1. Introduction
The European hake (Merluccius merluccius) belongs to the Merlucciidae family and is present in the North-East Atlantic, as well as the Mediterranean Sea with the bathymetric range from 25 to 1,000 m, dwelling near the bottom during daytime and moving to shallow waters at night to feed [1]. The species has been described as one of the most important demersal species both ecologically and commercially in the Mediterranean Sea [2, 3]. European hake is a species with a rather long lifespan that can reach more than 10 years of age [4], with the maximum reported age of 20 years [2]. Females prevail in sizes larger than 30 cm and make up the entire population at 38–40 cm [5]. The species is a multiple batch spawner with indetermined fecundity and a pelagic deposition [6]. Reproduction occurs throughout the year, with intensity and peaks varying among regions. Larger individuals exhibit delayed maturity in colder, nutrient-rich waters like the Atlantic compared to warmer waters of the Mediterranean Sea, and particularly the Adriatic [7].
In both Atlantic and Mediterranean waters, the European hake appears to be overfished or to have reached its maximum potential [8]. Reports throughout the Mediterranean including Algeria [9], Turkish central Aegean Sea [10], Sea of Marmara [11], Turkish northern Aegean Sea [12], and the Atlantic Moroccan coasts [13] indicate that hake populations appear to be heavily exploited or overexploited. Hake landings in Hellenic waters between 1992 and 2022 exhibited a declining trend, with an annual mean of 4,107 ± 889 tons [14] with Aegean catches representing on average (2010–2020) 81% the of the total Hellenic catches [15]. A fact of concern is that numerous reports indicate that the length at first maturity (L50) is higher than 20 cm [10, 16–23] which is the minimum landing size (MLS) in the Mediterranean, whereas the MLS in effect for hake populations of the Atlantic and North Sea is between 27 and 30 cm [24].
This study aims to update information on the present state of European hake’s biology in Hellenic waters, namely the population structure, growth, mortality, reproduction, and exploitation rate to contribute to the improvement of stock assessment and the facilitation of sustainable management of this commercially important species and one of the most exploited demersal species throughout the Mediterranean.
2. Materials and methods
2.1. Study area and sampling
A total of 861 individuals (413 males and 448 females) were collected using commercial bottom trawls, between May 2021 and October 2022 from the Aegean Sea (the Eastern Mediterranean) (north Euboean gulf, 38° 47′ 12" Ν 23° 08′ 12" Ε; Pagasitikos gulf 39° 14′ 08" Ν 23° 00′ 42" Ε; central Aegean Sea 39° 01′ 41" Ν 24° 19′ 48" Ε) (Figure 1). The bottom trawl used was a traditional Greek commercial trawl with a square mesh codend with a mesh size (bar length) of 28 mm (stretched). The sampling depth ranged between 80 and 200 m. Mean annual water temperature ranged between 14 and 16°C, salinity ranged at 37–39, pH between 8.40 and 8.46, and oxygen concentration between 5.3 and 5.9 mg/ℓ. Turbidity ranged between 17.4 and 19.8 m. Total length (TL) and total weight (TW) were measured to the nearest 0.01 cm and 0.01 g, respectively. The gonads and otoliths were extracted for further analysis.
2.2. Growth parameters
Growth parameters were calculated using the von Bertalanffy growth equation [25]:
where k (growth coefficient) is the rate at which the asymptotic length, L∞, is approached, t is the age in years, and t0 is the hypothetical age at which the fish has zero length.
The index of growth (in length) performance [26] was derived using the von Bertalanffy parameters:
The maximum lifespan was estimated according to [27] as:
2.3. Mortality and exploitation rate
Total mortality (Z) was calculated using Beverton and Holt’s [28] empirical equation:
where Lmean is the mean length of all fish in a sample representing a steady-state population and L′ is the cut-off length or the lower limit of the smallest length class included in the computation.
Natural mortality was calculated using Pauly’s [29] empirical equation:
Fishing mortality (F) was calculated by subtracting natural from total mortality according to [30]:
The exploitation rate (a measure of the amount of fish that are caught from a population each year) was calculated as the ratio of fishing mortality to the total mortality [31]:
The balance between population growth and mortality was assessed from the ratio of total mortality to the growth coefficient [13]:
The length class with the highest biomass (Lopt) (optimum length) was calculated according to [32]:
The length class at which the fish population can achieve its maximum sustainable yield (MSY) (eumetric length) (Le) was calculated according to [27, 33]:
The length of first capture (Lc) was the length at initial capture (smallest individual captured). Probability of capture was estimated at 25%, 50%, and 75% level by the linear regression-derived ascending data points of the selectivity curve [34].
