
The Central Conserved Peptides of Respiratory Syncytial Virus G Protein Enhance the Immune Response to the RSV F Protein in an Adenovirus Vector Vaccine Candidate

 ,
, <p>Designing and characterizing the Ad-based vaccines. (<b>a</b>) Schematic of Ad5-PreF, Ad5-Gcc, Ad5-PreF-P2A-Gcc and Ad5-PreF-Qa-P2A-Gcc. (<b>b</b>,<b>c</b>) Western blot analysis of F expression in cell lysis (<b>b</b>) and supernatant (<b>c</b>). (<b>d</b>,<b>e</b>) Western blot analysis of Gcc expression in cell lysis (<b>d</b>) and supernatant (<b>e</b>). (<b>f</b>) Identification of the conformation of F protein. Ad5-empty was used as control. Site Ø represents the unique antigenic sites of PreF.</p> "> Figure 2
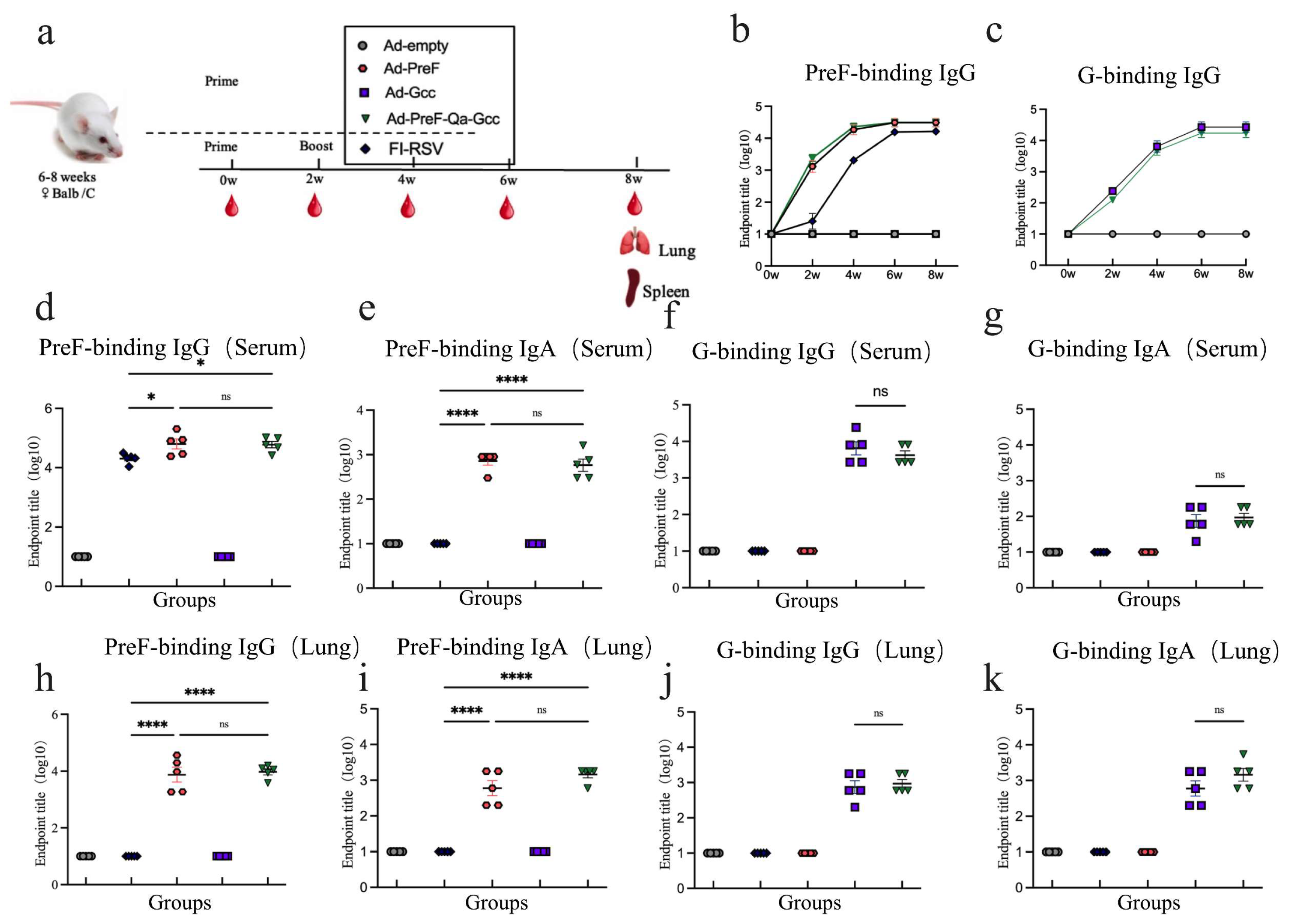
<p>Immune responses in immunized Balc/c mice. (<b>a</b>) Schedule of Ad5 vaccines immunization and bleeding strategies in female Balb/C mice. (<b>b</b>) Kinetics of PreF-specific total IgG reciprocal endpoint titers (log10) were measured within 8 weeks of initial vaccination. (<b>c</b>) Kinetics of G-specific total IgG reciprocal endpoint titers (log10) were measured within 8 weeks after vaccination. (<b>d</b>) Endpoint titer of PreF-specific total IgG was measured in serum at week 6. (<b>e</b>) Endpoint titer of PreF-specific total IgA was measured in serum at week 6. (<b>f</b>) Endpoint titer of G-specific total IgG was measured in serum at week 6. (<b>g</b>) Endpoint titer of G-specific total IgA was measured in serum at week 6. (<b>h</b>) Endpoint titer of PreF-specific total IgG was measured in BALF at week 8. (<b>i</b>) Endpoint titer of PreF-specific total IgA was measured in BALF at week 8. (<b>j</b>) Endpoint titer of G-specific total IgG was measured in BALF at week 8. (<b>k</b>) Endpoint titer of G-specific total IgA was measured in BALF at week 8. ns = not significant; * <span class="html-italic">p</span> < 0.05, **** <span class="html-italic">p</span> < 0.0001.</p> "> Figure 3
<p>Intranasal immunization induced a balanced Th1/Th2 immune response in mice vaccinated with Ad-PreF-Qa-P2A-Gcc. (<b>a</b>–<b>c</b>) Anti-PreF antibody subtype IgG1 (<b>a</b>) and IgG2a (<b>b</b>) were detected in serum at week 6, and the ratio of IgG2a/IgG1 (<b>c</b>) was calculated. (<b>d</b>–<b>f</b>) Anti-G antibody subtypes IgG1 (<b>d</b>) and IgG2a (<b>e</b>) were detected in serum at week 6, and the ratio of IgG2a/IgG1 (<b>f</b>) was calculated. ns = not significant; *** <span class="html-italic">p</span> < 0.001.</p> "> Figure 4
<p>Mice vaccinated with Ad5-PreF-Qa-Gcc induced superior neutralizing antibody. (<b>a</b>,<b>b</b>) Serum-neutralizing antibody response against RSV Long (<b>a</b>) and RSV B1 (<b>b</b>) at week 6. All data are shown as means ± SEM. <span class="html-italic">p</span>-values were analyzed with one-way ANOVA (* <span class="html-italic">p</span> < 0.05; ** <span class="html-italic">p</span> < 0.01; **** <span class="html-italic">p</span> < 0.0001).</p> "> Figure 5
