Amoebozoa
Az Amoebozoa nagy taxonómiai csoport, mintegy 2400 amőboid protisztafajjal,[1] melyek gyakran ujjszerű, lebenyes állábakkal és csőszerű mitokondriális cristákkal rendelkeznek.[2][3] A hagyományos és jelenleg nem támogatott besorolási sémák az Amoebozoát a Protista[4] vagy a Protozoa ország törzseként sorolják be.[5] A Nemzetközi Protisztológustársaság besorolatlan „szupercsoport”-nak tartja fenn az eukariótákon belül.[2] A molekuláris genetikai analízis szerint az Amoebozoa monofiletikus, így klád. Az eukarióták filogenetikai fáiról szóló modern tanulmányok szerint az Opisthokonta, egy, az állatokat és gombákat és számos más, összesen 300 egysejtűfajt tartalmazó kládot tartalmazó csoport testvércsoportja.[1][3] Az Amoebozoa és az Opisthokonta gyakran szerepel együtt magas szintű taxonban, melyet Unikonta,[5] Amorphea[2] vagy Opimoda néven is neveznek.[6]
| Evolúciós időszak: 1000–0 Ma | ||||||
 Chaos carolinensis
| ||||||
| Rendszertani besorolás | ||||||
| ||||||
| Szinonimák | ||||||
| ||||||
| Altörzsek, altörzságak és osztályok | ||||||
| Hivatkozások | ||||||
A Wikimédia Commons tartalmaz Amoebozoa témájú kategóriát. |
Az Amoebozoa sok ismert amőboid élőlényt tartalmaz, például a Chaost, az Entamoebát, Pelomyxát és az Amoebát is. Az Amoebozoa tagjai lehetnek héjasak vagy csupaszok, és a sejtek rendelkezhetnek ostorral. Édes-, tengervízben, mohákban és avarban is előfordulnak szabadon élő fajok. Néhányuk parazitaként vagy szimbiontaként él más élőlényekben, és egyesekről ismert, hogy betegséget okoz emberekben vagy más élőlényekben.
Bár a legtöbb fajuk egysejtű, a csoportnak része néhány nyálkagombaklád, melyek többsejtű életszakasszal is rendelkeznek, mely alatt az amőboid sejtek együtt maradnak többszöri sejtosztódás után plazmódiumot alkotva, vagy a sejtes nyálkagombák esetén aggregátumot képeznek.
Fajai mérete igen széles skálán van: egyesek csak 10-20 µm-esek, míg mások a legnagyobb protozoonok közt vannak. Az Amoeba proteus, mely akár 800 μm hosszú is lehet, gyakran szerepel iskolákban és laboratóriumokban modellszervezetként például a méretük miatt. A többmagvú amőbák, például a Chaos és a Pelomyxa hossza elérheti a néhány millimétert, és néhány többsejtű faj, például a Fuligo septica nyálkagomba területe akár néhány négyzetméter is lehet.[7]
Morfológia
szerkesztés
Az Amoebozoa nagy, sokszínű csoport, de számos jellemző sok tagjában megvan. Sejtjeik általában egy szemcsés központi endoplazmára és egy tiszta külső ektoplazmára bonthatók. Mozgásuk során az endoplazma előre-, az ektoplazma hátrahalad a sejt külseje mentén. Sok fajuknak ekkor egyértelműen meghatározott elülső és hátulsó oldaluk van, és monopodiális formájúak lehetnek, ahol az egész sejt egyetlen állábként működik. A nagy állábak számos részre, úgynevezett alállábakra (szubpszeudopódium) válhatnak szét, melyek egy bizonyos hosszig kinyúlnak, majd visszahúzódnak mozgás, vagy táplálékbevitel végett. Egy sejt akár több fő állábat is képezhet, melyeken keresztül a sejt belsejének egésze a haladás irányába mozog. Ezek többé-kevésbé csőszerűek, és nagyrészt endoplazmával telítettek. A sejt tömege egy vezető állábba kerül, és a többi végül visszahúzódik, kivéve az élőlény irányváltoztatása esetén.[8]
Bár legtöbb tagjuk „csupasz”, mint például az Amoeba- vagy a Chaos-fajok, vagy laza, kis pikkelyekből álló burokba burkolóznak, mint a Cochliopodium vagy a Korotnevella, az Arcellinida rend tagjai rideg héjakat alkotnak, melyeken egyetlen nyílás van, amelyen át az állábak létrejönnek. Ezek szerves anyagokból történő szekrécióval, mint például az Arcella esetén, vagy összegyűjtött részecskék összeállásából jöhetnek létre, mint a Difflugia esetén.