2.4. Condition factor
Fulton’s condition factor (K) was used as an estimate of body condition in fish, calculated according to [35]:
2.5. Reproduction
Sex and maturity stage were determined macroscopically for each specimen according to [36] as: I: Virgin; II: Virgin developing; III: Immature; IV: Maturing; V: Mature VI: Spent. Each gonad was removed and was weighed fresh after removal of excess water. The reproductive cycle was assessed seasonally with the use of the gonad-somatic index (GSI) calculated according to [37]:
The length at which 50% of individuals in a population have reached sexual maturity is known as the onset of sexual maturity (L50). To model the link between fish length and the likelihood of sexual maturity, a probit regression model utilizing maximum likelihood estimation based on a linear transformation of a sigmoid type of curve [38] was fitted to the data. Maturity stages I through III were categorized as immature, and stages IV through VI as mature.
2.6. Data analysis
The null hypothesis of no significant temporal (season) and phyletic (sex) effect on the Fulton’s condition factor or their interaction was tested with the ANOVA General Linear Model (GLM) using a least squares regression approach [39]. The chi-square goodness-of-fit test [40] was used to assess the null hypothesis of equal proportions between the male and female ratio and compare our findings with the published literature. Statistical analysis was performed with Jamovi (ver. 2.4.6) [41], at an alpha level of 0.05.
The length-weight relationship was calculated separately for females, males, and combined sex according to [42] by fitting the exponential curve (13) to the data, where TW is the total weight (g), TL is the total length (cm), a (growth factor) the intercept of the curve, and b the slope (allometry coefficient).
The Student’s t-test was used to check the isometry (b = 3) or allometry (b ≠ 3) of the length–weight relationships.
Age classes were initially estimated by length frequency distribution using Bhattacharya’s method [43], with the software FiSAT II (FAO, Rome, Italy) (version 1.2.2.) [44]. Results were refined using the maximum likelihood concept (NORMSEP) [45] separating normally distributed components of size-frequency samples (modal progression analysis). To verify the estimated age classes, 100 otoliths were randomly selected, and annual rings were read by three independent readers according to [46].
Yield and stock predictions (Relative Y/R and B/R analysis: Knife-edge selection) were performed using the relative yield-per-recruit model based on the Beverton and Holt model [47]. Biological reference points including F0.1, FMSY, and FMAX were estimated with the open-source software YPRLEN (V2.1.0) (NOAA Fisheries Integrated Toolbox, Silver Spring, MD, USA).
3. Results
3.1. Population characteristics
Sex ratio
The observed male to female ratio (0.92:1) did not significantly depart from the female to male ratio of 1:1 (X2 = 1.42, P > 0.05) or from the reported male to female ratio (0.89:1) [10] in the central Aegean Sea in the Turkish coast (X2 = 0.266, P > 0.05).
Length–weight relationship
The length–weight relationships for the total population and males and females, respectively, are shown in Figure 2.
Figure 2
Weight vs length for the entire M. merluccius population sampled (equations and associated coefficient of determination for the total population, and each sex are shown).