<p>Immunogenicity and protective efficacy in BALB/c mice. (<b>a</b>) Schematics of vaccine immunization and challenge in BALB/c mice (6–8 weeks old). Mice were challenged with 2 × 10<sup>6</sup> pfu RSV Long via the i.n. route at 4 weeks after boost vaccination. (<b>b</b>) Body weight change over 5 days post-infection. (<b>c</b>,<b>d</b>) Copies of RSV viral L gene in lung tissue (<b>c</b>) and nasal tissue (<b>d</b>). The limit of detection for the RT-qPCRs is 10 copies/mL. (<b>e</b>–<b>j</b>) Tissue sections of the lung tissues were stained with H&E and PAS for pathological examination. The magnification of all images is 20x. All data are shown as means ± SEM. <span class="html-italic">p</span>-values were analyzed with one-way ANOVA (* <span class="html-italic">p</span> < 0.05; ** <span class="html-italic">p</span> < 0.01; *** <span class="html-italic">p</span> < 0.001; **** <span class="html-italic">p</span> < 0.0001).</p> "> Figure 6
<p>Treg cells and Th17 cells in the lung of vaccinated mice induced by RSV challenge infection. Vaccinated mice were challenged i.n. with RSV Long at 6 weeks after the immunization. (<b>a</b>) The percentage of CD4<sup>+</sup>CD25<sup>+</sup>Foxp3<sup>+</sup> Treg cells. (<b>b</b>) The percentage of IL-17A<sup>+</sup> CD4<sup>+</sup> T cells. All data are shown as means ± SEM. <span class="html-italic">p</span>-values were analyzed with one-way ANOVA (** <span class="html-italic">p</span> < 0.01; *** <span class="html-italic">p</span> < 0.001; **** <span class="html-italic">p</span> < 0.0001).</p> ">
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Vaccine Preparation
2.3. Western Blot
2.4. Characterization of RSV PreF
2.5. Immunization and Challenge of Mice
2.6. Antibody ELISAs
2.7. Neutralization Assay
2.8. Histological Analysis
2.9. Treg Cells and Th17 Cells
2.10. Quantitative RT-PCR Assay
2.11. Statistical Analysis
3. Results
3.1. A Heptapeptide, Qa, Can Boost Protein Expression and Improve Self-Cleaving Efficiency
3.2. Intranasal Immunization with Ad5-PreF-Qa-Gcc Induces a High-Level Antibody Response in Mice
3.3. The Addition of the Gcc Antigen Improved the Neutralizing Antibody Titers against RSV Long and RSV B1
3.4. Intranasal Immunization Provided Protection against RSV Long In Vivo
