


Vaccines, Volume 8, Issue 2 (June 2020) – 196 articles
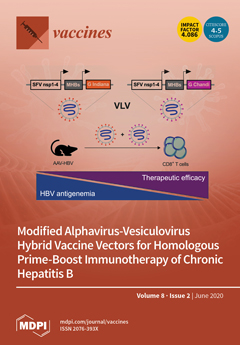
Cover Story (view full-size image):
Virus-like vesicles (VLV) are a hybrid virus-based vaccine platform composed of Semliki Forest virus nonstructural proteins (NSP) and vesicular stomatitis virus glycoprotein (G). Using dual subgenomic promoters and an envelope glycoprotein switch, prime-boost immunization with VLV expressing the hepatitis B virus (HBV) middle S protein (MHBs) showed enhanced immunogenicity and efficacy in a chronic HBV mouse model based on HBV genome delivery to the liver with adeno-associated virus (AAV). Mice with lower or intermediate HBV antigen levels had a sustained reduction of HBV following VLV prime-boost immunization. However, mice with higher HBV antigen levels showed no changes in HBV persistence, emphasizing the importance of HBV antigenemia for implementing immunotherapies against chronic hepatitis B. View this paper.
- Issues are regarded as officially published after their release is announced to the table of contents alert mailing list.
- You may sign up for e-mail alerts to receive table of contents of newly released issues.
- PDF is the official format for papers published in both, html and pdf forms. To view the papers in pdf format, click on the "PDF Full-text" link, and use the free Adobe Reader to open them.
Previous Issue
Next Issue