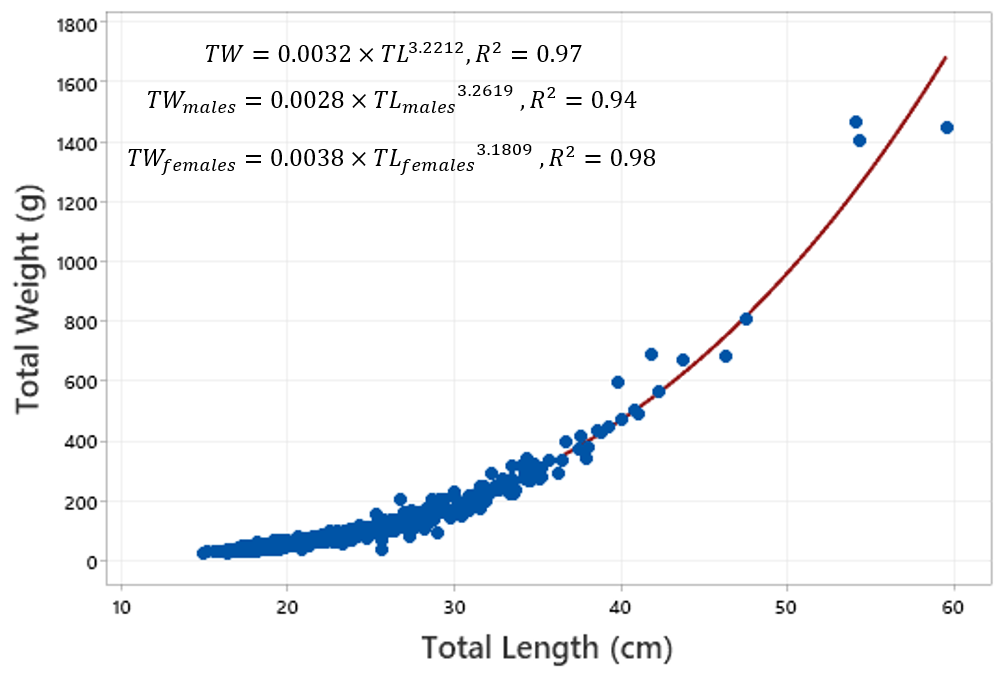
Slopes ranged between 3.181 and 3.262, exhibiting positive allometric growth. The t-test confirmed a highly significant difference from isometry for the total population and for each sex (t-test, P < 0.001).
Age composition
The age composition of the entire population is shown in Table 1.
The length-age equation that resulted from the otoliths measured was:
In total seven age classes were identified, with the most dominant being the second age class (45.6% of the total population) (Table 1).
Table 1
Population characteristics of the age groups identified for the entire M. merluccius population (mean length, standard deviation, and population percentage, at each age class)
| Age class | Total length (cm) | Standard deviation | Population % |
|---|---|---|---|
| 1 | 18.20 | 0.88 | 18.41 |
| 2 | 20.66 | 1.54 | 45.56 |
| 3 | 25.79 | 2.47 | 28.34 |
| 4 | 32.29 | 1.19 | 3.63 |
| 5 | 35.14 | 0.68 | 2.26 |
| 6 | 38.26 | 1.83 | 1.42 |
| 7 | 41.96 | 0.95 | 0.38 |
Condition factor
The Fulton’s condition factor exhibited the highest values during winter for the males and during winter and spring for females. The general linear model analysis of variance indicated that both season and sex and their interaction have a significant effect of the condition factor, with season exhibiting the greater effect (F-value 164.78, P < 0.001) followed by sex (F-value 17.19, P < 0.001) and their interaction (F-value 7.99, P < 0.001).
Reproduction
The highest reproductive intensity was exhibited during spring followed by winter (Figure 3).
Six distinct stages of gonadal development were identified in this study.

The first stage (immature) was identified at the 8.7% of the population, with the highest abundance during autumn followed by spring. The second stage was identified at 54.3% of the population, with the highest abundance during autumn followed by winter. The third stage was identified at 19.6% of the population, with the highest abundance during winter followed by autumn. The fourth stage was identified at 5.1% of the population, with the highest abundance during winter followed by autumn. The fifth stage was identified at 3.0% of the population, with the highest abundance during spring followed by autumn. The sixth stage was identified at 4.2% of the population, with the highest abundance during summer followed by spring.
All maturity stages were identified throughout every season, apart from stage V (spawning stage) which was not found during summer.
The onset of sexual maturity (L50) for the total population was estimated at 27.58 cm in total length (3.2 years) and 25.41 (2.6 years) and 30.97 (4.0 years) for males and females, respectively (Figure 4). The inflection point was estimated at 7.5 years for the total population (8.2 years for females and 5.2 years for males).
Length at first maturity from other relevant studies is shown in Table 2.
Growth and mortality
The von Bertalanffy growth equations for the total population and each sex were estimated as:
Growth index (Φ′) was estimated as 2.72.
Figure 4
Probit regression for the proportion of mature M. merluccius in relation to its total length (L25, L50, L75 and associated age are also indicated).
