
First Report on Choanephora cucurbitarum Causing Choanephora Rot in Chenopodium Plants and Its Sensitivity to Fungicide


<p>Symptoms of Choanephora rot on panicle necks, stems, and leaves of <span class="html-italic">Chenopodium quinoa</span> (<b>A</b>) panicle necks and stems of <span class="html-italic">C. formosanum</span> (<b>B</b>) and stems of <span class="html-italic">C. album</span> (<b>C</b>).</p> "> Figure 2
<p>Morphological characteristics of <span class="html-italic">Choanephora cucurbitarum</span> from <span class="html-italic">Chenopodium</span> plants. (<b>A</b>) Colony on PDA for 2 days, (<b>B</b>) sporangiophores bearing sporangiola, (<b>C</b>–<b>G</b>) sporangiophores with apically dilated and bearing secondary vesicles, (<b>H</b>,<b>I</b>) sporangiola readily detached at maturity, (<b>J</b>–<b>L</b>) secondary vesicles with a head of sporangiola, (<b>M</b>) mature sporangiola with mulberry-like head, (<b>N</b>) sporangiola with longitudinal striation, (<b>O</b>) sporangiophores bearing sporangia, (<b>P</b>,<b>Q</b>) pale yellow to yellow sporangium, (<b>R</b>) sporangiophore bearing mature sporangium, (<b>S</b>,<b>T</b>) dehiscent sporangium, (<b>U</b>) sporangiospores from sporangia, and (<b>V</b>) sporangiospores with appendages.</p> "> Figure 3
<p>Phylogenetic tree of <span class="html-italic">Choanephora cucurbitarum</span> isolates (LMJM-2, LMJM-3, LMJM-5, LMJM-7, and LMJM-9) from <span class="html-italic">Chenopodium</span> plants and their related isolates based on LSU and ITS sequences using the maximum likelihood (ML) method. Ex-type strains were indicated with (T) in the end of the taxa labels, and our strains are in bold.</p> "> Figure 4
<p>Germination of the sporangiola (<b>A</b>) and sporangiospores (<b>B</b>) of <span class="html-italic">Choanephora cucurbitarum</span> at 2, 3, and 4 h (<b>C</b>).</p> "> Figure 5
<p>Symptoms on <span class="html-italic">Chenopodium quinoa</span> (<b>A</b>), <span class="html-italic">C. formosanum</span> (<b>B</b>), and <span class="html-italic">C. album</span> (<b>C</b>) induced by inoculation of representative isolates (LMJM-2, LMJM-3, LMJM-5, LMJM-7, and LMJM-9) of <span class="html-italic">Choanephora cucurbitarum</span>.</p> "> Figure 6
<p>Effect of temperature on pathogenicity of the representative isolates of <span class="html-italic">Choanephora cucurbitarum</span>. (<b>A</b>) Pathogenicity on stems of <span class="html-italic">C. quinoa</span>, <span class="html-italic">C. formosanum</span>, and <span class="html-italic">C. album</span> at different temperatures, (<b>B</b>) lesions lengths on panicle necks of white and red quinoa that were inoculated with <span class="html-italic">Choanephora cucurbitarum</span> and incubated at different temperatures. Data were analyzed with SPSS statistics 19.0 by one-way ANOVA, and means were compared using Tukey’s test at a significance level of <span class="html-italic">p</span> = 0.05. Different letters indicate significant differences (<span class="html-italic">p</span> = 0.05).</p> "> Figure 7