3.5. PreF-Gcc Immunization Enhanced Treg Cells in the Post-Challenge Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rijsbergen, L.C.; Lamers, M.M.; Comvalius, A.D.; Koutstaal, R.W.; Schipper, D.; Duprex, W.P.; Haagmans, B.L.; de Vries, R.D.; de Swart, R.L. Human Respiratory Syncytial Virus Subgroup A and B Infections in Nasal, Bronchial, Small-Airway, and Organoid-Derived Respiratory Cultures. mSphere 2021, 6, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Freitas, F.T.M.; Pimentel, C.C.P.; Bianchini, P.R.; Carvalho, R.M.; Serafim, A.P.; Costa, C.F.A. Evaluation of Severe Acute Respiratory Syndrome surveillance caused by respiratory viruses in a pediatric unit, 2013 to 2019. Rev. Paul. Pediatr. 2023, 42, e2022215. [Google Scholar] [CrossRef] [PubMed]
- Tabor, D.E.; Fernandes, F.; Langedijk, A.C.; Wilkins, D.; Lebbink, R.J.; Tovchigrechko, A.; Ruzin, A.; Kragten-Tabatabaie, L.; Jin, H.; Esser, M.T.; et al. Global Molecular Epidemiology of Respiratory Syncytial Virus from the 2017–2018 INFORM-RSV Study. J. Clin. Microbiol. 2020, 59, 10-1128. [Google Scholar] [CrossRef]
- Fuentes, S.; Coyle, E.M.; Beeler, J.; Golding, H.; Khurana, S. Antigenic Fingerprinting following Primary RSV Infection in Young Children Identifies Novel Antigenic Sites and Reveals Unlinked Evolution of Human Antibody Repertoires to Fusion and Attachment Glycoproteins. PLoS Pathog. 2016, 12, e1005554. [Google Scholar] [CrossRef]
- Fuentes, S.; Coyle, E.M.; Golding, H.; Khurana, S. Nonglycosylated G-Protein Vaccine Protects against Homologous and Heterologous Respiratory Syncytial Virus (RSV) Challenge, while Glycosylated G Enhances RSV Lung Pathology and Cytokine Levels. J. Virol. 2015, 89, 8193–8205. [Google Scholar] [CrossRef]
- Battles, M.B.; McLellan, J.S. Respiratory syncytial virus entry and how to block it. Nat. Rev. Microbiol. 2019, 17, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, D.A.; Baradaran, K.; McIntosh, K.; Patterson, J.L. Appearance of a soluble form of the G protein of respiratory syncytial virus in fluids of infected cells. J. Gen. Virol. 1987, 68 Pt 6, 1705–1714. [Google Scholar] [CrossRef]
- Chirkova, T.; Lin, S.; Oomens, A.G.P.; Gaston, K.A.; Boyoglu-Barnum, S.; Meng, J.; Stobart, C.C.; Cotton, C.U.; Hartert, T.V.; Moore, M.L.; et al. CX3CR1 is an important surface molecule for respiratory syncytial virus infection in human airway epithelial cells. J. Gen. Virol. 2015, 96, 2543–2556. [Google Scholar] [CrossRef]
- Johnson, S.M.; McNally, B.A.; Ioannidis, I.; Flano, E.; Teng, M.N.; Oomens, A.G.; Walsh, E.E.; Peeples, M.E. Respiratory Syncytial Virus Uses CX3CR1 as a Receptor on Primary Human Airway Epithelial Cultures. PLoS Pathog. 2015, 11, e1005318. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Saribas, A.S.; Liu, J.; Lin, Y.; Bodnar, B.; Zhao, R.; Guo, Q.; Ting, J.; Wei, Z.; Ellis, A.; et al. Protein expression/secretion boost by a novel unique 21-mer cis-regulatory motif (Exin21) via mRNA stabilization. Mol. Ther. 2023, 31, 1136–1158. [Google Scholar] [CrossRef]