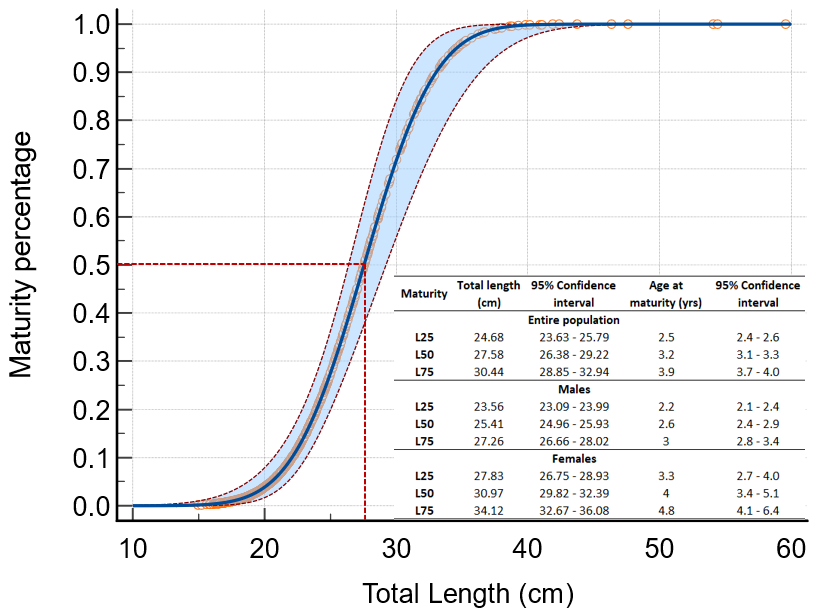
Table 2
Comparative table of length at first maturity among relevant studies
| Authors | L50 males (cm) | L50 females (cm) | Area of study |
|---|---|---|---|
| Present study | 25.41 | 30.97 | Hellenic waters |
| Khoufi et al. [19] | – | 29 | Tunisian coast |
| Soykan et al. [10] | 25.6 | 21.4 | central Aegean Sea |
| Kahraman et al. [11] | 22.5 | 29.9 | Sea of Marmara |
| Carbonara et al. [21] | – | 30.03 | Sardinia |
| 31.95 | Southern Adriatic Sea | ||
| 33.03 | South and central Tyrrhenian Sea | ||
| 32.94 | Western Ionian Sea | ||
| Candelma et al. [22] | – | 30.81 (macroscopic) 33.73 (histological) | Central Adriatic Sea |
| ELGazzar et al. [23] | – | 31.95 | Southern Adriatic |
| 34.4 | Egyptian Mediterranean |
Natural mortality (M) was estimated as 0.284, total mortality (Z) as 0.975, and fishing mortality (F) as 0.69. The exploitation rate (E) was estimated at 0.72. The length class with the highest biomass (Lopt) was estimated at 38.16 cm. The ratio of total mortality to growth coefficient (Z/k) was calculated as 7.81. Eumetric length (length class at which the fish population can achieve its maximum sustainable yield) was estimated at 37.04 cm. Longevity was estimated at 22.7 years for the total population (24.9 years for females and 16.1 years for males).
Growth and mortality parameters from other relevant studies are shown in Table 3.
Probability of capture
Probability of capture was estimated at 25% (LC25), 50% (LC50), and 75% (LC75) levels as 16.7, 17.7, and 18.5 cm, respectively (Figure 5), with age at 50% probability of capture (t50) being estimated at 1.2 years.
Relative Y/R and B/R analysis: Knife-edge selection
The yield per recruit (Y/R) against the fishing mortality and exploitation rate for different levels of natural mortality are shown in Figures 6 and 7, respectively.
Results of the yield-per-recruit analysis and biological reference points are shown in Table 4.