Az Amoebozoa-sejtek mitokondriumai általában elágazó tubuláris cristákkal rendelkeznek. Azonban az anoxiás vagy mikroaerofil környezethez adaptált Archamoebaeben nem találhatók mitokondriumok.
Csoportosítás
szerkesztésAz Amoebozoa helye az eukarióták közt
szerkesztésA molekuláris genetika szerint az Amoebozoa az állatok és gombák testvércsoportja, mely e csoportok elválása után vált el tőlük,[9] ahogy az alábbi diagram is szemlélteti:
| Opimoda |
| ||||||||||||||||||||||||||||||||||||


Az Amoebozoa és az Opisthokonta hasonlóságai vezettek ahhoz a feltételezéshez, hogy külön kládot alkotnak.[10] Thomas Cavalier-Smith az „unikontok” (Unikonta) nevet javasolta ezen ágnak, melynek őse egy alapi testtel és az abból kiinduló ostorral rendelkeztek. Azonban bár az Amoebozoa–Opisthokonta kapcsolatot támogatják a források, a későbbi munkák szerint az egyostorúős-hipotézis hamis. A Revised Classification of Eukaryotes (2012) című munkában Adl és társai javasolták az Amorphea elnevezést egy hasonló összetételű kládra, mely a Diaphoretickes testvércsoportja.[2] A későbbi munkák az Amorpheát a Malawimonasszal és a Collodictyonidaeval helyezik egy kládba, melynek neve Opimoda, mely az egyik leghamarabb különváló rendszertani csoport – a másik a Diphoda.[6]
Altörzsek az Amoebozoában: Lobosa és Conosa
szerkesztésA hagyományos rendszertanok a lebenyes állábú Amoebozoa-tagokat a Lobosa osztályba helyezték, az ezen kívüli tagokat a Sarcodina (Rhizopoda) csoportba, de ezekről kiderült, hogy nem természetes csoportok. A szerkezeti és genetikai tanulmányok a Percolozoát és néhány Archamoebae-csoportot független csoportnak nyilvánítottak. Az rRNS-alapú törzsfák szerint ezek képviselői függetlenek a többi amőbától, és az eukarióták fejlődésének elején váltak ki, ahogy a legtöbb nyálkagomba is.
Azonban a Cavalier-Smith és Chao által 1996-ban közölt módosított törzsfák[11] szerint a Lobosa többi tagja monofiletikus csoportot alkot, melynek az Archamoebae és Mycetozoa közeli rokona, de a Percolozoa nem. Ezenkívül létrehozták az Amoebozoát, mely a Lobosa és az új Conosa altörzseket tartalmazza, mely magában foglalja az Archamoebaet és a Mycetozoát.[12]
A legújabb molekuláris genetikai adat az Amoebozoa Lobosára és Conosára való feloszthatósága mellett van.[3] Cavalier-Smith és munkatársai definíciója szerint előbbi nagyrészt a klasszikus Loboseából – ostor nélküli, lebenyes állábú amőbák (Amoeba, Acanthamoeba, Arcella, Difflungia stb.) áll. Utóbbit amőboid és ostoros sejtek egyaránt alkotják, jellemzően csúcsosabb vagy elágazó állábakkal (Archamoebae, Mycetozoa).
