insectos. De hecho, los trilobites fueron los primeros animales en desarrollar ojos complejos, lo que probablemente influyó en su éxito evolutivo. En la parte ventral del céfalon se insertan las antenas unirrámeas, largas y multiarticuladas, y se abre la boca, tras la cual hay tres pares de patas muy similares entre sí. Las antenas son equivalentes, por su posición preoral, a las de los insectos y miriápodos y al primer par de los crustáceos.
Tórax
[editar]
El tórax lo formaban entre 2 y 40 metámeros[3] bien diferenciados; estaban articulados entre sí permitiendo el encabalgamiento de segmentos contiguos, con lo que el animal podía enrollarse en forma de bola.[4] Ventralmente, cada metámero llevaba un par de patas parecidos entre sí y a las del céfalon.
Pigidio
[editar]El pigidio formaba la parte final del cuerpo y constaba de un número variable de metámeros fusionados que formaban una estructura compacta. Cada segmento llevaba un par de patas similares a las del céfalon y del tórax; a veces, había un par de cercos apicales multiarticulados.
Apéndices
[editar]Las patas de los trilobites muestran una sorprendente uniformidad. Usualmente se consideran apéndices birrámeos, que comienzan en una precoxa, de la cual parte un exopodio multiarticulado y con expansiones foliáceas y que probablemente tuviera función branquial y nadadora, y una coxa de la que parte el endopodio, compuesto seis artejos y una uña terminal. Las extremidades estaban protegidas por unas expansiones laterales llamadas lóbulos pleurales, que se extendían hacia afuera desde el lóbulo axial central.
Este tipo de apéndices no son homologables con los apéndices de otros grupos de artrópodos. En los crustáceos, que también tienen apéndices birrámeos, la rama externa (exopodio) se inserta en la coxa. Tampoco tienen relación directa con los apéndices de los Merostomados.
Dimorfismo sexual
[editar]
La excepcional secuencia temporal registrada en los yacimientos cámbricos de Murero y la abundancia de fósiles de trilobites permitió, en el año 2003,[7] identificar dimorfismo sexual en trilobites, en aquella fecha el caso más antiguo reconocido en el reino animal. Para ello se seleccionaron cuatro especies filogenéticamente próximas y de amplia distribución estratigráfica: Acadoparadoxides mureroensis, Eccaparadoxides rouvielli, E. sequeirosi y E. mediterraneus. En todos los casos se comprobó que se presentaban dos morfotipos diferentes para cada especie y que esta diferencia se mantenía a lo largo de la sucesión sedimentaria o, lo que es equivalente, en el tiempo.
Los rasgos que diferencian ambos morfotipos son diferentes para cada especie, pero incluyen tanto la longitud del pigidio, más corto en los machos, como variaciones en la longitud de algunos segmentos del tórax, presentando las hembras una zona más estrecha en el tórax, correspondiente a segmentos posteriores al segmento en el que probablemente se ubicaría el poro genital (el segundo segmento torácico, como en los xifosuros actuales).[8]
Paleobiología
[editar]El desarrollo de los trilobites comportaba una serie de estadios larvales. Las especies más primitivas presentaban un desarrollo larvario completo mientras que en las posteriores el proceso se simplificaba.
El primer estadio larval se conoce como protaspis, formado básicamente por el céfalon y el protopigidio. A este seguía el estadio meraspis en el que se diferenciaban ya algunos segmentos del tórax y el pigidio. El tercer período u holaspis comprende las larvas que ya han adquirido la metamerización completa, pero son aún mucho más pequeñas que un adulto, al que se llegaba después de una serie de mudas.
Paleoecología
[editar]Exclusivamente marinos, estaban totalmente ausentes de ambientes de agua dulce y salobre; por su forma aplanada, ojos en posición dorsal y dureza de la cara dorsal se deduce que la mayoría eran animales bentónicos. Seguramente eran micrófagos, filtrando el barro del fondo en que vivían para obtener el alimento, ya que carecían de apéndices excavadores o prénsiles, así como de piezas bucales trituradoras. Algunas especies se hicieron secundariamente pelágicas y desarrollaron expansiones espinosas para favorecer la flotabilidad. Estas espinas también estaban presentes en las larvas protaspis, que son, por tanto, consideradas formas pelágicas con un gran potencial colonizador.
La reducción y pérdida de los ojos experimentada por diversas especies está relacionada seguramente con una adaptación a la zona afótica y la colonización de aguas profundas.
Tafonomía
[editar]Como consecuencia de las mudas es muy frecuente el hallazgo de exuvias desarticuladas en el registro fósil, sobre todo cefalones —o cranidios y mejillas librígenas por separado— y pigidios (un mismo individuo puede producir indicios múltiples de su existencia).
Icnofósiles
[editar]

