


Biomedicines, Volume 9, Issue 5 (May 2021) – 140 articles
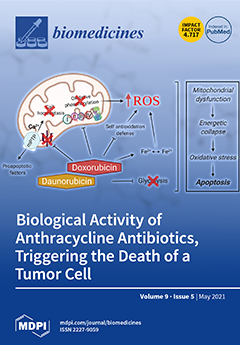
Cover Story (view full-size image):
Anthracycline antibiotics—i.e., doxo and daunorubicin—deactivate respiratory chain complexes and disrupt calcium homeostasis, as well as the self-antioxidant defense system, leading to the overproduction of free radicals, plus mPTP formation, which promotes the proapoptotic factor release into cell cytosol and triggers the apoptosis cascade. In addition, they inhibit the glycolytic energy production pathway in tumor cells, thereby exhibiting cytotoxicity.View this paper
- Issues are regarded as officially published after their release is announced to the table of contents alert mailing list.
- You may sign up for e-mail alerts to receive table of contents of newly released issues.
- PDF is the official format for papers published in both, html and pdf forms. To view the papers in pdf format, click on the "PDF Full-text" link, and use the free Adobe Reader to open them.
Previous Issue
Next Issue