- Brakel, K.A.; Ma, Y.; Binjawadagi, R.; Harder, O.; Watts, M.; Li, J.; Binjawadagi, B.; Niewiesk, S. Codon-optimization of the respiratory syncytial virus (RSV) G protein expressed in a vesicular stomatitis virus (VSV) vector improves immune responses in a cotton rat model. Virology 2022, 575, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Krarup, A.; Truan, D.; Furmanova-Hollenstein, P.; Bogaert, L.; Bouchier, P.; Bisschop, I.J.M.; Widjojoatmodjo, M.N.; Zahn, R.; Schuitemaker, H.; McLellan, J.S.; et al. A highly stable prefusion RSV F vaccine derived from structural analysis of the fusion mechanism. Nat. Commun. 2015, 6, 8143. [Google Scholar] [CrossRef] [PubMed]
- Rainho-Tomko, J.N.; Pavot, V.; Kishko, M.; Swanson, K.; Edwards, D.; Yoon, H.; Lanza, L.; Alamares-Sapuay, J.; Osei-Bonsu, R.; Mundle, S.T.; et al. Immunogenicity and protective efficacy of RSV G central conserved domain vaccine with a prefusion nanoparticle. NPJ Vaccines 2022, 7, 74. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, K.; Xing, M.; Zhuo, Y.; Guo, J.; Du, M.; Wang, Q.; An, Y.; Li, J.; Gao, P.; et al. Heterologous prime-boost immunizations with chimpanzee adenoviral vectors elicit potent and protective immunity against SARS-CoV-2 infection. Cell Discov. 2021, 7, 123. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.R.; Walsh, E.E. Formalin-inactivated respiratory syncytial virus vaccine induces antibodies to the fusion glycoprotein that are deficient in fusion-inhibiting activity. J. Clin. Microbiol. 1988, 26, 1595–1597. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.M.; Lee, Y.; Kim, K.H.; Jung, Y.J.; Li, Z.; Jeeva, S.; Lee, S.; Moore, M.L.; Kang, S.M. Antigenicity and immunogenicity of unique prefusion-mimic F proteins presented on enveloped virus-like particles. Vaccine 2019, 37, 6656–6664. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Qin, H.; Lei, L.; Lou, W.; Li, R.; Pan, Z. Virus-like particles containing a prefusion-stabilized F protein induce a balanced immune response and confer protection against respiratory syncytial virus infection in mice. Front. Immunol. 2022, 13, 1054005. [Google Scholar] [CrossRef] [PubMed]
- Greinacher, A.; Thiele, T.; Warkentin, T.E.; Weisser, K.; Kyrle, P.A.; Eichinger, S. Thrombotic Thrombocytopenia after ChAdOx1 nCov-19 Vaccination. N. Engl. J. Med. 2021, 384, 2092–2101. [Google Scholar] [CrossRef]
- Hwang, J.; Lee, S.B.; Lee, S.W.; Lee, M.H.; Koyanagi, A.; Jacob, L.; Tizaoui, K.; Yon, D.K.; Shin, J.I.; Smith, L. Comparison of vaccine-induced thrombotic events between ChAdOx1 nCoV-19 and Ad26.COV.2.S vaccines. J. Autoimmun. 2021, 122, 102681. [Google Scholar] [CrossRef]