A la actividad biológica de los trilobites se atribuyen, principalmente, los icnofósiles Cruziana, Diplichnites y Rusophycus que, por otra parte, son también realizados por otros organismos.[9] Normalmente se encuentran como contramoldes, en relieve invertido, en la base (cara inferior) de capas de arenisca o cuarcita.
Cruziana se interpreta como pistas fósiles debidas a la locomoción y la alimentación (repichnia y pascichnia) de trilobites en el fondo arenoso, e incluye, dada la gran diversidad de Trilobita, numerosas paraespecies. Son pistas longitudinales que en epirelieve aparecen formadas por dos surcos que dejan una cresta central, en ocasiones con otros dos surcos más pequeños laterales, y en hiporelieves como dos crestas longitudinales con un surco central. En ambos casos aparecen finas estriaciones oblicuas en forma de "V" —que indican el sentido contrario al del avance— formadas por los apéndices. También han recibido el nombre informal de "Bilobites", actualmente en desuso.[10]
Diplichnites se interpreta como pistas del desplazamiento rápido (repichnia) sobre la superficie arenosa. Aparece como dos pistas paralelas formadas por multitud de pequeñas huellas estrechamente espaciadas entre sí. Las pequeñas huellas se corresponden con los apéndices del organismo y su profundidad y distribución indican las condiciones de este desplazamiento. En ocasiones las huellas aparecen como finas estriaciones cuando el organismo sufrió un desplazamiento lateral debido a corrientes marinas y a veces evoluciona hacia rastros tipo Cruziana a través de formas de transición.[11]
Rusophycus se interpreta como huellas de reposo (cubichnia) o quizás de alimentación. Se presentan como dos surcos cortos y finamente estriados, más profundos que Cruziana y por lo general aislados de otras pistas. Son resultado de un proceso de excavación o enterramiento del organismo productor para su ocultamiento o acecho en el que se marcan dos lóbulos estriados formados por el movimiento de los dos grupos de apéndices.[12]
Primeros trilobites
[editar]Los trilobites más antiguos que se conocen del registro fósil son falotaspidoideos y redlichioideos, ambos del orden Redlichiida, y bigotínidos (orden Ptychoparida, superfamilia Ellipsocephaloidea), datados de 540 a 520 millones de años.[13][14] Los primeros trilobites incluyen a Profallotaspis jakutensis (Siberia), Fritzaspis spp. (oeste de Estados Unidos), Hupetina antiqua (Marruecos)[15] y Serrania gordaensi (España).[16] Se piensa que los trilobites se originaron en lo que hoy es Siberia, con posterior distribución y radiación desde esta región.[13]
-

-

-
 Diminuto trilobites (Elrathia kingii) de 1 cm de longitud.
Diminuto trilobites (Elrathia kingii) de 1 cm de longitud. -