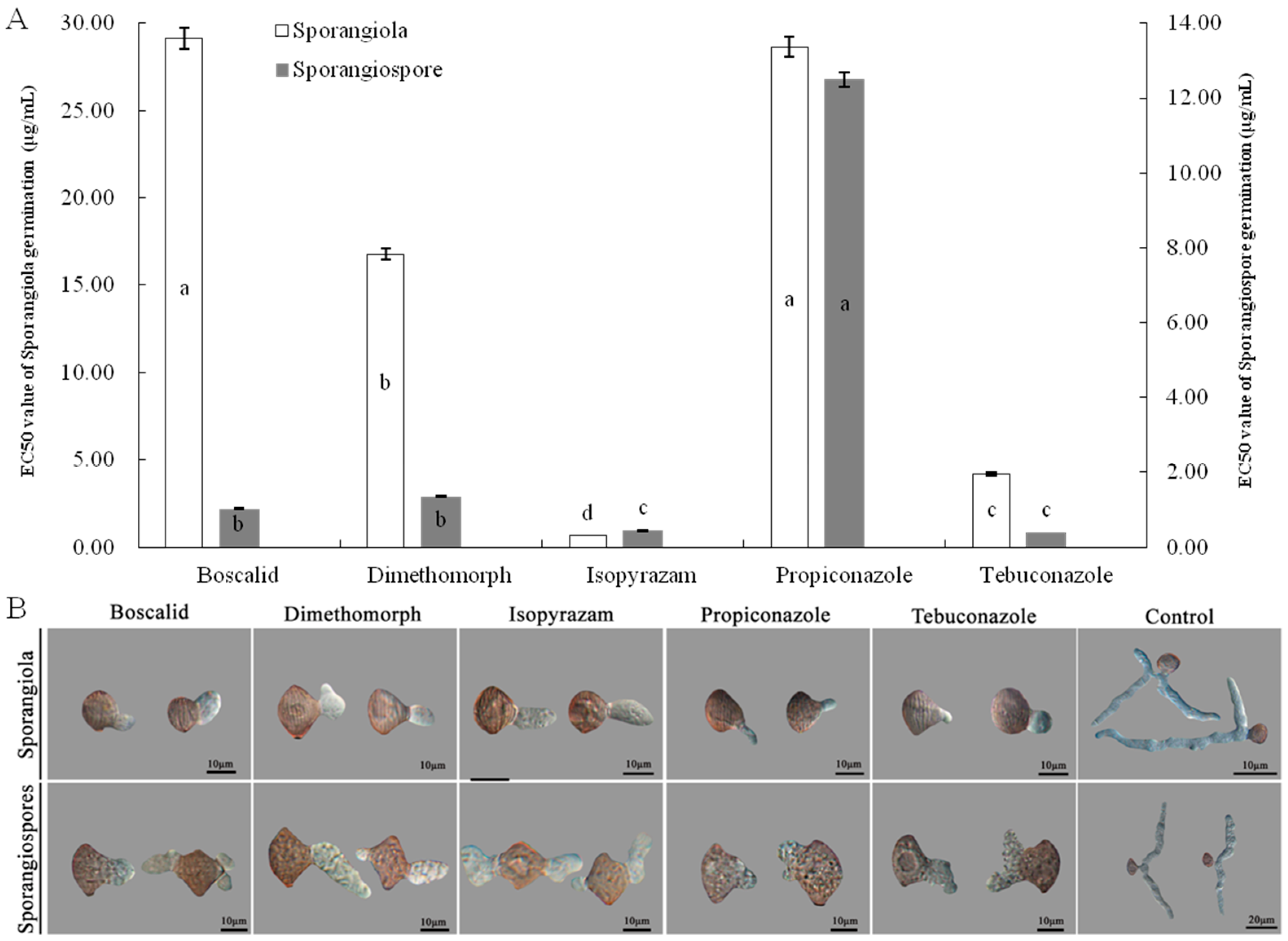
<p>Effects of five fungicides on germination of sporangiola and sporangiospores of isolate LMJM-2. (<b>A</b>) Inhibition activities of five fungicides on germination of sporangiola and sporangiospores, (<b>B</b>) morphology of germ tubes of sporangiola and sporangiospores treated with five fungicides at EC<sub>50</sub>. Different letters indicate significant differences (<span class="html-italic">p</span> = 0.05).</p> ">
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Pathogen Isolation
2.2. Morphological Analysis
2.3. Molecular Identification
2.4. Sporangiola and Sporangiospore Germination
2.5. Pathogenicity Tests
2.6. Sensitivity of Sporangiola and Sporangiospore Germination to Five Fungicides
2.7. Data Statistics and Analysis
3. Results
3.1. Field Symptoms
3.2. Morphological Characteristiscs of the Choanephora cucurbitarum
3.3. Phylogenetic Analysis of the Choanephora cucurbitarum
3.4. Sporangiola and Sporangiospore Germination of Choanephora cucurbitarum
3.5. Pathogenicity Analysis of Isolates LMJM-2, LMJM-3, LMJM-5, LMJM-7, and LMJM-9
3.6. Effect of Temperature on the Pathogenicity of Choanephora cucurbitarum
3.7. Effect of Five Fungicides on Spore Germination of Isolate LMJM-2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huai, K.H.Y.; Pei, S.J. Wild plants in the diet of arhorchin mongol herdsmen in inner mongolia. Econ. Bot. 2000, 54, 528–536. [Google Scholar] [CrossRef]
- Vega-Gálvez, A.; Miranda, M.; Vergara, J.; Uribe, E.; Puente, L.; Martínez, E.A. Nutrition facts and functional potential of quinoa (Chenopodium quinoa Willd.), an ancient Andean grain: A review. J. Sci. Food Agric. 2010, 90, 2541–2547. [Google Scholar] [CrossRef]
- Yang, B.Y.; Cheng, M.H.; Ko, C.H.; Wang, Y.N.; Chen, W.H.; Hwang, W.S.; Yang, Y.C.; Chen, H.T.; Chang, F. Potential bioethanol production from taiwanese chenopods (Chenopodium formosanum). Energy 2014, 76, 59–65. [Google Scholar] [CrossRef]
- Mangelson, H.; Jarvis, D.E.; Mollinedo, P.; Rollano-Penaloza, O.M.; Palma-Encinas, V.D.; Gomez-Pando, L.R.; Jellen, E.N.; Maughan, P.J. The genome of Chenopodium pallidicaule: An emerging andean super grain. Appl. Plant Sci. 2019, 7, e11300. [Google Scholar] [CrossRef] [PubMed]