- Afkhami, S.; D’Agostino, M.R.; Zhang, A.; Stacey, H.D.; Marzok, A.; Kang, A.; Singh, R.; Bavananthasivam, J.; Ye, G.; Luo, X.; et al. Respiratory mucosal delivery of next-generation COVID-19 vaccine provides robust protection against both ancestral and variant strains of SARS-CoV-2. Cell 2022, 185, 896–915.e19. [Google Scholar] [CrossRef]
- Mekseepralard, C.; Toms, G.L.; Routledge, E.G. Protection of mice against Human respiratory syncytial virus by wild-type and aglycosyl mouse-human chimaeric IgG antibodies to subgroup-conserved epitopes on the G glycoprotein. J. Gen. Virol. 2006, 87, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.E.; Pérez Marc, G.; Zareba, A.M.; Falsey, A.R.; Jiang, Q.; Patton, M.; Polack, F.P.; Llapur, C.; Doreski, P.A.; Ilangovan, K.; et al. Efficacy and Safety of a Bivalent RSV Prefusion F Vaccine in Older Adults. N. Engl. J. Med. 2023, 388, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Falsey, A.R.; Williams, K.; Gymnopoulou, E.; Bart, S.; Ervin, J.; Bastian, A.R.; Menten, J.; De Paepe, E.; Vandenberghe, S.; Chan, E.K.H.; et al. Efficacy and Safety of an Ad26.RSV.preF-RSV preF Protein Vaccine in Older Adults. N. Engl. J. Med. 2023, 388, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.S.; Kwon, Y.M.; Lee, J.S.; Yoo, S.E.; Lee, Y.N.; Ko, E.J.; Kim, M.C.; Cho, M.K.; Lee, Y.T.; Jung, Y.J.; et al. Co-immunization with virus-like particle and DNA vaccines induces protection against respiratory syncytial virus infection and bronchiolitis. Antiviral Res. 2014, 110, 115–123. [Google Scholar] [CrossRef] [PubMed]
- van der Fits, L.; Bolder, R.; Heemskerk-van der Meer, M.; Drijver, J.; van Polanen, Y.; Serroyen, J.; Langedijk, J.P.M.; Schuitemaker, H.; Saeland, E.; Zahn, R. Adenovector 26 encoded prefusion conformation stabilized RSV-F protein induces long-lasting Th1-biased immunity in neonatal mice. NPJ Vaccines 2020, 5, 49. [Google Scholar] [CrossRef] [PubMed]
- Joyce, C.; Scallan, C.D.; Mateo, R.; Belshe, R.B.; Tucker, S.N.; Moore, A.C. Orally administered adenoviral-based vaccine induces respiratory mucosal memory and protection against RSV infection in cotton rats. Vaccine 2018, 36, 4265–4277. [Google Scholar] [CrossRef] [PubMed]
- Ruckwardt, T.J.; Bonaparte, K.L.; Nason, M.C.; Graham, B.S. Regulatory T cells promote early influx of CD8+ T cells in the lungs of respiratory syncytial virus-infected mice and diminish immunodominance disparities. J. Virol. 2009, 83, 3019–3028. [Google Scholar] [CrossRef] [PubMed]
- Fulton, R.B.; Meyerholz, D.K.; Varga, S.M. Foxp3+ CD4 regulatory T cells limit pulmonary immunopathology by modulating the CD8 T cell response during respiratory syncytial virus infection. J. Immunol. 2010, 185, 2382–2392. [Google Scholar] [CrossRef]