Filogenetika, taxonómia
szerkesztésCavalier Smith, Chao és Lewis 2016-os,[13] valamint Silar 2016-os tanulmánya alapján.[14] Szintén a legutóbbi filogenetika szerint a Lobosa parafiletikus: a Conosa a Cutosea testvércsoportja.[15][16][17]
| Az Amoebozoa törzsfája | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Amoebozoa phylum Lühe 1913 emend. Cavalier-Smith 1998 [Amoebobiota; Eumycetozoa Zopf 1884 emend Olive 1975]
- Discosea cladus Cavalier-Smith 2004 stat. nov. Adl et al. 2018
- ?Stereomyxida ordo Grell 1971
- ?Stygamoebida ordo Smirnov & Cavalier-Smith 2011
- Centramoebia classis Cavalier-Smith et al. 2016
- Centramoebida ordo Rogerson & Patterson 2002 em. Cavalier-Smith 2004
- Himatismenida ordo Page 1987 [Cochliopodiida]
- Pellitida ordo Page 1987 [Cochliopodiida]
- Flabellinia osztály Smirnov & Cavalier-Smith 2011 em. Kudryavtsev et al. 2014
- Thecamoebida ordo Schaeffer 1926 em. Smirnov & Cavalier-Smith 2011
- Dermamoebida ordo Cavalier-Smith 2004 em. Smirnov & Cavalier-Smith 2011
- Vannellida ordo Smirnov et al. 2005
- Dactylopodida ordo Smirnov et al. 2005
- Tevosa cladus Kang et al. 2017
- Tubulinea cladus Smirnov et al. 2005 stat. nov. Adl et al. 2018
- Corycidia osztály Kang et al. 2017 stat. nov. Adl et al. 2018
- Trichosida ordo Moebius 1889
- Microcoryciidae familia de Saedeleer 1934
- Echinamoebia classis Cavalier-Smith 2016 stat. nov. Adl et al. 2018
- Echinamoebida ordo Cavalier-Smith 2004 em. 2011
- Elardia classis Kang et al. 2017 stat. nov. Adl et al. 2018
- Leptomyxia subclassis Cavalier-Smith 2016
- Leptomyxida ordo Pussard & Pons 1976 em. Page 1987
- Eulobosia subclassis Cavalier-Smith 2016
- Euamoebida ordo Lepşi 1960 em. Cavalier-Smith 2016
- Arcellinida ordo Kent 1880
- Leptomyxia subclassis Cavalier-Smith 2016
- Corycidia osztály Kang et al. 2017 stat. nov. Adl et al. 2018
- Evosea cladus Kang et al. 2017 stat. nov. Adl et al. 2018
- Cutosa cladus Cavalier-Smith 2016 stat. nov.
- Cutosea cladus Cavalier-Smith 2016
- Squamocutida ordo Cavalier-Smith 2016
- Cutosea cladus Cavalier-Smith 2016
- Conosa subphylum Cavalier-Smith 1998 stat. nov.