- Chamkhi, I.; Charfi, S.; EI Hachlafi, N.; Mechchate, H.; Guaouguaou, F.E.; EI Omari, N.; Bakrim, S.; Balahbib, A.; Zengin, G.; Bouyahya, A. Genetic diversity, antimicrobial, nutritional, and phytochemical properties of Chenopodium album: A comprehensive review. Food Res. Int. 2022, 154, 110979. [Google Scholar] [CrossRef]
- Jarvis, D.E.; Sproul, J.S.; Navarro-Domínguez, B.; Krak, K.; Jaggi, K.; Huang, Y.F.; Huang, T.Y.; Lin, T.C.; Jellen, E.N.; Maughan, P.J. Chromosome-scale genome assembly of the hexaploid taiwanese goosefoot “djulis” (Chenopodium formosanum). Genome Biol. Evol. 2022, 14, evac120. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, R.; Singh, K.V. Quinoa (Chenopodium quinoa Willd.), functional superfood for today’s world: A review. World Sci. News 2016, 58, 84–96. [Google Scholar]
- Pellegrini, M.; Lucas-Gonzales, R.; Ricci, A.; Fontecha, J.; Fernández-López, J.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Chemical, fatty acid, polyphenolic profile, techno-functional and antioxidant properties of flours obtained from quinoa (Chenopodium quinoa Willd.) seeds. Ind. Crops Prod. 2018, 111, 38–46. [Google Scholar] [CrossRef]
- Lin, P.H.; Chao, Y.Y. Different drought-tolerant mechanisms in quinoa (Chenopodium quinoa Willd.) and djulis (Chenopodium formosanum Koidz.) based on physiological analysis. Plants 2021, 10, 2279. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.S.; Qin, P.Y.; Guo, H.M.; Ren, G.X. Quinoa industry development in China. Cienc. Investig. Agrar. 2019, 46, 208–219. [Google Scholar]
- Qi, T.M.; Li, Z.J.; Qin, P.Y.; Ren, G.X.; Zhou, B.W. Research and application prospect of quinoa cultivation technology. J. Agric. Sci. Technol. 2022, 24, 157–165. [Google Scholar]
- Yin, H.; Zhou, J.B.; Chen, Y.L.; Ren, L.; Qin, N.; Xing, Y.L.; Zhao, X.J. Morphology, phylogeny, and pathogenicity of Trichothecium, Alternaria, and Fusarium species associated with panicle rot on Chenopodium quinoa in Shanxi Province, China. Plant Pathol. 2022, 71, 344–360. [Google Scholar] [CrossRef]
- Yin, H.; Chen, Y.L.; Qin, N.; Zhou, J.B.; Ren, L.; Zhao, X.J. Identification of the pathogen causing quinoa gray mold and effects of environmental factors on its mycelial growth and conidial germination. J. Plant Prot. 2022, 49, 899–908. [Google Scholar]
- Sun, S.L.; Zhu, Z.D.; Zhang, J.L.; Mei, L. Outbreak of Choanephora stem rot caused by Choanephora cucurbitarum on quinoa (Chenopodium quinoa) in China. Plant Dis. 2018, 102, 2379. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Zhou, J.B.; Lv, H.; Qin, N.; Chang, F.J.; Zhao, X.J. Identification, pathogenicity, and fungicide sensitivity of Ascochyta caulina (teleomorph: Neocamarosporium calvescens) associated with black stem on quinoa in China. Plant Dis. 2020, 104, 2585–2597. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Zhou, J.B.; Lv, H.; Chang, F.J.; Qin, N.; Zhai, S.Y.; Xing, K.; Zhao, F.; Zhao, X.J. Identification of the pathogen causing Cercospora leaf spot on quinoa. Acta Phytopathol. Sin. 2019, 49, 408–414. [Google Scholar]