- Loebbermann, J.; Durant, L.; Thornton, H.; Johansson, C.; Openshaw, P.J. Defective immunoregulation in RSV vaccine-augmented viral lung disease restored by selective chemoattraction of regulatory T cells. Proc. Natl. Acad. Sci. USA 2013, 110, 2987–2992. [Google Scholar] [CrossRef]
- Durant, L.R.; Makris, S.; Voorburg, C.M.; Loebbermann, J.; Johansson, C.; Openshaw, P.J. Regulatory T cells prevent Th2 immune responses and pulmonary eosinophilia during respiratory syncytial virus infection in mice. J. Virol. 2013, 87, 10946–10954. [Google Scholar] [CrossRef]
- Boukhvalova, M.S.; Prince, G.A.; Blanco, J.C. The cotton rat model of respiratory viral infections. Biologicals 2009, 37, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Cox, F.; Saeland, E.; Thoma, A.; van den Hoogen, W.; Tettero, L.; Drijver, J.; Vaneman, C.; van Polanen, Y.; Ritschel, T.; Bastian, A.R.; et al. RSV A2-Based Prefusion F Vaccine Candidates Induce RSV A and RSV B Cross Binding and Neutralizing Antibodies and Provide Protection against RSV A and RSV B Challenge in Preclinical Models. Vaccines 2023, 11, 672. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Nucleotides Sequence (5′-3′) |
|---|---|
| RSV Long q-PCR-F | GAACTCAGTGTAGGTAGAATGTTTGCA |
| RSV Long q-PCR-R | TTCAGCTATCATTTTCTCTGCCAAT |
| RSV Long probe | 5′FAMATTTGAACCTGTCTGAACATTCCCGGTTGCAT 3′BHQ1 |
| Inoculum | Alveolitis a | Interstitial Pneumonia a | Perivascular Bronchitis a |
|---|---|---|---|
| PBS | 0 | 0 | 0 |
| Ad5-empty | 1.6 ± 0.08 | 1.6 ± 0.15 | 2.25 ± 0.09 |
| FI-RSV | 2.12 ± 0.13 | 2.21 ± 0.12 | 2.9 ± 0.1 |
| Ad5-PreF | 0 | 0.6 ± 0.08 | 1.02 ± 0.05 |
| Ad5-Gcc | 0.82 ± 0.08 | 1.51 ± 0.05 | 1.51 ± 0.08 |
| Ad5-PreF-Qa-Gcc | 0 | 0.8 ± 0.12 | 1.15 ± 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chai, P.; Shi, Y.; Yu, J.; Liu, X.; Li, D.; Li, J.; Li, L.; Li, D.; Duan, Z. The Central Conserved Peptides of Respiratory Syncytial Virus G Protein Enhance the Immune Response to the RSV F Protein in an Adenovirus Vector Vaccine Candidate. Vaccines 2024, 12, 807. https://doi.org/10.3390/vaccines12070807
Chai P, Shi Y, Yu J, Liu X, Li D, Li J, Li L, Li D, Duan Z. The Central Conserved Peptides of Respiratory Syncytial Virus G Protein Enhance the Immune Response to the RSV F Protein in an Adenovirus Vector Vaccine Candidate. Vaccines. 2024; 12(7):807. https://doi.org/10.3390/vaccines12070807
Chicago/Turabian StyleChai, Pengdi, Yi Shi, Junjie Yu, Xiafei Liu, Dongwei Li, Jinsong Li, Lili Li, Dandi Li, and Zhaojun Duan. 2024. "The Central Conserved Peptides of Respiratory Syncytial Virus G Protein Enhance the Immune Response to the RSV F Protein in an Adenovirus Vector Vaccine Candidate" Vaccines 12, no. 7: 807. https://doi.org/10.3390/vaccines12070807