- Archamoebae infraphylum Cavalier-Smith 1993 stat. n. 1998
- Archamoebea classis Cavalier-Smith 1983 stat. n. 2004
- Tricholimacidae familia Cavalier-Smith 2013
- Endamoebidae familia Calkins 1926
- Entamoebida ordo Cavalier-Smith 1993
- Pelobiontida ordo Page 1976 emend. Cavalier Smith 1987
- Archamoebea classis Cavalier-Smith 1983 stat. n. 2004
- Semiconosia infraphylum Cavalier-Smith 2013
- Variosea classis Cavalier-Smith et al. 2004
- ?Flamellidae ordo Cavalier-Smith 2016
- ?Holomastigida ordo Lauterborn 1895 [Artodiscida Cavalier-Smith 2013]
- Phalansteriida ordo Hibberd 1983
- Ramamoebida ordo Cavalier-Smith 2016
- Profiliida ordo Kang et al. 2017 [Protosteliida Olive & Stoianovitch 1966 em. Shadwick & Spiegel 2012]
- Fractovitellida ordo Lahr et al. 2011 em. Kang et al. 2017
- superclassis Mycetozoa ordo de Bary, 1859 ex Rostafinski, 1873
- Dictyostelea classis Hawksworth et al. 1983
- Acytosteliales ordo Baldauf, Sheikh & Thulin 2017
- Dictyosteliales ordo Lister 1909 em. Olive 1970
- Ceratiomyxomycetes classis Hawksworth, Sutton & Ainsworth 1983
- Protosporangiida ordo Shadwick & Spiegel 2012
- Ceratiomyxida ordo Martin 1961 ex Farr & Alexopoulos
- Myxomycetes classis Link 1833 em. Haeckel 1866 [18]
- Lucisporomycetidae subclassis Leontyev et al. 2019
- Cribrarianae superordo Leontyev 2015
- Cribrariales ordoMacbr. 1922
- Trichianae superordo Leontyev 2015
- Reticulariales ordo Leontyev 2015
- Liceales ordo Jahn 1928
- Trichiales ordo Macbride 1922
- Cribrarianae superordo Leontyev 2015
- Columellomycetidae subclassis Leontyev et al. 2019
- ?Echinosteliopsidales ordo Shchepin et al.
- Echinostelianae superordo Leontyev 2015
- Echinosteliales ordo Martin 1961
- Stemonitanae superordo Leontyev 2015 [Fuscisporida Cavalier-Smith 2012]
- Clastodermatales ordo Leontyev 2015
- Meridermatales ordo Leontyev 2015
- Stemonitales ordo Macbride 1922
- Physarales ordo Macbride 1922
- Lucisporomycetidae subclassis Leontyev et al. 2019
- Dictyostelea classis Hawksworth et al. 1983
- Variosea classis Cavalier-Smith et al. 2004
- Archamoebae infraphylum Cavalier-Smith 1993 stat. n. 1998
- Cutosa cladus Cavalier-Smith 2016 stat. nov.
- Tubulinea cladus Smirnov et al. 2005 stat. nov. Adl et al. 2018
Fosszíliák
szerkesztésA világszerte talált vázaalakú mikrofosszíliák (VAM) szerint az Amoebozoa a neoproterozoikum óta létezik. A fosszilis Melanocyrillium hexodiadema, Palaeoarcella athanata és Hemisphaeriella ornata mintegy 750 millió éves sziklákból ismert. Mindhárom VAM félgömb alakú, üreggel és szabályos betüremkedésekkel, melyek a modern héjas amőbákra hasonlítanak. Például a P. athanata hasonlít a létező Arcella génuszra.[19][20]
Humán patogén amőbák listája
szerkesztésMeiózis
szerkesztésAz Acanthamoeba genomszekvenciájából kiderült, hogy az ivaros eukarióták meiózisában szerepet játszó gének ortológjai vannak benne jelen. E gének közé tartoznak az Spo11, Mre11, Rad50, Rad51, Rad52, Mnd1, Dmc1, Msh és az Mlh.[21] Ez alapján lehetséges, hogy az Acanthamoeba képes meiózisra és ivaros szaporodásra.