- Brahmanage, R.S.; Liu, M.; Wanasinghe, D.N.; Dayarathne, M.C.; Mei, L.; Jeewon, R.; Li, X.H.; Hyde, K.D. Heterosporicola beijingense sp. nov. (Leptosphaeriaceae, Pleosporales) associated with Chenopodium quinoa leaf spots. Phytopathol. Mediterr. 2020, 59, 219–227. [Google Scholar]
- Patel, R.; Patel, D.; Pandey, A.K. First report of Alternaria alternata on Chenopodium album L. from India. Biol. Forum 2014, 6, 150–152. [Google Scholar]
- Liu, X.J.; Guo, Y.L. Studies on some species of the genus Cercosporidium in China. Actamycologicasinica 1982, 1, 88–102. [Google Scholar]
- Jiang, W.Y.; Shen, Z.B.; Cai, Y.N.; Li, Y.G.; Wang, S.; Sun, L. First report of leaf spot of Chenopodium album caused by Fusarium equiseti in China. Plant Dis. 2019, 103, 3285. [Google Scholar] [CrossRef]
- Chen, X.-Y.-L.; Zhang, C.; Yang, H.; Yang, M.-F.; Cernava, T. First report of leaf spot on Chenopodium album caused by Nigrospora pyriformis in China. Plant Dis. 2020, 104, 1872. [Google Scholar] [CrossRef]
- Liu, C.Y.; Liu, Y.L.; Xie, Z.H.; Yin, X.G.; Lu, J.N. First report of seedling rot of castor (Ricinus communis) caused by Choanephora cucurbitarum in China. Plant Dis. 2022, 106, 2521. [Google Scholar] [CrossRef]
- Roberts, P.D.; Urs, R.R.; Kucharek, T.A.; Semer, C.R.; Benny, G.L.; Pernezny, K. Outbreak of Choanephora blight caused by Choanephora cucurbitarum on green bean and pepper in Florida. Plant Dis. 2003, 87, 1149. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Cho, S.E.; Choi, I.Y.; Shin, H.D. First report of Choanephora rot of okra caused by Choanephora cucurbitarum in Korea. J. Phytopathol. 2015, 163, 503–506. [Google Scholar] [CrossRef]
- Park, J.H.; Cho, S.E.; Han, K.S.; Kim, B.S.; Shin, H.D. First report of Choanephora rot caused by Choanephora cucurbitarum on Hosta plantaginea in Korea. Plant Dis. 2014, 99, 158. [Google Scholar] [CrossRef]
- Park, J.H.; Cho, S.E.; Hong, S.H.; Shin, H.D. Choanephora flower rot caused by Choanephora cucurbitarum on Abelmoschus manihot. Trop. Plant Pathol. 2015, 40, 147–149. [Google Scholar] [CrossRef]
- Park, J.H.; Cho, S.E.; Suyama, M.; Degawa, Y.; Shin, H.D. Identification and characterization of Choanephora spp. causing Choanephora flower rot on Hibiscus syriacus. Eur. J. Plant Pathol. 2016, 146, 949–961. [Google Scholar] [CrossRef]
- Park, J.H.; Choi, I.Y.; Park, M.J.; Han, K.S.; Shin, H.D. First report of Choanephora flower blight on Dahlia pinnate caused by Choanephora cucurbitarum in Korea. Plant Dis. 2016, 100, 534. [Google Scholar] [CrossRef]