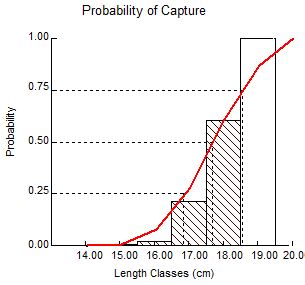
Table 3
Comparative table of growth and mortality parameters among relevant studies
| Author | Sex | L∞ | K | t0 | Φ′ | Z | M | F | E | Area of study |
|---|---|---|---|---|---|---|---|---|---|---|
| This study | Both | 65.19 | 0.125 | −1.341 | 2.72 | 0.975 | 0.284 | 0.69 | 0.72 | Hellenic waters |
| Soykan et al. [10] | Both | 54.53 | 0.315 | −0.223 | 2.97 | 1.539 | 0.579 | 0.959 | 0.62 | Central Aegean Sea |
| Kahraman et al. [11] | Both | 103.97 | 0.087 | −0.926 | 2.97 | 2.01 | 0.19 | 1.81 | 0.9 | Marmara Sea |
| Uzer et al. [12] | Both | 102.6 | 0.0992 | −0.8 | 3.01 | 2.21 | 0.57 | 1.61 | 0.72 | North Aegean Sea |
| Boudjadi and Rachedi [48] | Both | 44.08 | 0.29 | −0.517 | 2.7 | 1.53 | 0.61 | 0.92 | 0.6 | North-East of Algeria |
| El Bouzidi et al. [13] | Both | 77.39 | 0.3 | −0.427 | 3.25 | 1.49 | 0.51 | 0.98 | 0.66 | North of Moroccan Atlantic coasts |
| Slimani et al. [49] | Both | 76.65 | 0.11 | −1.21 | 2.81 | 0.65 | 0.28 | 0.37 | 0.57 | Moroccan Mediterranean coast |
Figure 6
Sensitivity analysis of yield per recruit against fishing mortality for different levels of natural mortality.
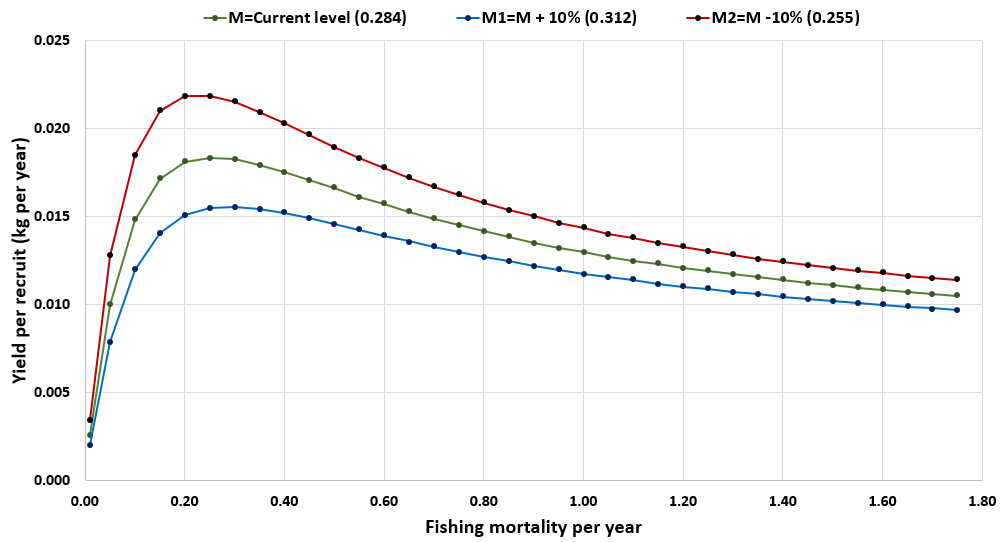
Figure 7
Sensitivity analysis for yield per recruit against exploitation rate for different levels of natural mortality (green line E0.1, red line E0.5 and yellow line Emax).

Table 4
Relative yield/recruit analysis (knife edge) and biological reference points of the population
E, exploitation rate; YPR, yield per recruit; B/R, biomass per recruit; Emax, exploitation rate which produces the maximum yield; E0.1, exploitation rate at which the marginal increase of relative yield-per-recruit is 1/10th of its value at E = 0; E0.5, value of E under which the stock has been reduced to 50% of its unexploited biomass; FMSY, level of fishing mortality that will allow a fish population to achieve MSY; FMAX, fishing mortality rate at maximum sustainable yield; F0.1, level of fishing mortality where the slope of the YPR curve is reduced to 10% of the value at the origin; FMAX, fishing effort where yield is maximum.
4. Discussion
A wide range of variables, such as temperature, salinity, habitat variation, food availability, maturity stage, fishing season, high fishing mortality, and genetic factors, affect a teleost’s sex ratio, GSI, and spawning period [50]. At smaller sizes (less than 30 cm in total length), the sex ratio favored males, but at larger sizes (more than 30 cm in total length), the sex ratio favored females. While the opposite pattern was noted in the Sea of Marmara (Turkey) by Yildiz [51], this pattern was also noticed by Piñeiro and Saínza [4], El Habouz [52], and Costa [53] in Iberian Atlantic waters, Eastern Central Atlantic, and Portuguese coast, respectively. The disparity in growth rates between sexes, the higher natural mortality rate of older males relative to females, or the overfishing-related decrease in catch availability may all contribute to the sex ratio’s unpredictability [54].