Az ivarosan szaporodó eukariótákban a homológ rekombináció (HR) általában meiózis során történik. Egy meiózisspecifikus rekombináz, a Dmc1 szükséges a hatékony meiotikus HR-hez, és a Dmc1 kifejeződik az Entamoeba histolyticában.[22] A tisztított Dmc1 az E. histolyticából preszinaptikus szálakat alkot, és katalizálja az adenozin-trifoszfát-dependens homológ DNS-párososdást és DNS-szálcserét legalább néhányezer bázispáron át.[22] A DNS-párosodást és szálcserét segíti az eukarióta meiózisspecifikus rekombinációs segédfaktor, a Hop2-Mnd1.[22] E folyamatok fontosak a meiotikus rekombinációhoz, így feltehetően az E. histolytica esetén is történik meiózis.[22]
Az Entamoeba invadensszel foglalkozó tanulmányok szerint a tetraploid egymagvú trofozoita négymagvú cisztává válásakor a homológ rekombináció nagyobb mértékű.[23] A meiotikus rekombináció fő lépéseihez kapcsolódó funkciókkal rendelkező gének expressziója növekszik a cisztaképzéskor is.[23] Ezek, valamint az E. histolyticáról szóló tanulmányok alapján lehetséges a meiózis jelenléte az Entamoebában. Egy genetikai összehasonlítás szerint a meiózis jelen lehet minden főbb Amoebozoa-csoportban.[24]
Humán egészség
szerkesztésAz amőbiázis, más néven entamőbiázis[25][26] az Entamoeba csoportba tartozó amőbák okozta fertőzés. A tünetek Entamoeba histolytica-fertőzés esetén a leggyakoribbak. Lehet tünetmentes, de jelentkezhet enyhe vagy súlyos tünetekkel. Tünetei közé tartoznak a hasfájás, hasmenés, dizentéria vagy a súlyos, nekrózissal és perforációval jelentkező colitis. A perforáció okozhat peritonitist is. Az érintettek anémiát is kaphatnak vérveszteség miatt.[27]
Oka és diagnózisa
szerkesztésA bélfal fertőzése amőbás vérhast vagy amőbás colitist okozhat.[27] Ha a parazita a véráramba kerül, az egész testben elterjedhet, leggyakrabban a májba kerülve, ahol amőbás májtályogot okozhat.[27] Ez megtörténhet hasmenés nélkül is.[27] Az Entamoeba-ciszták akár egy hónapig is élhetnek a talajban, illetve 45 percig a körmök alatt.[27] Fontos az amőbiázis megkülönböztetése a bakteriális colitistől. A leggyakoribb vizsgálati módszer a mikroszkópos székletvizsgálat, de ez képzett mikroszkópkezelőt igényel, és nem feltétlenül megbízható a fertőzés kizárásához. E módszer azonban nem képes megkülönböztetni különböző típusokat. A leukocitózis súlyos esetekben jelen van, enyhékben nem. A pontosabb teszt az antitestteszt, de ez kezelés után is pozitív maradhat.[27]
Megelőzés és gyógyítás
szerkesztésAz amőbiázis megelőzése jellemzően a megfelelő fertőtlenítést és az ételek, italok széklettől való elválasztását jelenti. Jelenleg (2013) nincs vakcina.[27] Két kezelés van a fertőzés helyétől függően.[27] A szöveti amőbiázist metronidazollal, tinidazollal, nitazoxaniddal, dehidroemetinnel vagy klorokinnel kezelik, míg a luminális fertőzést diloxanid-furoáttal vagy dijód-hidroxikinolinnal.[27] Hogy a kezelés hatékony legyen minden amőbaállapottal szemben, több gyógyszeres kombináció is szükséges lehet. A tünetmentes kezelések nem szükségképp igényelnek kezelést, de a fertőzöttek másokra is terjeszthetik a parazitát, ami miatt szükség lehet rá.[27] A nem E. histolytica által okozott fertőzések kezelése nem feltétlenül szükséges.[27]
Epidemiológia, történet