- Choi, I.Y.; Kim, J.H.; Park, J.H.; Cho, S.E.; Shin, H.D. First report of Choanephora flower blight caused by Choanephora cucurbitarum on Althaea officinalis in Korea. Plant Dis. 2016, 100, 1953. [Google Scholar] [CrossRef]
- Wang, F.F.; Tang, T.; Mao, T.; Guo, J.; Duan, Y.Y.; Guo, X.L.; Yuan, B.; You, J.M. First report of blight on Pinellia ternata (banxia) caused by Choanephora cucurbitarum in China. Plant Dis. 2021, 105, 4165. [Google Scholar] [CrossRef]
- Abdel-Motaal, F.F.; El-Sayed, M.A.; El-Zayat, S.A.; Nassar, M.S.M.; Ito, S. Choanephora rot of floral tops of Hyoscyamus muticus caused by Choanephora cucurbitarum. J. Gen. Plant Pathol. 2010, 76, 358–361. [Google Scholar] [CrossRef]
- Kagiwada, S.; Kayano, Y.; Hoshi, H.; Kawanishi, T.; Oshima, K.; Hamamoto, H.; Horie, H.; Namba, S. First report of Choanephora rot of ice plant (Mesembryanthemum crystallinum) caused by Choanephora cucurbitarum in Japan. J. Gen. Plant Pathol. 2010, 76, 345–347. [Google Scholar] [CrossRef]
- Oliveira, G.S.; Fernandes, R.A.S.; Pinho, D.B.; Bonaldo, S.M. First report of Choanephora cucurbitarum causing leaf wilt, flower rot, and stem necrosis on Crotalaria breviflora. Plant Dis. 2021, 105, 1562. [Google Scholar] [CrossRef]
- Ryu, H.; Hong, S.; Choi, H.-W.; Son, K.-A. First report of Choanephora rot on lettuce (Lactuca sativa L.) caused by Choanephora cucurbitarum in Korea. Plant Dis. 2023, 107, 1217. [Google Scholar] [CrossRef]
- Kwon, J.H.; Jee, H.J. Soft rot of eggplant (Solanum melongena) caused by Choanephora cucurbitarum in Korea. Mycobiology 2005, 33, 163–165. [Google Scholar] [CrossRef]
- He, Y.Y.; Chen, X.Q.; Huang, M.Z.; Gao, X.; Zhang, Z.B.; Zhang, C.L.; Tian, Y.; Li, F. First report of seed pod rot of Moringa oleifera caused by Choanephora cucurbitarum in China. Plant Dis. 2017, 101, 1824. [Google Scholar] [CrossRef]
- Liu, P.Q.; Wei, M.Y.; Zhang, J.Z.; Wang, R.B.; Li, B.J.; Chen, Q.H.; Weng, Q.Y. Leaf blight on okra caused by Choanephora cucurbitarum in China. Can. J. Plant Pathol. 2019, 41, 366–371. [Google Scholar] [CrossRef]
- Hossain, M.A.; Munshi, A.R.; Hossen, M.S.; Rahman, K.Z.; Karim, M.R.; Kimura, Y. Morpho-molecular characterization of causative agents of wilting, leaf spot, fruit blight and stem canker of okra (Abelmoschus esculentus L.). Arch. Phytopathol. Plant Prot. 2021, 54, 1501–1518. [Google Scholar] [CrossRef]
- Pornsuriya, C.; Chairin, T.; Thaochan, N.; Sunpapao, A. Choanephora rot caused by Choanephora cucurbitarum on Brassica chinensis in Thailand. Australas. Plant Dis. Notes 2017, 12, 13. [Google Scholar] [CrossRef]
- Holcomb, G.E. First report of Choanephora flower spot and blight of Periwinkle. Plant Dis. 1998, 82, 447. [Google Scholar] [CrossRef]
