


Int. J. Mol. Sci., Volume 19, Issue 6 (June 2018) – 276 articles
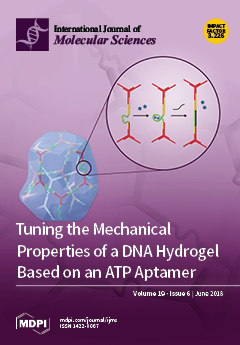
Cover Story (view full-size image):
A Y-scaffold (red line) and linear linker (yellow line) were designed to form a DNA hydrogel by hybridization of the reactive ends. An adenosine triphosphate (ATP) aptamer sequence (green line) was inserted into the linker region. On addition of ATP (blue star), the conformational transition of the aptamer had a great influence on the mechanical properties of the DNA hydrogel. The mechanical properties can be further tuned by adding a fully complementary strand (black line) of the aptamer. View Paper here.
- Issues are regarded as officially published after their release is announced to the table of contents alert mailing list.
- You may sign up for e-mail alerts to receive table of contents of newly released issues.
- PDF is the official format for papers published in both, html and pdf forms. To view the papers in pdf format, click on the "PDF Full-text" link, and use the free Adobe Reader to open them.
Previous Issue
Next Issue