szerkesztésAz amőbiázis a világ egészén jelen van.[28] Mintegy 480 millió ember fertőzött E. histolyticának tűnő élőlénnyel, és 40-110 000 ember hal meg amőbiázisban évente.[27] A legtöbb fertőzést azonban az E. dispar okozza.[27] Ez gyakoribb egyes területeken, de a tünetokozó esetek száma kisebb lehet a korábbi becsléseknél.[27] Az első amőbiázisos esetet 1875-ben írták le, 1891-ben pedig részletesen is leírták a betegséget, létrehozva az „amőbás vérhas” és „amőbás májtályog” fogalmakat.[27] 1913-ban a Fülöp-szigeteken beszámoltak, hogy E. histolytica-ciszták bevitele után az önkéntesek fertőzöttek lettek.[27] 1897 óta ismert, hogy legalább egy nem betegségokozó Entamoeba-faj (az Entamoeba coli) létezett, de először csak 1997-ben számolt be a WHO, hogy az E. histolytica valójában két faj volt, bár először erről 1925-ben számoltak be.[27] A már elismert E. dispar mellett ismert legalább két további hasonlóan kinéző Entamoeba-faj: az E. moshkovskii and Entamoeba bangladeshi. Ezeket a fajokat egészen 2013-ig nem különböztették meg a hasonlóságuk miatt.[27]
Gallery
szerkesztés-
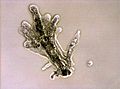
-
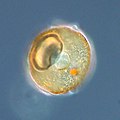 Arcella sp. test (Lobosa: Tubulinea)
Arcella sp. test (Lobosa: Tubulinea) -
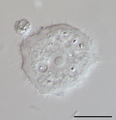 Acanthamoeba sp. (Lobosa: Discosea)
Acanthamoeba sp. (Lobosa: Discosea) -
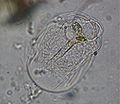 Thecamoeba sp. (Lobosa: Discosea)
Thecamoeba sp. (Lobosa: Discosea) -

-
 Pelomyxa palustris (Conosa: Archamoebae)
Pelomyxa palustris (Conosa: Archamoebae) -
 Stemonitis sp. (Conosa: Myxogastria)
Stemonitis sp. (Conosa: Myxogastria) -
 Dictyostelium discoideum (Conosa: Dictyostelia)
Dictyostelium discoideum (Conosa: Dictyostelia)








Jegyzetek
szerkesztés- ↑ a b (2012. november 6.) „CBOL protist working group: barcoding eukaryotic richness beyond the animal, plant, and fungal kingdoms”. PLOS Biology 10 (11), e1001419. o. DOI:10.1371/journal.pbio.1001419. PMID 23139639. PMC 3491025.
- ↑ a b c d (2012. szeptember 1.) „The revised classification of eukaryotes”. The Journal of Eukaryotic Microbiology 59 (5), 429–93. o. DOI:10.1111/j.1550-7408.2012.00644.x. PMID 23020233. PMC 3483872.
- ↑ a b c (2015. február 1.) „Multigene phylogeny resolves deep branching of Amoebozoa”. Molecular Phylogenetics and Evolution 83, 293–304. o. DOI:10.1016/j.ympev.2014.08.011. PMID 25150787.
- ↑ Corliss, John O. (1984. november 4.). „The Kingdom Protista and its 45 Phyla”. BioSystems 17 (2), 87–126. o. DOI:10.1016/0303-2647(84)90003-0. PMID 6395918.
- ↑ a b Cavalier-Smith, Thomas (2003. november 4.). „Protist phylogeny and the high-level classification of Protozoa”. European Journal of Protistology 39 (4), 338–348. o. DOI:10.1078/0932-4739-00002.
- ↑ a b (2015. február 1.) „Bacterial proteins pinpoint a single eukaryotic root”. Proceedings of the National Academy of Sciences of the United States of America 112 (7), E693–9. o. DOI:10.1073/pnas.1420657112. PMID 25646484. PMC 4343179.
- ↑ (2002) „Zinc accumulation by the slime mold Fuligo septica (L.) Wiggers in the former Soviet Union and North Korea”. Journal of Environmental Quality 31 (3), 1038–42. o. DOI:10.2134/jeq2002.1038. PMID 12026071.