- Zheng, X.R.; Chen, J.J. First report of leaf spot on pecan caused by Choanephora cucurbitarum in China. Plant Dis. 2023, 107, 2241. [Google Scholar] [CrossRef]
- Alfenas, R.F.; Bonaldo, S.M.; Fernandes, R.A.S.; Colares, M.R.N. First report of Choanephora cucurbitarum on Crotalaria spectabilis: A highly aggressive pathogen causing a flower and stem blight in Brazil. Plant Dis. 2018, 102, 1456. [Google Scholar] [CrossRef]
- Garcia-Estrada, R.S.; Rivera-Salas, M.M.; Marquez-Zequera, I.; Osuna-Garcia, L.A.; Felix-Arellano, V.; Castro-Alvarado, L.; Cruz-Lachica, I. First report of Cucurbita blossom blight and fruit rot caused by Choanephora cucurbitarum in Mexico. Plant Dis. 2023. [Google Scholar] [CrossRef]
- Žerjav, M.; Schroers, H.-J. First report of Cucurbita fruit rot caused by Choanephora cucurbitarum in Slovenia. Plant Dis. 2019, 103, 760. [Google Scholar] [CrossRef]
- Holcomb, G.E. First report of Petunia blight caused by Choanephora cucurbitarum in the United States. Plant Dis. 2003, 87, 751. [Google Scholar] [CrossRef]
- Saroj, A.; Kumar, A.; Qamar, N.; Alam, M.; Singh, H.N.; Khaliq, A. First report of wet rot of Withania somnifera caused by Choanephora cucurbitarum in India. Plant Dis. 2012, 96, 293. [Google Scholar] [CrossRef]
- Xu, J.Q.; Yan, G.F.; Tian, J.; Che, Z.P.; Kan, Y.H. Effects of carbendazim, tebuconazole and azoxystrobin on sporulation and conidial germination of Cladosporium paeoniae causing tree peony red spot. J. Plant Prot. 2016, 43, 850–857. [Google Scholar]
- Sun, G.Y.; Zhang, R.; Zhang, Z.; Zhang, M. Isolation of sooty blotch and flyspeck fungi from apple surface by picking up the thalli. Acta Phytopathol. Sin. 2003, 33, 479–480. [Google Scholar]
- Yaegashi, H.; Kanematsu, S.; Ito, T. Molecular characterization of a new hypovirus infecting a phytopathogenic fungus, Valsa ceratosperma. Virus Res. 2012, 165, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.H.; Zhang, L.; Yang, X.X.; Huang, X.Q.; Wu, W.X.; Zhou, X.Q.; White, J.F.; Liu, Y.; Li, C.J. Characterization, phylogenetic analyses, and pathogenicity of Colletotrichum species on Morus alba in Sichuan Province, China. Plant Dis. 2019, 103, 2624–2633. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Huang, Y.F.; Wang, H.C.; Chen, Q.Y.; Wang, J.; Zhang, C.Q.; Lu, H.X. Inhibitory activities of six fungicides against mycelial growth and conidial germination of Alternaria alternate. Chin. J. Pestic. Sci. 2016, 18, 263–267. [Google Scholar]
