


Antioxidants, Volume 11, Issue 12 (December 2022) – 196 articles
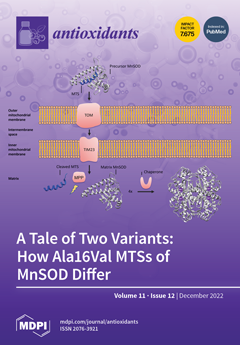
Cover Story (view full-size image):
Manganese superoxide dismutase (MnSOD) controls oxidative stress in mitochondria by scavenging reactive oxygen species (ROS). A single nucleotide polymorphism (SNP) on its gene has been linked to various types of cancer and autoimmune diseases. This study determines the effect of the SNP on the secondary structure of the mitochondrial targeting sequence (MTS) of the MnSOD protein using molecular dynamics simulations in the CHARMM36m force field. Our findings show that the alanine variant of the MTS maintains an α-helical structure that is favorable for protein transport into mitochondria, whereas the valine variant's α-helix breaks down. These results suggest that the alanine MTS may be the more active variant of MnSOD, though further experiments are needed to confirm this. View this paper
- Issues are regarded as officially published after their release is announced to the table of contents alert mailing list.
- You may sign up for e-mail alerts to receive table of contents of newly released issues.
- PDF is the official format for papers published in both, html and pdf forms. To view the papers in pdf format, click on the "PDF Full-text" link, and use the free Adobe Reader to open them.
Previous Issue
Next Issue