-






Referencias
[editar]Notas
[editar]Referencias
[editar]- ↑ Trilobites, forma idéntica tanto para el singular como para el plural (plurale tantum).
- ↑ Paterson, John R. (2020-01). «The trouble with trilobites: classification, phylogeny and the cryptogenesis problem». Geological Magazine (en inglés) 157 (1): 35-46. ISSN 0016-7568. doi:10.1017/S0016756819000426. Consultado el 14 de octubre de 2022.
- ↑ a b Riba, O. & Reguant, S., 1988. Registre fòssil. Història Natural dels Països Catalans, 8. Enciclopèdia Catalana, S. A., Barcelona, 478 pp. ISBN 84-7739-022-3.
- ↑ Scholtz, Gerhard (2008). «Scarab beetles at the interface of wheel invention in nature and culture?». Contributions to Zoology (National Museum of Natural History Naturalis) 77 (3). ISSN 1875-9866. Archivado desde el original el 21 de octubre de 2016. Consultado el 19 de enero de 2009.
- ↑ Dies Álvarez, M. E.; Rushtonc, A. W. A.; Gozalo, R.; Pillola, G. L.; Liñán, E. y Ahlber, P. (2010) «Paradoxides brachyrhachis Linnarsson, 1883 versus Paradoxides mediterraneus Pompeckj, 1901: a problematic determination». GFF, 132: 95-104
- ↑ Gozalo, R.; Andrés, J. A.; Chirivella, J. B.; Dies Álvarez, M. E.; Esteve, J.; Gámez Vintaned, J. A.; Mayoral, E.; Zamora, S. y Liñán, E. (2010) «Murero y la explosión del Cámbrico: controversias acerca de este acontecimiento Archivado el 13 de noviembre de 2013 en Wayback Machine.». Enseñanza de las Ciencias de la Tierra, 18(1): 47-59
- ↑ Gozalo, R.; Liñán, E. y Dies, M. E. (2003) «Intraspecific dimorphism in an evolutionary series of paradoxidids from the Middle Cambrian of Murero, Spain». En: Lane, P. D.; Siveter, D. J. y Fortey, R. A. (eds.) Trilobites and their relatives (proceedings of Oxford conference 2001). Special Papers in Palaeontology, 70: 141-156
- ↑ Liñán, E.; Gámez Vintaned, J. A.; Gozalo, R.; Dies Álvarez, M. E.; Chirivella Martorell, J. B. y Esteve, J. (2009) «Las especies de "paradoxides" del yacimiento cámbrico de Murero y el hallazgo de dimorfismo sexual en los trilobites Archivado el 10 de junio de 2015 en Wayback Machine.». Naturaleza Aragonesa, 23: 4-11
- ↑ Luis A. Buatois, M. Gabriela Mángano (2011). Ichnology: Organism-Substrate Interactions in Space and Time. Cambridge University Press.
- ↑ KU Icnology (ed.). «Cruziana». Consultado el 18 de agosto de 2015.
- ↑ KU Icnology (ed.). «Diplichnites». Consultado el 18 de agosto de 2015.
- ↑ KU Icnology (ed.). «Rusophycus». Consultado el 18 de agosto de 2015.
- ↑ a b Lieberman, B. S. (2002) «Phylogenetic analysis of some basal early Cambrian trilobites, the biogeographic origins of the eutrilobita, and the timing of the Cambrian radiation Archivado el 5 de marzo de 2016 en Wayback Machine.». Journal of Paleontology, 76(4): 692–708
- ↑ Fortey, Richard (2000) Trilobite!: Eyewitness to Evolution. Londres: Harper-Collins ISBN 0-00-257012-2
- ↑ Hollingsworth, J. S. (2008). The first trilobites in Laurentia and elsewhere. En Rábano, I.; Gozalo, R. y García-Bellido, D., ed. «Advances in trilobite research». Cuadernos del Museo Geominero 9 (Madrid: Instituto Geológico y Minero de España).
- ↑ Liñán, Eladio; Gozalo, Rodolfo; Dies Alvarez, María Eugenia (2008). «Nuevos trilobites del Ovetiense inferior (Cámbrico Inferior bajo) de Sierra Morena (España)». Ameghiniana 45 (1): 123-138.
Bibliografía
[editar]- Liñán, E. (1996): Los trilobites. En: Melic, A. (Ed.): PaleoEntomología [sic]. Boletín de la Sociedad Entomológica Aragonesa, 16: 45-56
- Rábano, I. (1999): Historia evolutiva de los trilobites. En: Melic, A.; Haro, J.J. de; Méndez, M. y Ribera, I. (Eds.): Evolución y Filogenia de Arthropoda. Boletín de la Sociedad Entomológica Aragonesa, 26: 225-233
Enlaces externos
[editar] Wikimedia Commons alberga una categoría multimedia sobre Trilobita.
Wikimedia Commons alberga una categoría multimedia sobre Trilobita.- A Guide to the Orders of Trilobites, sitio web sobre trilobites.
- El yacimiento con los mayores trilobites del mundo, una investigación española y portuguesa.