- Tang, X.L.; Yangjing, G.S.; Zhuoma, G.S.; Guo, X.F.; Cao, P.X.; Yi, B.L.; Wang, W.M.; Ji, D.; Pasquali, M.; Baccelli, I.; et al. Biological characterization and in vitro fungicide screenings of a new causal agent of wheat Fusarium head blight in Tibet, China. Front. Microbiol. 2022, 13, 941734. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhou, X.X.; Wei, S.H.; Huang, H.J.; Lan, Y.N.; Li, W.Y.; Sun, Y.; Huang, Z.F. Multiple resistance to ALS-inhibiting and PPO-inhibiting herbicides in Chenopodium album L. from China. Pestic. Biochem. Physiol. 2022, 186, 105155. [Google Scholar] [CrossRef] [PubMed]
- Walther, G.; Pawłowska, J.; Alastruey-Izquierdo, A.; Wrzosek, M.; Rodriguez-Tudela, J.L.; Dolatabadi, S.; Chakrabarti, A.; de Hoog, G.S. DNA barcoding in mucorales: An inventory of biodiversity. Persoonia 2013, 30, 11–47. [Google Scholar] [CrossRef]
- Hyde, K.D.; Nilsson, R.H.; Alias, S.A.; Ariyawansa, H.A.; Blair, J.E.; Cai, L.; de Cock, A.W.A.M.; Dissanayake, A.J.; Glockling, S.L.; Goonasekara, I.D.; et al. One stop shop: Backbones trees for important phytopathogenic genera: I (2014). Fungal Divers. 2014, 67, 21–125. [Google Scholar] [CrossRef]
- Barnett, H.L.; Lilly, V.G. The effects of humidity, temperature and carbon dioxide on sporulation of Choanephora cucurbitarum. Mycologia 1955, 47, 26–29. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host | Disease | Country | References |
|---|---|---|---|
| Abelmoschus esculentus | Blossom blight | Korea | [24] |
| Leaf blight | China | [37] | |
| Pod soft rot | Korea, Bangladesh | [24,38] | |
| Stem canker | Bangladesh | [38] | |
| A. manihot | Blossom blight | Korea | [26] |
| Flower wet rot | |||
| Althaea officinalis | Flower blight | Korea | [29] |
| Brassica chinensis | Leaf wet rot | Thailand | [39] |
| Capsicum annuum | Blossom blight | United States | [23] |
| Fruit soft rot | |||
| Leaf blight | |||
| Shoot tip dieback | |||
| Catharanthus roseus | Flower blight | United States | [40] |
| Carya illinoinensis | Leaf spot | China | [41] |
| Crotalaria spectabilis | Flower blight | Brazil | [42] |
| Stem blight | |||
| Cucurbita pepo | Blossom blight | Mexico | [43] |
| Fruit soft rot | Slovenia | [44] | |
| C. moschata | Blossom blight | Slovenia | [44] |
| Fruit soft rot | |||
| C. quinoa | Stem rot | China | [14] |
| Crotalaria breviflora | Flower rot | Brazil | [33] |
| Leaf wilt | |||
| Stem necrosis | |||
| Dahlia pinnata | Flower blight | Korea | [28] |
| Hibiscus syriacus | Flower rot | Korea | [27] |
| Hosta plantaginea | Flower wet rot | Korea | [25] |
| Hyoscyamus muticus | Floral tops rot | Japan | [31] |
| Lactuca sativa | Leaf rot | Korea | [34] |
| Mesembryanthemum crystallinum | Leaf rot | Japan | [32] |
| Stem rot | |||
| Moringa oleifera | Seed pod rot | China | [36] |
| Petunia hybrida | Flower blight | United States | [45] |
| Flower wet rot | |||
| Phaseolus vulgaris | Blossom blight | United States | [23] |