- ↑ Jeon, Kwang W.. Biology of Amoeba. New York: Academic Press, 100. o. (1973). ISBN 978-0-323-14404-9
- ↑ Eichinger L, Pachebat JA, Glöckner G, Rajandream MA, Sucgang R, Berriman M, Song J, Olsen R, Szafranski K, Xu Q, Tunggal B, Kummerfeld S, Madera M, Konfortov BA, Rivero F, Bankier AT, Lehmann R, Hamlin N, Davies R, Gaudet P, Fey P, Pilcher K, Chen G, Saunders D, Sodergren E, Davis P, Kerhornou A, Nie X, Hall N, Anjard C, Hemphill L, Bason N, Farbrother P, Desany B, Just E, Morio T, Rost R, Churcher C, Cooper J, Haydock S, van Driessche N, Cronin A, Goodhead I, Muzny D, Mourier T, Pain A, Lu M, Harper D, Lindsay R, Hauser H, James K, Quiles M, Madan Babu M, Saito T, Buchrieser C, Wardroper A, Felder M, Thangavelu M, Johnson D, Knights A, Loulseged H, Mungall K, Oliver K, Price C, Quail MA, Urushihara H, Hernandez J, Rabbinowitsch E, Steffen D, Sanders M, Ma J, Kohara Y, Sharp S, Simmonds M, Spiegler S, Tivey A, Sugano S, White B, Walker D, Woodward J, Winckler T, Tanaka Y, Shaulsky G, Schleicher M, Weinstock G, Rosenthal A, Cox EC, Chisholm RL, Gibbs R, Loomis WF, Platzer M, Kay RR, Williams J, Dear PH, Noegel AA, Barrell B, Kuspa A (2005. május 1.). „The genome of the social amoeba Dictyostelium discoideum”. Nature 435 (7038), 43–57. o. DOI:10.1038/nature03481. PMID 15875012. PMC 1352341.
- ↑ Dawkins, Richard. The Ancestor's Tale (2016). ISBN 978-0544859937
- ↑ (1996. december 1.) „Molecular phylogeny of the free-living archezoan Trepomonas agilis and the nature of the first eukaryote”. Journal of Molecular Evolution 43 (6), 551–62. o. DOI:10.1007/BF02202103. PMID 8995052.
- ↑ (1998. augusztus 1.) „A revised six-kingdom system of life”. Biological Reviews of the Cambridge Philosophical Society 73 (3), 203–66. o. DOI:10.1111/j.1469-185X.1998.tb00030.x. PMID 9809012.
- ↑ (2016. június) „187-gene phylogeny of protozoan phylum Amoebozoa reveals a new class (Cutosea) of deep-branching, ultrastructurally unique, enveloped marine Lobosa and clarifies amoeba evolution”. Molecular Phylogenetics and Evolution 99, 275–296. o. DOI:10.1016/j.ympev.2016.03.023. PMID 27001604.
- ↑ Silar, Philippe. Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes, HAL archives-ouvertes, 1–462. o. (2016). ISBN 978-2-9555841-0-1
- ↑ (2017. szeptember 1.) „Between a Pod and a Hard Test: The Deep Evolution of Amoebae”. Molecular Biology and Evolution 34 (9), 2258–2270. o. DOI:10.1093/molbev/msx162. PMID 28505375. PMC 5850466.
- ↑ (2016. május 1.) „First multigene analysis of Archamoebae (Amoebozoa: Conosa) robustly reveals its phylogeny and shows that Entamoebidae represents a deep lineage of the group”. Molecular Phylogenetics and Evolution 98, 41–51. o. DOI:10.1016/j.ympev.2016.01.011. PMID 26826602.
- ↑ (2019. március) „Towards a phylogenetic classification of the Myxomycetes”. Phytotaxa 399 (3), 209–238. o. DOI:10.11646/phytotaxa.399.3.5.