The highest reproductive intensity in this study was observed during spring followed by winter. A number of studies in the region have reported spawning between November and March, including the Mediterranean coastline in Egypt [23], Central, and Western Mediterranean [21], the Sea of Marmara Turkey [11], and the Atlantic Moroccan coast [52]. Spawning season can be affected by several factors including temperature, food availability, and the condition factor [55].
The males’ length at first maturity was comparable to that measured in the central Aegean [10]. According to [21, 52], variations in L50 could be attributed to phenotypic responses to environmental factors (such as competition, population distribution, space, and food availability) or to selective pressure, such as fishing effort, which leads to overexploitation of the stock and lower spawning biomass in various areas. The observed difference between males and females in the length at first maturity (L50) with females maturing in larger size compared to males (30.97 cm versus 25.41 cm) could be attributed to the stock’s level of exploitation [56]. The effect of growth and mortality could also lead to a higher proportion of larger-sized females if males grow at a slower rate, especially following the onset of maturity [57].
Asymptotic length (L∞) was lower than those obtained in similar studies from the Strait of Gibraltar [4], Atlantic [52], Tunisian coast [19], Sea of Marmara [20], and the North Aegean [12], with the maximum size in the Mediterranean reportedly limited to 80–90 cm [7]. Growth index (Φ′) in this study (2.7) was lower than those observed in previous studies (between 2.8 and 3.4). The growth rate for the European hake may vary according to genetic variance, biotic factors including food availability and longevity, and regional environmental differences such as temperature, salinity, and depth [58].
This study estimated a relatively high fishing mortality (F = 0.69/year) with the calculated Z/K ratio (7.81) greater than the population equilibrium (Z/K ratio = 1) where mortality is balanced by growth [59], indicating that mortality dominates growth, similarly to [13] (4.97) in the Moroccan Atlantic coast. The exploitation rate calculated in this study (E = 0.72) indicated that the population is under a high level of exploitation, indicating that the species is overfished or at best case fully exploited [60]. Similar exploitation rates were estimated in the north and central Aegean Sea along the Turkish coastline (0.72 and 0.74, respectively) [12, 61]. The optimal exploitation rate for a fish population will vary depending on the species, environment, and management practices in place. However, a general rule of thumb is that exploitation should not exceed 0.5, as this could lead to overfishing. Thus, values greater than 0.5 are an indication that the stock is overfished [30, 62].
Length at first capture (LC50) was estimated at 17.7 cm, with age at probability of capture (t50) being estimated at 1.2 years, whereas the onset of sexual maturity (L50) of the population was estimated at 27.58 cm in total length (3.2 years) and 25.41 (2.6 years) and 30.97 (4.0 years) for males and females, respectively, indicating that the population is exploited from an early age and size, well before sexual maturity. It is noteworthy that despite the minimum landing size (MLS) for M. merluccius in the Eastern Mediterranean of 20 cm total length [24], roughly 64% of the present population was comprised of 1- and 2-year old individuals, the vast majority of which with the total length lower than the MLS. Furthermore, the length class with the highest biomass in an unfished population (Lopt) estimated at 38.16 cm (greater than 5-year-old individuals in this study) comprises less than 4% of the total population, indicating that the target of capturing individuals with an average length equal to this size, theoretically allowing the stock to yield MSY level, would be unattainable.
Studies of crucial biological characteristics, namely growth [4], and the exploitation rate [63] are essential for stock assessment and development of management strategies. Results of this study indicated that further management measures need to be enforced to protect the stock and in particular the juveniles to prevent possible stock depletion. Possible measures could include the increase of MLS through alteration in mesh size, application of nets with a different geometry (square vs. diamond), seasonal closures, closed areas (spawning grounds; nursery areas), improved management of illegal and unregulated fishing (increased coastguard policing) or a combination of the above.
5. Conclusion
This study aimed to assess the population characteristics of the commercially important European hake (Merluccius merluccius) in Hellenic waters (the Eastern Mediterranean). Length at first maturity was estimated at a larger total length compared to the MLS and length at first capture with asymptotic length smaller than those observed in other studies (the Mediterranean). Results indicated that the population is overexploited with mortality dominating growth. It is recommended that further management measures can be put in place to avoid overfishing, given the importance of this highly prized commercial species.