| Fruit soft rot | |||
| Leaf blight | |||
| Shoot tip dieback | |||
| Pinellia ternata | Flower blight | China | [30] |
| Leaf rot | |||
| Stem rot | |||
| Ricinus communis | Seedling rot | China | [22] |
| Solanum melongena | Soft rot | Korea | [35] |
| Withania somnifera | Leaf wet rot | India | [46] |
| Stem wet rot |
| Species | Strain Number | GenBank Accession Number | |
|---|---|---|---|
| LSU | ITS | ||
| C. cucurbitarum | LMJM-2 | OR002181 | OR002157 |
| LMJM-3 | OR002182 | OR002158 | |
| LMJM-5 | OR002183 | OR002159 | |
| LMJM-7 | OR002184 | OR002160 | |
| LMJM-9 | OR002185 | OR002161 | |
| CBS 178.76T | MT523842 | JN206235 | |
| CBS 674.93 | JN939195 | JN206233 | |
| JPC1 | MH041504 | MH041502 | |
| JSAFC2346 | OP315251 | OP315248 | |
| JSAFC2347 | OP315252 | OP315249 | |
| JSAFC2348 | OP315253 | OP315250 | |
| KA47637 | KJ461160 | KJ461159 | |
| KA47639 | KJ461162 | KJ461161 | |
| KUS-F27485 | KR867729 | KR867728 | |
| KUS-F27538 | KP726892 | KP726891 | |
| KUS-F27540 | KM200035 | KM200034 | |
| KUS-F27657 | KR867731 | KR867730 | |
| KUS-F28029 | KT581013 | KT581012 | |
| KUS-F28066 | KP406600 | KP406599 | |
| KUS-F29113 | KU316935 | KU316934 | |
| QJFY1 | MW341527 | MW295532 | |
| C. infundibulifera | CBS 153.51 | JN939193 | JN206236 |
| KUS-F27535 | KJ486538 | KJ486539 | |
| KUS-F28085 | KR867733 | KR867732 | |
| KUS-F28106 | KR867735 | KR867734 | |
| Blakeslea trispora | CBS 564.91T | JN206515 | JN206230 |
| Fungicide | Concentration (μg/mL) | |
|---|---|---|
| Sporangiola Germination | Sporangiospore Germination | |
| Tebuconazole | 0.5, 1, 5, 10, 20 | 0.1, 0.15, 0.2, 0.5, 1 |
| Propiconazole | 1, 25, 50, 75, 100 | 5, 7.5, 10, 15, 20 |
| Boscalid | 10, 15, 20, 40, 60 | 0.5, 0.75, 1, 1.5, 2 |
| Isopyrazam | 0.1, 0.5, 1, 1.5, 2 | 0.1, 0.25, 0.35, 0.5, 1 |
| Dimethomorph | 10, 15, 20, 25, 30 | 0.5, 0.75, 1, 2, 2.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, H.; Tian, M.; Peng, Y.; Qin, N.; Lü, H.; Ren, L.; Zhao, X. First Report on Choanephora cucurbitarum Causing Choanephora Rot in Chenopodium Plants and Its Sensitivity to Fungicide. J. Fungi 2023, 9, 881. https://doi.org/10.3390/jof9090881
Yin H, Tian M, Peng Y, Qin N, Lü H, Ren L, Zhao X. First Report on Choanephora cucurbitarum Causing Choanephora Rot in Chenopodium Plants and Its Sensitivity to Fungicide. Journal of Fungi. 2023; 9(9):881. https://doi.org/10.3390/jof9090881
Chicago/Turabian StyleYin, Hui, Miao Tian, Yufei Peng, Nan Qin, Hong Lü, Lu Ren, and Xiaojun Zhao. 2023. "First Report on Choanephora cucurbitarum Causing Choanephora Rot in Chenopodium Plants and Its Sensitivity to Fungicide" Journal of Fungi 9, no. 9: 881. https://doi.org/10.3390/jof9090881