- ↑ (2020) „Outline of Fungi and fungus-like taxa”. Mycosphere 11, 1060–1456. o. DOI:10.5943/mycosphere/11/1/8.
- ↑ (2003) „Vase-shaped microfossils from the Neoproterozoic Chuar Group, Grand Canyon: a classification guided by modern testate amoebae”. Journal of Paleontology 77 (3), 409–429. o. DOI:<0409:VMFTNC>2.0.CO;2 10.1666/0022-3360(2003)077<0409:VMFTNC>2.0.CO;2.
- ↑ Porter, Susannah M.. The Proterozoic Fossil Record of Heterotrophic Eukaryotes, Neoproterozoic Geolobiology and Paleobiology, Topics in Geobiology. Springer, 1–21. o.. DOI: 10.1007/1-4020-5202-2 (2006). ISBN 978-1-4020-5201-9
- ↑ (2015. június 1.) „Is there evidence of sexual reproduction (meiosis) in Acanthamoeba?”. Pathogens and Global Health 109 (4), 193–5. o. DOI:10.1179/2047773215Y.0000000009. PMID 25800982. PMC 4530557.
- ↑ a b c d (2015) „Entamoeba histolytica Dmc1 Catalyzes Homologous DNA Pairing and Strand Exchange That Is Stimulated by Calcium and Hop2-Mnd1”. PLOS ONE 10 (9), e0139399. o. DOI:10.1371/journal.pone.0139399. PMID 26422142. PMC 4589404.
- ↑ a b (2013) „Homologous recombination occurs in Entamoeba and is enhanced during growth stress and stage conversion”. PLOS ONE 8 (9), e74465. o. DOI:10.1371/journal.pone.0074465. PMID 24098652. PMC 3787063.
- ↑ (2018. november 1.) „Comparative genomics supports sex and meiosis in diverse Amoebozoa”. Genome Biol Evol 10 (11), 3118–3128. o. DOI:10.1093/gbe/evy241. PMID 30380054. PMC 6263441.
- ↑ Entamoebiasis - MeSH - NCBI. www.ncbi.nlm.nih.gov . (Hozzáférés: 2015. július 21.)
- ↑ Entamoebiasis. mesh.kib.ki.se . [2015. július 22-i dátummal az eredetiből archiválva]. (Hozzáférés: 2015. július 21.)
- ↑ a b c d e f g h i j k l m n o p q r Manson's Tropical Diseases. Elsevier Health Sciences, 664–671. o. (2013. október 26.). ISBN 978-0-7020-5306-1
- ↑ 19, Lecture Notes: Tropical Medicine. John Wiley & Sons, 177–182. o. (2014. április 17.). ISBN 978-1-118-73456-8
Fordítás
szerkesztésEz a szócikk részben vagy egészben az Amoebozoa című angol Wikipédia-szócikk ezen változatának fordításán alapul. Az eredeti cikk szerkesztőit annak laptörténete sorolja fel. Ez a jelzés csupán a megfogalmazás eredetét és a szerzői jogokat jelzi, nem szolgál a cikkben szereplő információk forrásmegjelöléseként.
Források
szerkesztés- The Amoebozoa, Dictyostelium discoideum Protocols, Methods in Molecular Biology, 1–15. o.. DOI: 10.1007/978-1-62703-302-2_1 (2013). ISBN 978-1-62703-301-5
További információk
szerkesztés- Pawlowski, Jan: Molecular Phylogeny of Amoeboid Protists — Tree of Amoebozoa. Molecular Systematics Group (MSG). Department of Zoology and Animal Biology, University of Geneva. [2005. január 2-i dátummal az eredetiből archiválva].
- Tree of Life Eukaryotes. Tree of Life Project
- Leidy, Joseph. Amoeba Plates, Fresh-Water Rhizopods of North America. Washington D.C.: Government Printing Office (1879)
- Amoebozoa. NCBI Taxonomy Browser